

Ботанический журнал, 2019, T. 104, № 6, стр. 888-899
Влияние осушения на динамику растительного покрова мезотрофных травяно-сфагновых болот южной Карелии
С. И. Грабовик 1, Л. В. Канцерова 1, *, С. А. Кутенков 1
1 Институт биологии Карельского научного центра РАН
185910 г. Петрозаводск,
ул. Пушкинская, 11, Россия
* E-mail: Kancerova.L@mail.ru
Поступила в редакцию 07.05.2019
После доработки 14.05.2019
Принята к публикации 16.05.2019
Аннотация
Исследования проводились на территории Киндасовского лесо-болотного научного стационара Карельского научного центра РАН, в подзоне средней тайги в заказнике Койву-Ламбасуо. В статье представлены результаты 4-летних исследований пост-мелиоративной динамики видового состава и структуры растительного покрова в пространственно-временном аспекте, который позволил проследить ход сукцессий на примере мезотрофного травяно-сфагнового болотного массива (аапа болото). Под влиянием осушения и сукцессий растительного покрова на аапа болотах отмечается изменение видового состава растительного покрова, снижение его биоразнообразия и выравнивание микрорельефа.
Растительный покров осушенных болот имеет сложную структуру, которая зависит от ряда факторов: от типа болотного массива, видового состава растительных сообществ, интенсивности и давности осушения. Здесь важную роль играет экологическая пластичность видов растений, слагающих фитоценозы, а также конкурентная способность видов, внедряющихся после осушения. Наиболее чутко реагируют на осушение гипергидрофильные виды, такие как Carex chordorrhiza Ehrh., C. limosa L., Sphagnum subsecundum Nees, которые требовательны к постоянному наличию грунтовых вод, полностью исчезнувших с болотного участка в первые годы после осушения. Сукцессионный процесс идет в сторону мезофитизации фитоценозов по сравнению с исходными. Анализ наших данных свидетельствует о том, что спустя 47 лет после осушения болота растительный покров не достигает устойчивого состояния, а находится на стадиях сукцессий, что требует дальнейших наблюдений за его динамикой.
Одним из наиболее распространенных способов трансформации болотных экосистем таежной зоны является лесоосушительная мелиорация, в результате которой происходит изменение гидрологического режима болотных массивов. Все это приводит к изменениям состава и структуры растительного покрова на осушенных болотах, которые происходят с различной скоростью в зависимости от типов болот, интенсивности осушения, а также их географического положения.
Изменению растительности болот и заболоченных лесов под влиянием осушения посвящено достаточно много работ как у нас в стране, так и за рубежом (Sarasto, 1957; Eliseeva, 1964; Bush, Abolin’, 1968; Laine et al., 1995; Neshataev, 1986). Анализ литературы показывает, что большая часть материалов об изменении растительного покрова болот и заболоченных лесов получена на основе однократных учетов, проведенных через достаточно большой период после осушения. При этом динамика напочвенного покрова рассматривается в сравнительно немногих работах, в основном на болотах Европейской части России (Nitsenko, 1951; Yurkovskaya, 1963; Guzlena, 1963); в Западной Сибири (Eliseeva, 1964; Platonov, 1967), основное же внимание уделяется реакции на осушение древесного яруса или его формированию на открытых болотах.
Гораздо меньше данных, полученных на постоянных пробных площадях путем повторных исследований в первые 5–10–15 лет после осушения, которые позволяют полнее раскрыть механизмы влияния осушения не только на древесный ярус, но и на напочвенный покров и экологические условия произрастания (Grabovik, 1989; Kuznetsov, Grabovik, 2010).
Болота на территории Карелии занимают 3.6 млн га, из них на долю травяно-сфагново-гипновых (карельские кольцевые аапа болота) приходится 0.9 млн га. Значительная их часть в южной и средней Карелии была осушена в 60–70 годы прошлого века для целей лесного и сельского хозяйства (Medvedeva, 1989; Orlov, 1991; Sakovets et al., 2000).
В связи с этим целью данной работы явилось представление результатов 47-летних исследований постмелиоративной динамики видового состава и структуры растительного покрова в пространственно-временном аспекте на примере травяно-сфагнового болотного массива Койвусуо.
МАТЕРИАЛЫ И МЕТОДЫ
Комплексные научно-исследовательские работы по изучению структуры и динамики болот и заболоченных лесов в естественном состоянии и под влиянием мелиорации проводятся с 1970 года на территории Киндасовского лесо-болотного научного стационара Карельского научного центра РАН, в подзоне средней тайги в заказнике Койву-Ламбасуо (61°48' с.ш., 33°35' в.д.). В климатическом отношении территория стационара характеризуется следующими средними многолетними данными: продолжительность вегетационного периода – 148 дней, температура воздуха за вегетационный период – 11.7°С, количество атмосферных осадков за год 565 мм, за вегетационный период – 316 мм.
На территории стационара наиболее распространены четыре типа болотных массивов: сфагновый грядово-мочажинный олиготрофный, травяно-сфагновый мезотрофный, травяно-сфагново-гипновый (карельские кольцевые аапа болота) и мезотрофный сфагновый лесной (Elina et al., 1984), значительная их часть до 1969 г. была в естественном состоянии, затем в период 1969–1974 гг. часть болот была осушена.
Исследования динамики видового состава и структуры растительного покрова выполнялись апробированными и модифицированными методами (Grabovik, 1989). Таксономия сосудистых растений приводится по С.К. Черепанову (Cherepanov, 1995) с учетом более поздней работы А.В. Кравченко (Kravchenko, 2007), мхов по М.С. Игнатову и др. (Ignatov et al., 2006).
Основные таксационные показатели древостоя определялись методами, применяемыми в лесной таксации (Anuchin, 1982).
В пределах изучаемых болотных комплексов выбирали наиболее типичные для них участки (по форме микрорельефа и растительному покрову) – постоянные пробные площади (ППП), на которых в последующие годы вели постоянные наблюдения. Размеры ППП (от 50 до 150 м2) зависели от сложности структуры болотного комплекса и величины отдельных элементов микрорельефа. Всего было заложено 11 пробных площадей, на которых выполняли геоботанические описания, отбирались образцы торфа на ботанический состав и степень разложения. Горизонтальная структура растительного покрова исследовалась методом крупномасштабного картирования. Наблюдения за динамикой видового состава растительного покрова вели на постоянных метровых площадках (ПМП), расположенных в пределах изучаемого болотного комплекса. Для оценки положения сообществ в осях экологических факторов использованы двухфакторные экологические ряды В.Д. Лопатина (Lopatin, 1983) для болотной растительности. В качестве меры сходства сообществ применен коэффициент Съеренсена–Чекановского (Ks), учитывающий проективное покрытие видов. Данный коэффициент использован как для сравнения площадок между собой, так и во временном ряду для каждой отдельной метровой площадки.
Болотный массив Койвусуо, площадью 40 га, относится к южнокарельскому варианту аапа болот и имеет торфяную залежь мощностью около 1.5 м. Пробная площадь, заложена в центральной части болотного массива, в кочковато-топяном комплексе Sphagneta magellanici + Herbeta, который занимал 27% от площади массива.
Болото осушено в 1971 г., расстояние между осушительными каналами 100 м, в настоящее время каналы заросли травянистой растительностью.
До осушения при геоботаническом описании во флористическом составе участка отмечено 20 видов: деревьев – 2, кустарничков – 4, трав – 8, мхов – 6. Отдельные сосны и березы, высотой 1–2 м, приурочены к повышениям. Для более детального изучения горизонтальной структуры растительного покрова в центральной части участка было сделано крупномасштабное картирование. Кочки занимали 36%, их растительный покров был представлен фитоценозами Andromeda polifolia–Carex lasiocarpa–Sphagnum magellanicum + S. angustifolium. Сообщества мочажин Carex lasiocarpa–Sphagnum subsecundum и Carex limosa–Menyanthes trifoliata занимали 64% (табл. 1). Проективное покрытие травяно-кустарничкового яруса повышений 80%, мхов – 100%, понижений, соответственно, 65 и 55%.
Мониторинг изменений растительного покрова вели на постоянных метровых площадках размером 1 × 1 м, в центральной части массива, в пределах участка кочковато-топяного комплекса Sphagneta magellanici + Herbeta. Всего, на различном удалении от мелиоративных каналов и в условиях разных элементов микрорельефа заложено 13 метровых площадок. В первые годы после осушения на ПМП проективное покрытие видов (%) травяно-кустарничкового и мохового ярусов оценивалось с периодичностью 1–2 года, реже до 7 лет. Первые наблюдения сделаны в год проведения гидромелиорации болотного массива.
РЕЗУЛЬТАТЫ
Растительный покров осушенных болот имеет сложную структуру, которая зависит от ряда факторов: от типа болотного массива, видового состава растительных сообществ, интенсивности и давности осушения. На разных типах болот изменение структуры растительных сообществ происходит неодинаково. Здесь важную роль играет экологическая пластичность видов растений, слагающих фитоценозы, а также конкурентная способность видов, внедряющихся после осушения.
Динамика видового состава растительного покрова центральной части аапа болота
Активные изменения растительного покрова в пределах отдельных метровых площадок наблюдались с момента проведения мелиорации в течение 13 лет (до 1984 г.), когда значение Ks между 6-летними срезами растительности составляло по площадкам в среднем 0.3–0.6, затем произошла относительная стабилизация растительности с Ks 0.6–0.9 за 6 лет. При этом наблюдалось и общее выравнивание состава растительности по участку, среднее значение Ks между метровыми площадками в первые 15 лет составляло 0.4, в последние годы – 0.5–0.7.
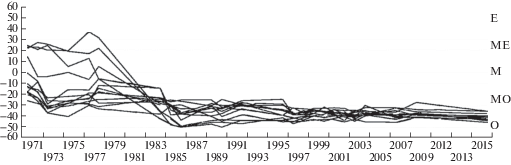
По отношению к фактору увлажнения растительность от соответствующей коврам и влажным мочажинам, преобладающей на начальном этапе, в течение 13 лет сменилась на характерную для низких и средних кочек (рис. 1). В это же время произошли и значительные изменения по отношению к фактору трофности. Изначально широкий спектр сообществ – от мезоевтрофных до мезоолиготрофных, сузился до олиготрофного типа (рис. 2). Наибольшие изменения видового состава наблюдались на изначально более сырых и богатых питанием метровых площадках. В дальнейшем положение сообществ в осях экологических факторов практически не изменялось.
Рис. 1.
Изменение положения растительности постоянных метровых площадок в экологическом ряду условий увлажнения (Лопатин, 1983).
Примечания. Микрорельеф и ступени увлажнения: HH – высокие кочки, MH – средние кочки, LH – низкие кочки, L – ковры, SF – маловлажные мочажины, F – влажные мочажины.
Fig. 1. Change of the position of the permanent sample plot’s vegetation in the moisture ecological series (Lopatin, 1983).
Notes. Microtopography and moisture levels: HH – high hummocks, MH – middle hummocks, LH – low hummocks, L – lawns, SF – slightly wet flarks, F – wet flarks.
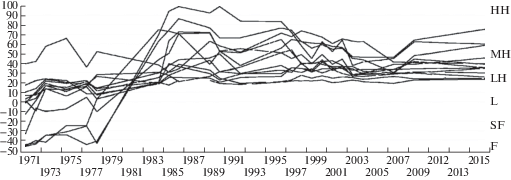
Рис. 2.
Изменение положения растительности постоянных метровых площадок в экологическом ряду типов питания (Лопатин, 1983).
Примечания. Тип питания: E – евтрофный, ME – мезоевтрофный, M – мезотрофный, МО – мезоолиготрофный, O – олиготрофный.
Fig. 2. Change of the position of the permanent sample plot’s vegetation in the nutrition types ecological series (Lopatin, 1983).
Notes. Nutrition type: E – eutrophic, ME – mesoeutrophic, M – mesotrophic, МО – mesooligotrophic, O – oligotrophic.
В отношении реакции отдельных видов на осушение (рис. 3) можно выделить шесть групп растений: 1) быстро (в течение 5 лет) исчезнувшие на участке: Menyanthes trifoliata L., Carex chordorrhiza, C. limosa, Sphagnum subsecundum – виды, характерные для богатых низинных мочажин; 2) с постепенно снизившимся покрытием: Eriophorum angustifolium Honck., Andromeda polifolia L., Carex lasiocarpa Ehrh.; 3) остающиеся относительно стабильными: Chamaedaphne calyculata (L.) Moench; 4) с возросшим, а затем снизившимся покрытием: Betula nana L., Polytrichum strictum Brid., Pleurozium schreberi (Brid.) Mitt., Sphagnum capillifolium (Ehrh.) Hedw.; 5) с противоположной предыдущим видам динамикой, с провалом в средней части периода наблюдения: Sphagnum fuscum (Schimp.) H. Klinggr. и S. magellanicum Brid11; 6) с увеличившимся покрытием: Oxycoccus palustris Pers., Vaccinium vitis-idaea L., V. uliginosum L., Eriophorum vaginatum L., S. angustifolium (C.E.O. Jensen ex Russow) C.E.O. Jensen. Покрытие последнего вида поначалу снизилось, но затем значительно возросло, достигнув максимума среди всех видов за весь период наблюдения.
Рис. 3.
Изменение проективных покрытий (%) основных видов растений на мелиорированном участке (усредненные значения по всем постоянным метровым площадкам, сглаженные за три года).
Fig. 3. Change of the percent cover (%) of major plant species in the drained site (3-year smoothed values averaged across all permanent sample plots).
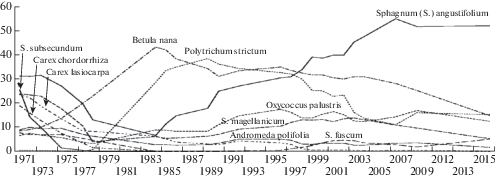
Таким образом, среди видов-эдификаторов растительного покрова на изменения гидрологического режима в первую очередь отреагировали виды, требовательные к постоянному наличию грунтовых вод, полностью исчезнув с болотного участка. Положительным образом быстрее других на понижение уровня грунтовых вод и улучшение аэрации корнеобитаемого слоя среагировали Betula nana и Polytrichum strictum, достигнувшие в течение 10–14 лет 40% покрытия и, затем, около 20 лет доминирующие на участке. У данных видов не только возросло покрытие на кочках, ими также были заселены бывшие топкие мочажины с отмершим Sphagnum subsecundum. Их развитие, наряду с понижением уровня воды, на некоторое время подавило рост сфагновых мхов. Формирование древесного яруса, в свою очередь, привело к снижению покрытия Betula nana. Через 30 лет после осушения выраженным эдификатором напочвенного покрова стал Sphagnum angustifolium, распространившийся по всем ПМП. Betula nana так же отмечается во всех ПМП, при этом ее жизненность и общее покрытие постепенно снижаются.
Сукцессии растительного покрова в пространственно-временном аспекте
В это же время на болотном участке происходило переформирование кочковато-топяного болотного комплекса Sphagneta magellanici + Herbeta и развитие древостоя.
Так, через 5 лет после осушения на месте прежнего кочковато-топяного комплекса сформировался кочковато-равнинный Mixtofruticuleto–Sphagneta angustifolii+Herbeta. Растительный покров кочек образовали фитоценозы Betulanana+Andromeda polifolia–Sphagnum magellanicum + S. angustifolium, а ковров – фитоценоз Carex lasiocarpa–Menyanthes trifoliata (табл.). Из растительного покрова исчезли гипергигрофильные виды Carex limosa, C. rostrata Stoces, C. chordorrhiza, S. subsecundum. Доминантные виды остались те же, но изменились их жизненность и проективное покрытие.
Через 13 лет после осушения на месте кочковато-равнинного Mixtofruticuleto–Sphagneta angustifolii+Herbeta сформировался кочковато-равнинный комплекс Betuleta nanae+Herbeta. Растительный покров кочек представлен фитоценозами Betula nana–Sphagnum angustifolium + S. magellanicum, а ковров – Carex lasiocarpa–S. angustifolium (табл.). На кочках Betula nana усиленно разрастается в связи с улучшением аэрации корнеобитаемого слоя. Ухудшение водного режима и освещения на кочках снижают жизненность и проективное покрытие сфагновых мхов, начинается их деградация. Зеленые мхи, а именно Dicranum polysetum Sw. и Polytrichum strictum полностью заселяют участки с мертвым Sphagnum subsecundum, в это время поселяется и Polytrichastrum longisetum (Sw. ex Brid.) G.L. Sm. В это же время здесь началось поселение сосны. Общее число стволов составляет до 1103 экз./га, в том числе сосны 937, березы – 166 экз./га (Grabovik, 2007).
Через 35 и 47 лет после осушения сформировался сосняк ерниково-сфагновый с мозаичной структурой, обусловленной размерами клонов Betula nana, с редким (относительная полнота 0.36) древесным ярусом из сосны (средняя высота до 6 метров, средний диаметр 8 см) (табл.) Состав древостоя 10С35 +Б. Общее количество стволов 1663 экз./га, в том числе сосны 1292 экз./га, березы 371 экз./га. В последние годы появились всходы ели до 540 экз./га и сосны – 70 экз./га высотой до 0.7 м. В напочвенном покрове отмечается снижение жизненности и проективного покрытия Betula nana (15%) и Andromeda polifolia (10%).
Происходит дальнейшее сглаживание микрорельефа и расселение Sphagnum angustifolium по бывшим топким мочажинам, отмечается не только изменение видового состава участка, встречаемости, проективного покрытия ряда видов. Наблюдается ослабление фитоценотической роли Carex lasiocarpa, уменьшение ее жизненности, высота ее падает до 30–40 см, она не цветет и не плодоносит, это свидетельствует о застойном режиме увлажнения и значительном снижении уровня грунтовых вод (УГВ) до –25–30 см ниже поверхности, что также подтверждается и исчезновением Menyanthes trifoliata. Изменения состава флоры участка направлены в сторону полного исчезновения или снижения жизненности гигрофильных трав и мхов, снизились жизненность и обилие большинства болотных видов кустарничков и трав и только Andromeda polifolia и особенно Betula nana обильно разрослись после осушения. В микроценозах с разреженным ерником, занятых дернинками Polytrichum strictum и Polytrichastrum longisetum, от которых микрорельеф становится мелкокочковатым, поселяются небольшие клоны Vaccinium uliginosum и V. vitis-idaea. В это же время отмечается появление елового подроста.
ОБСУЖДЕНИЕ
Наши данные согласуются с исследованиями Т.К. Юрковской (Yurkovskaya, 1963), которая в Карелии изучала изменение растительного покрова под влиянием осушения безлесных травяно-сфагновых переходных болот и сосняков сфагновых с давностью осушения 30–50 лет. При интенсивном осушении травяно-сфагновых переходных болот доминантами травяно-кустарничкового и мохового ярусов в сформировавшихся производных лесных сообществах стали лесные растения и лесные мхи. При более слабой степени осушения изменения, происшедшие в напочвенном покрове, незначительны, исчезают лишь самые гидрофильные болотные растения, увеличивается обилие болотных кустарничков, лесные растения встречаются единично на приствольных повышениях. Изменения напочвенного покрова зависят не только от степени осушения, но и от давности осушения. Так, исследования Т.К. Юрковской показали, что на переходных травяно-сфагновых болотах через 3 года после осушения видовой состав не только не изменился, но сохранились почти те же количественные соотношения. Изменяется лишь ценотическая роль отдельных видов осок.
Финские исследователи (Laine et al., 1995) изучали изменения напочвенного покрова в сосняках сфагновых с давностью осушения от 3 до 55 лет. Они отмечают, что в первые годы после осушения исчезают гидрофильные осоки (Carex lasiocarpa и C. rostrata), покрытие болотных кустарничков снижается с возрастанием сомкнутости древесного яруса, участие сфагновых мхов с уменьшением освещенности происходит в следующем порядке Sphagnum fuscum > S. magellanicum > S. russowii Warnst.
Исследования, проведенные В.Н. Федорчуком (Fedorchuk et al., 2005), на осушенных болотах северо-запада Европейской части России, показали, что стадии относительной стабилизации растительного покрова обычно наступают не ранее 20–25 лет после мелиорации.
Финские исследователи L. Kheikurainen (1983) и J. Laine (1989) указывают, что в Ленинградской области в результате осушения болот и заболоченных лесов в течение 20–90 лет формируются леса различного типа.
ЗАКЛЮЧЕНИЕ
Детальный анализ динамики видового состава и структуры растительного покрова мезотрофного травяно-сфагнового болота в пространственно-временном аспекте позволил проследить ход сукцессий, выражающийся в обобщенных временных сериях. Эти серии являются моделью, отображающей направление происходящих смен фитоценозов во времени под влиянием осушения.
Под влиянием осушения и сукцессий растительного покрова происходит выравнивание микрорельефа.
На аапа болотах отмечается не только изменение видового состава растительного покрова, но и снижение его биоразнообразия. Наиболее чутко реагируют на осушение гипергидрофильные виды, такие как Carex chordorrhiza, C. limosa, Sphagnum subsecundum, которые требовательны к постоянному наличию грунтовых вод, полностью исчезнув с болотного участка в первые годы после осушения. Растительный покров осушенных болот имеет сложную структуру, которая зависит от ряда факторов: от типа болотного массива, видового состава растительных сообществ, интенсивности и давности осушения.
Важную роль играет экологическая пластичность видов растений, слагающих фитоценозы, а также конкурентная способность видов, внедряющихся после осушения. Сукцессионный процесс идет в сторону мезофитизации фитоценозов по сравнению с исходными.
Анализ наших данных по постмелиоративной динамике видового состава и структуры растительности свидетельствует о том, что спустя 47 лет после осушения растительный покров не достиг устойчивого состояния, а находится на стадиях сукцессий, что делает оправданным дальнейшее наблюдение за его динамикой
Список литературы
[Anuchin] Анучин Н.П. 1982. Лесная таксация. М. 552 с.
[Bush, Abolin’] Буш К.К., Аболинь А.А. 1968. Строение и изменение растительного покрова важнейших типов леса под влиянием осушения. – В кн.: Вопросы гидролесомелиорации. Рига. С. 71–126.
[Cherepanov] Черепанов С.К. 1995. Сосудистые растения России и сопредельных государств (в пределах бывшего СССР). СПб. 992 с.
[Elina et al.] Елина Г.А., Кузнецов О.Л., Максимов А.И. 1984. Структурно-функциональная организация и динамика болотных экосистем Карелии. Л. 128 с.
[Eliseeva] Елисеева В.М. 1964. К вопросу об изменении естественной растительности под влиянием осушения. – Изв. Томск. Отд. Всесоюзн. Ботан. об-ва. 5: 85–87.
[Grabovik] Грабовик С.И. 1989. Динамика растительного покрова болотных массивов мезотрофного травяно-сфагнового типа под влиянием осушения. – Бот. журн. 74 (12): 1757–1768.
[Grabovik] Грабовик С.И. 2007. Постмелиоративная динамика биологической продуктивности мезотрофных травяно-сфагновых болот южной Карелии. – Бот. журн. 92 (5): 670–680.
[Guzlena] Гузлена А.Д. 1963. Изменения растительного покрова низинного болота Бейбежи под влиянием осушения и освоения. – Уч. зап. Тарт. гос. ун-та. 145(7): 298–305.
Hassel K., Kyrkjeeide M.O., Yousefi N., Prestø T., Stenøien H.K., Show J.A., Elatberg K.I. 2018. Sphagnum divinum (sp. nov.) and S. medium Limpr. and their relationship to S. magellanicum Brid. – Journal of Bryology. 40(3): 197–222. https://doi.org/10.1080/03736687.2018.1474424
Ignatov M.S., Afonina O.M., Ignatova E.A. et al. 2006. Check-list of mosses of East Europe and North Asia. – Arctoa. 15: 1–130.
[Kravchenko] Кравченко А.В. 2007. Конспект флоры Карелии. Петрозаводск. 403 с.
[Kuznetsov, Grabovik] Кузнецов О.Л., Грабовик С.И. 2010. Мониторинг флоры и растительности болотных экосистем. – В кн: Мониторинг и сохранение биоразнообразия таежных экосистем Европейского Севера России. Петрозаводск. С. 19–31.
Laine J. 1989. Metsaojitettujen soiden luokittelu (English summary: Classification of peatlands drained for forestry). – Suo. 40: 37–51.
Laine J., Vasander H., Laiho R. 1995. Long-term effects of water level drawdown on the vegetation of drained pine mires in southern Finland. – J. Appl. Ecol. 32: 785–802.
[Lopatin] Лопатин В.Д. 1983. Экологические ряды растительности болот. – В кн: Структура растительности и ресурсы болот Карелии. Петрозаводск. С. 5–38.
[Medvedeva] Медведева В.М. 1989. Формирование лесов на осушенных землях среднетаежной подзоны. Петрозаводск. 168 с.
[Neshataev] Нешатаев В.Ю. 1986. Изменение растительности травяно-сфагновых сосняков под влиянием осушения. – Бот. журн. 71 (4): 429–440.
[Nitsenko] Ниценко А.А. 1951. Наблюдения над изменениями растительного покрова под влиянием осушения. – Бот. журн. 36 (34): 349–355.
[Orlov] Орлов Е.Д. 1991. Грунтовое водное питание на объектах лесоосушения в Карелии. Л. 164 с.
[Platonov] Платонов Г.М. 1967. Смена растительности болот под влиянием осушения. – Взаимоотношения леса и болота. М. С. 128–140.
[Sakovets et al.] Саковец В.И., Германова Н.И., Матюшкин В.А. 2000. Экологические аспекты гидролесомелиорации в Карелии. Петрозаводск. 155 с.
Sarasto Ju. 1957. Metsan kasvattamiseksi ojiteeujen soiden aluskasvillisunden rakenteesta ja kehityksesta Suomen etelapuoliskossa. – Acta Forestation Fennica. 65 (7): 1–108.
[Fedorchuk et al.] Федорчук В.Н., Нешатаев В.Ю., Кузнецова М.Л. 2005. Лесные экосистемы северо-западных районов России – В кн.: Типология, динамика, хозяйственные особенности. Санкт-Петербург. 382 с.
[Kheikurainen] Хейкурайнен Л. 1983. Болота. Москва. 40 с.
[Yurkovskaya] Юрковская Т.К. 1963. Изменение растительного покрова переходных болот южной Карелии под влиянием осушения. – Учен. зап. Тарт. ун-та. 145 (7): 337–345.
Дополнительные материалы отсутствуют.
Инструменты
Ботанический журнал