

Ботанический журнал, 2019, T. 104, № 6, стр. 900-917
МОРФОЛОГИЯ ПЫЛЬЦЕВЫХ ЗЕРЕН И ИНДИВИДУАЛЬНАЯ ИЗМЕНЧИВОСТЬ ФОРМЫ РАСПОЛОЖЕНИЯ АПЕРТУР ПЫЛЬЦЫ ПРЕДСТАВИТЕЛЕЙ РОДА NICOTIANA (SOLANACEAE)
В. В. Григорьева 1, *, А. Е. Пожидаев 1, **, А. Н. Семенов 1, Д. А. Брицкий 1, ***
1 Ботанический институт им. В.Л. Комарова РАН
197376 Санкт-Петербург,
ул. Проф. Попова, 2, Россия
* E-mail: mikhailov_val@mail.ru
** E-mail: pae62@mail.ru
*** E-mail: dmibri@mail.ru
Поступила в редакцию 17.01.2019
После доработки 15.03.2019
Принята к публикации 16.05.2019
Аннотация
Проведены исследования морфологии пыльцевых зерен 16 видов рода Nicotiana методами световой и сканирующей электронной микроскопии. Типичные пыльцевые зерна изученных видов 3-бороздно-оровые, эллипсоидальные, широко эллипсоидальные или почти сфероидальные, средних размеров, длина полярной оси не превышают 50 мкм. У 8 изученных видов встречаются единичные пыльцевые зерна, отличающиеся от типичных (3-бороздно-оровых) числом и расположением апертур. Все обнаруженные формы расположения апертур (типичные и атипичные) могут быть выстроены в непрерывный и геометрически закономерный морфологический ряд. Закономерности многообразия форм пыльцы оказываются в высокой степени транзитивны (таксон-неспецифичны и рангово независимы). Хотя палиноморфологические признаки не удается использовать для уточнения границ подродов и секций Nicotiana, форма пыльцевых зерен, особенности строения апертур, детали скульптуры являются хорошими дополнительными признаками для характеристики некоторых видов или отдельных групп видов Nicotiana.
Род Nicotiana (подсемейство Nicotianoideae) – один из пяти крупных родов семейства Solanaceae. T.H. Goodspeed (1954) рассматривал род в объеме 64 видов, 44 из которых произрастают в Северной и Южной Америке, 20 видов являются нативными видами Австралии. Два вида N. tabacum L. и N. rustica L. широко культивируются в тропических и субтропических областях. Центром происхождения и разнообразия Nicotiana считается Южная Америка (Goodspeed, 1947; Chase, 2003). В современных работах, посвященных молекулярной филогении рода, указывается разное число видов Nicotiana: 60–95 (Borisjuk et al., 1997) и 75–76 (Сlarkson et al., 2004). Жизненные формы представителей рода разнообразны. Среди видов Nicotiana встречаются низкорослые однолетники высотой несколько сантиметров, высокие более 1.5 м травянистые многолетники и кустарники. Разнообразны размеры, форма и окраска цветков.
По данным макроморфологии, кариологии и географии род разделен на 3 подрода: Rustica (секции: Paniculatae, Thyrsiflorae, Rusticae); Tabacum (секции: Tomentosae, Genuinae); и Petunioides (секции:Undulatae, Trigonophyllae, Alatae, Noctiflorae, Acuminatae, Suaveolentes) (Goodspeed, 1947, 1954).
Пыльца представителей рода Nicotiana на сегодняшний день исследована недостаточно подробно. Сведения о морфологии пыльцы отдельных видов табака можно найти в статьях, посвященных палиноморфологии семейства Solanaceae (Basak, 1967; Murry, Eshbaugh, 1971; Goodward et al., 1994; Perveen, Qaiser, 2007, Finot et al., 2018 и др.). Для многих видов морфологические данные и изображения пыльцы получены только с помощью свето-оптического микроскопа. Самая значительная по числу исследованных видов работа содержит данные по морфологии пыльцы 10 видов Nicotiana флоры Чили (Collao-Alvarado et al., 2016). J.W. Gish (2000) изучил пыльцу 7 видов Nicotiana с помощью светооптического и электронного сканирующего микроскопов. Ультраструктура спородермы 3 видов Nicotiana (N. sanderae Hort. ex Watson, N. alata Link & Otto, N. forgetiana Hort. ex Hemsley) была изучена с помощью трансмиссионного электронного микроскопа (Goodward, et al., 1994); было показано, что спородерма имеет тектатно-колумеллятное строение.
Пыльцевые зерна различных Solanaceae, в том числе и Nicotiana, нечасто, но отмечаются в составе спорово-пыльцевых спектров геологических отложений, начиная с Миоцена. При этом их определение даже до рода затруднено (Leopold, 1969; Warner, 2012; Särkinen et al., 2013). Пыльцевые зерна представителей рода Nicotiana сходны с пыльцевыми зернами некоторых видов Cestrum, Petunia, Lycium (Holloway, Dean, 2000). Кроме того, пыльца Nicotiana была обнаружена при археологических раскопках в Калифорнии и Колорадо (Gish, 2000). Так же палинологический метод широко используют при исследовании древних курительных трубок (Bryant et al., 2012).
В связи с этим встает проблема идентификации видов рода Nicotiana по пыльце. Для чего необходимо выявление таксономически значимых морфологических признаков, позволяющих достоверно отличать пыльцу Nicotiana от пыльцы других родов семейства Solanaceae. Кроме чисто практических задач, оценка таксономического значения наблюдаемой индивидуальной изменчивости морфологических признаков пыльцы актуальна с теоретической точки зрения (Pozhidaev, 2015). Поэтому цель данной работы – описать вариабельность признаков пыльцы у представителей рода Nicotiana и оценить уровень таксономической специфичности исследованных признаков. Для этого так же была исследована индивидуальная изменчивость признаков формы пыльцевого зерна (числа и расположения апертур).
Анализ литературы показал, что нет единого мнения о значении морфологических признаков пыльцы для систематики рода. T. Goodspeed (1954) в своей монографии, посвященной роду Nicotiana, основываясь на данных светооптической микроскопии, отмечал, что разные виды табака имеют сходную пыльцу. R.G. Holloway, G. Dean (2000), проведя статистическую обработку данных, также полученных с помощью светооптического микроскопа, пришли к выводу, что дискриминантный анализ позволяет отличать по палинологическим данным род Nicotiana от родов, имеющих сходную с ним пыльцу. Кроме того авторы отмечают, что возможно различать даже виды в пределах рода Nicotiana. Сравнительный морфологический анализ пыльцы чилийских видов (Collao-Alvarado et al., 2016) выявляет, по мнению авторов, незначительные различия пыльцы разных видов Nicotiana. Несмотря на то, что количественные признаки пыльцы (соотношение длины полярной оси и экваториального диаметра, длина борозд, толщина экзины) имеют статистически значимые различия, авторы делают вывод, что на видовом уровне значение признаков пыльцы ограничено.
Слабая степень изученности пыльцы Nicotiana не позволяет достоверно оценить таксономическое значение палиноморфологических признаков, ограничивает использование особенностей строения пыльцевых зерен для уточнения спорных вопросов систематики и филогении этого рода, а так же затрудняет определение дисперсной пыльцы этих растений в палиноспектрах различного происхождения.
МАТЕРИАЛ И МЕТОДИКА
Материалом для исследования послужили зрелые пыльцевые зерна 16 видов рода Nicotiana, взятые с гербарных образцов, хранящихся в Гербарии Ботанического института им. В.Л. Комарова (LE).
В работе использовали методы световой (СМ) и сканирующей (СЭМ) электронной микроскопии. Для светооптического исследования применяли классический ацетолизный метод Эрдтмана (Erdtman, 1952). Световые микрофотографии делались при увеличении 1000 раз на микроскопах Carl Zeiss Jenaval с помощью цифровой фотокамеры Canon EOS 20D.
Особенности строения поверхности пыльцевых зерен исследовали на сканирующем электронном микроскопе JEOL JSM-6390 в центре коллективного пользования Ботанического института им. В.Л. Комарова. Для исследования предварительно обработанную ацетолизной смесью пыльцу закрепляли на специальном столике с помощью двусторонней липкой ленты, затем напыляли сплав золота и палладия в вакуумной установке.
Описания проводили по общепринятой схеме. Учитывали: тип и число апертур, форму и очертания пыльцевых зерен, размеры полярной оси и экваториального диаметра, особенности строения борозд и межапертурных участков, толщину экзины и характер скульптуры (Kupriyanova, Aleshina, 1967, 1972). Измерения проводили с помощью окулярмикрометра. В каждом образце измеряли не менее 15 пыльцевых зерен.
Латинские названия видов рода Nicotiana и фамилии авторов (табл. 1) приведены в соответствии с системой T. Goodspeed (1954) и проверены по базе данных International Plant Names Index (IPNI). Исследованные образцы видов рода Nicotiana:
Таблица 1.
Описания пыльцевых зерен видов Nicotiana Table 1. Description of pollen grains of the studied Nicotiana species
| № | Исследованные
образцы Studied samples |
Длина полярной
оси (мкм) Polar axis length (μm) |
Длина
экваториального
диаметра (мкм) Equatorial axis length (μm) |
Ширина
борозд (мкм) Colpus width (μm) |
Ширина
мезокольпиума
(мкм) Mesocolpium width (μm) |
Диаметр
апокольпиума
(мкм) Apocolpium diameter (μm) |
Толщина
экзины (мкм) Exine thickness (μm) |
Cкульптура
(СЭМ) Sculpture (SEM) |
|---|---|---|---|---|---|---|---|---|
| 1 | N. acuminata (Graham) Hook. | 40.0–49.1 | 38.1–43.8 | 8.2–10.2 | 8.2–10.2 | 6.7–7.9 | 2.6–2.7 | Струйчатая. Струи длинные, до 8.5 мкм дл., 0.5–0.7 мкм шир., на расстоянии 0.1–0.3 мкм друг от друга/ Striate. The striae long, up to 8.5 μm long and 0.5–0.7 µm wide, 0.1–0.3 µm from each other |
| 2 | N. alata Link et Otto | 33.0–37.9 | 29.1–32.1 | 4.8–6.0 | 20.7–23.5 | 7.2–9.5 | 2.2–2.5 | Струйчатая. Струи недлинные, до 3.5 мкм дл., до 0.3 мкм шириной, слегка извитые, плотно
прилегают друг к другу/ Striate. The striae short, up to 3.5 μm long and 0.3 µm wide, tightly adjacent to each other |
| 3 | N. attenuata Torrey ex S. Watson |
35.6–42.9 | 32.1–39.5 | 8.2–9.1 | 25.3–27.1 | 6.0–7.8 | 2.1–2.4 | Струйчатая, на полюсах почти гладкая. Струи короткие, до 2.5 мкм дл., до 0.3 мкм шир., на небольшом расстоянии друг от друга/Striate; poles psilate. The striae short, up to 2.5 μm long and 0.3 µm wide, adjacent to each other |
| 4 | N. benavidesii Goodsp. | 34.3–42.3 | 27.0–30.0 | 5.0–5.8 | 24.7–26.8 | 5.2–8.8 | 2.0–2.7 | Струйчато-сетчатая; на полюсах и по краям борозд почти гладкая. Струи короткие, до
2.0 мкм дл., широкие, до 0.8 мкм шир., с редкими перфорациями. Striato-reticulate; poles psilate. The striae short and wide, up to 2 μm long and 0.8 µm wide, with rare perforations |
| 5 | N. berteroana G. Don | 37.5–47.0 | 34.9–40.0 | 6.3–7.5 | 19.4–21.4 | 4.5–5.0 | 2.0–2.7 | Струйчатая. Струи разноразмерные: короткие до 1.0 мкм, длинные до 5.0 мкм дл., на небольшом расстоянии друг от друга/Striate. The striae short (up to 1 μm long) and long (up to 5 μm long), adjacent to each other |
| 6 | N. bigelovii (Torrey) S. Watson | 37.2–44.2 | 30.0–40.0 | 6.8–8.9 | 25.7–28.7 | 5.8–12.0 | 2.1–2.5 | Струйчато-сетчатая. Струи 1.7–3.5 мкм длиной, около 0.2 мкм шириной/Striato-reticulate. The striae short, up to 1.7–3.5 μm long and 0.2 µm wide |
| 7 | N. bonariensis Lehm. | 32.8–36.4 | 25.7–28.7 | 4.2–4.5 | 18.9–20.8 | 3.0–5.0 | 1.5–2.0 | Струйчатая. Струи длинные, до 5 мкм дл., 0.3 мкм шир., плотно прилегают друг к другу/Striate. The striae long, up to 5 μm long and 0.3 µm wide, tightly adjacent to each other |
| 8 | N. californica Grabovetz. | 40.6–48.6 | 31.3–38.1 | 7.2–9.0 | 23.0–29.6 | 7.0–8.5 | 1.6–2.0 | Струйчато-сетчатая, на полюсах и по краям борозд перфорированная. Струи около 1.5
мкм дл., около 0.3 мкм шир./ Striato-reticulate; poles perforate. The striae short, up to 1.5 μm long and 0.3 µm wide |
| 9 | N. clevelandii A. Gray | 42.5–50.0 | 25.5–36.6 | 5.5–7.8 | 21.1–23.9 | 5.5–7.0 | 2.0–2.3 | Струйчатая. Струи 1.3–3.3 мкм дл., 0.5–0.8 мкм шир., на расстоянии около 0.2 мкм друг от друга/Striate. The striae short, 1.3–3.3 μm long and 0.5–0.8 µm wide. The distance between the striae is 2 µm |
| 10 | N. dilatata Link | 44.9–50.0 | 34.3–40.0 | 5.5–8.7 | 22.6–30.0 | 7.7–10.2 | 2.3–2.5 | Струйчато-гранулярная. Струи длинные, неширокие, направлены меридионально, практически параллельно друг другу. Между струями мелкие гранулы/Striato-granular. The striae long, narrow, directed meridionally, almost parallel to each other. Small granules are visible between the striae |
| 11 | N. glauca Graham | 28.8–35.5 | 28.6–31.4 | 5.9–6.6 | 20.0–23.1 | 6.6–10.0 | 1.6–2.0 | Струйчатая. Струи до 1.7 мкм длиной, до 0.3 мкм шириной/ Striate. The striae are short, up to 1.7 μm long and 0.3 µm wide |
| 12 | N. longiflora Cav. | 41.7–49.8 | 35.4–41.5 | 9.6–10.0 | 25.0–28.4 | 7.4–10.5 | 2.0–2.2 | Струйчатая. Струи длинные, 0.3-0.5 мкм шириной, располагаются на небольшом расстоянии друг от друга. Между струями перфорации/Striate. The striae are long, 0.3–0.5 µm wide. Small perforations are visible between the striae |
| 13 | N. noctiflora var. noctiflora Hook. | 35.9–40.0 | 33.3–37.3 | 6.9–8.7 | 21.8–27.5 | 5.2–9.1 | 1.8–2.1 | Струйчатая, на полюсе почти гладкая. Струи 1.6–2.5 мкм дл., до 0.6 мкм шир./Striate; poles psilate. The striae short, up to 1.6–2.5 μm long and 0.3 µm wide |
| 14 | N. paniculata L. | 30.0–36.4 | 30.0–35.4 | 5.8–6.8 | 24.0–25.3 | 7.0–8.0 | 1.8–1.9 | Струйчато-перфорированная. Струи недлинные, 0.3–0.5 мкм шир. Между струями перфорации/Striato-perforate. The striae short, 0.3–0.5 µm wide. Small perforations are visible between the striae |
| 15 | N. rustica L. | 33.1–36.4 | 25.2–34.0 | 2.1–3.3 | 19.6–21.5 | 5.1–8.0 | 1.5–2.2 | Струйчато-сетчатая. На полюсах почти гладкая/Striato-reticulate; poles psilate. |
| 16 | N. tabacum L. | 36.3–44.3 | 30.0–32.5 | 6.6–7.4 | 17.8–20.4 | 5.4–7.8 | 2.3–2.7 | Струйчато-сетчатая, на полюсах перфорированная/ Striato-reticulate; poles perforate |
N. acuminata (Graham) Hook. Herb. Mertens. Hort. Dr. Schmidt. 06.1844.
N. alata Link & Otto Ex hort. bot. Petropol. Herb. Fischer. N 34.
N. attenuate Torr. ex Watson Utah. San Juan Co. J.L. Gentry, G. Davidse. 31.07.1967. N 1766.
N. benavidesii Goodsp. Peru. Departm. Apurimac. Prov. Abancay. P. C. Hutchinson. 3.09.1957.
N. berteroana Hort. ex G. Don Cult. in hort. bot. Petropol. 1834 ex 1835.
N. bigelovii. (Torrey) S. Watson California. 1907. S.A. Barrett.
N. bonariensis Lehm. Herb. Americanum. Concepcion del Uruguay. P.G. Lorentz. 1877.
N. californica Hort. ex Grabovetz. Collectio Karpinskyana. 13.08.1852.
N. clevelandii A. Gray California. Bot. Gard. Of Univ. of California. 1923. A. Munz.
N. dilatata Link Herb. Fischer.
N. glauca Graham Jerusalem. 12.07.1954. I. Amdursky.
N. longiflora Cav. St. Hil. Ex horto bot. Petropol. Herder. 6.07.1858.
N. noctiflora var. noctiflora Hook. Argentina. 19.01.1944. C. A. O’Donell.
N. paniculata L. Horn. Ex Hort. Bot. Petropol. 21.07.1852.
N. rusica L. Flora Pekinensis, 1877, Dr. Bretschneider, N 544.
N. tabacum L. Mexique. Mont. Zacoaleo. 10.07.1865. M. Bourgeau. N 540; Ex horto. 1852. NW India. Hb. Royle.
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
Исследована морфология пыльцы 16 видов рода Nicotiana; описания пыльцы изученных видов представлены в таблице 1. В палиноморфологическом отношении род достаточно однороден, пыльца всех изученных видов может быть отнесена к одному палинотипу.
Типичные пыльцевые зерна изученных видов 3-бороздно-оровые, эллипсоидальные (табл. I, 2) или широко эллипсоидальные (табл. II, 7), реже почти сфероидальные; в очертании с полюса 3-лопастные, с экватора эллиптические или широкоэллиптические, с заостренными полюсами. Мезокольпиумы у пыльцы большинства исследованных видов ромбические (табл. I, 12) или близкие к ромбическим, реже эллиптические (табл. I, 2). Пыльца средних размеров, с полярной осью 30–50 мкм. Самые мелкие пыльцевые зерна обнаружены у N. glauca (полярная ось 28.8–35.5 мкм, экваториальный диаметр 28.6–31.4 мкм). Самая крупная пыльца у N. dilatata (полярная ось 44.9–50.0 мкм, экваториальный диаметр 34.3–40.0 мкм); у пыльцевых зерен N. clevelandii и N. acuminata длина полярной оси может достигать 50 мкм. Размеры зерен в пределах вида варьируют незначительно, в пределах 10 мкм. Максимальное варьирование длины полярной оси пыльцевых зерен отмечено у N. berteroana и составляет 9.5 мкм.
Таблица I. Морфология пыльцы представителей рода Nicotiana (СМ, СЭМ).
1–5 – N. tabacum; 6–10 – N. glauca; 11–15 – N. benavidesii. 1, 3, 6, 8, 11, 13 – общий вид пыльцевого зерна с полюса. 2, 4, 7, 9, 12, 14 – общий вид пыльцевого зерна с экватора. 5, 10, 15 – скульптура поверхности. 1, 2, 6, 7, 11, 12 – СМ; 3–5, 8–10, 13–15 – СЕМ. Для каждого пыльцевого зерна показан верхний и нижний фокус (CМ). Масштабные линейки, мкм: 1–4, 6–9, 11–14 – 10; 5, 10, 15 – 1.
Plate I . Pollen morphology of the Nicotiana species (LM, SEM).
1–5 – N. tabacum; 6–10 – N. glauca; 11–15 – N. benavidesii. 1, 3, 6, 8, 11, 13 – polar view of pollen grain. 2, 4, 7, 9, 12, 13 – equatorial view of pollen grain. 5, 10, 15 – exine ornamentation. 1, 2, 6, 7, 11, 12 – LM; 3–5, 8–10, 13–15 – SEM. The upper and lower foci are given for each pollen grain (LM). Scale bars, μm: –4, 6–9, 11–14 – 10; 5, 10, 15 – 1.
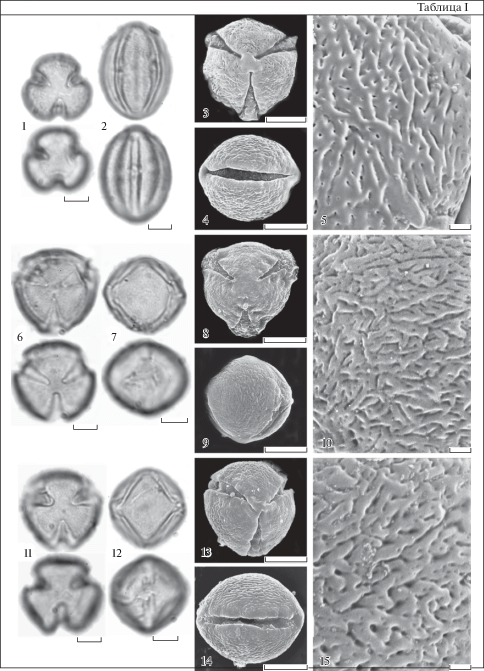
Таблица II. Морфология пыльцы представителей рода Nicotiana (СМ, СЭМ).
1–5 – N. bigelovii; 6–10 – N. berteroana; 11–15 – N. paniculata. 1, 3, 6, 8, 11, 13 – общий вид пыльцевого зерна с полюса. 2, 4, 7, 9, 12, 14 – общий вид пыльцевого зерна с экватора. 5, 10, 15 – скульптура поверхности. 1, 2, 6, 7, 11, 12 – СМ; 3–5, 8–10, 13–15 – СЕМ. Для каждого пыльцевого зерна показан верхний и нижний фокус (CМ). Масштабные линейки, мкм: 1–4, 6–9, 11–14 – 10; 5, 10, 15 – 1.
Plate II . Pollen morphology of the Nicotiana species (LM, SEM).
1–5 – N. bigelovii; 6–10 – N. berteroana; 11–15 – N. paniculata. 1, 3, 6, 8, 11, 13 – polar view of pollen grain. 2, 4, 7, 9, 12, 14 – equatorial view of pollen grain. 5, 10, 15 – exine ornamentation. 1, 2, 6, 7, 11, 12 – LM; 3–5, 8–10, 13–15 – SEM. The upper and lower foci are given for each pollen grain (LM). Scale bars, μm: 1–4, 6–9, 11–14 – 10; 5, 10, 15 – 1.
Апертуры сложные, состоят из внешней меридионально ориентированной борозды и внутренней экваториально расположенной оры. Борозды (экзоапертуры) длинные, глубокие, широкие, сужающиеся к полюсам, со слегка закругленными концами и более или менее ровными краями. Оры (эндоапертуры) у пыльцы большинства исследованных видов неясные, с нечеткими, часто неровными краями, иногда округлые или вытянутые вдоль полярной оси или в экваториальной плоскости. Оры небольшие, почти не заходят за границы борозд. Среди исследованных видов выделяется пыльца N. paniculata, с четко очерченными, крупными (до 10–11 мкм), почти округлыми орами (табл. II, 11, 15).
Скульптура струйчатая. При светооптическом исследовании у большинства видов почти неразличима; отчетливая струйчатость поверхности наблюдается только у пыльцы N. acuminata (табл. III, 11–15). Исследования показали, что особенности скульптуры, важные для характеристики пыльцы отдельных видов Nicotiana, можно выявить только при электронном микроскопировании, при увеличении не менее чем в 10 000 раз. Пыльцевые зерна разных видов различаются длиной, шириной струй, расстоянием между ними, характером расположения на поверхности пыльцевого зерна. Поверхность пыльцевых зерен на полюсах и по краям борозд у многих видов почти гладкая (табл. I, 4, 14). Экзина около 2 мкм толщиной. Слои экзины и скульптура поверхности пыльцевых зерен у большинства видов с помощью свето-оптического микроскопа почти неразличимы.
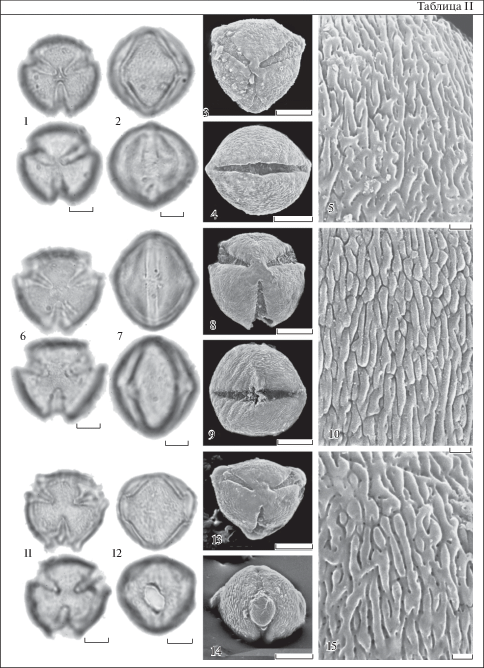
Таблица III. Морфология пыльцы представителей рода Nicotiana (СМ, СЭМ).
1–5 – N. noctiflora var. noctiflora; 6–10 – N. clevelandii; 11–15 – N. acuminata. 1, 3, 6, 8, 11, 13 – общий вид пыльцевого зерна с полюса. 2, 4, 7, 9, 12, 14 – общий вид пыльцевого зерна с экватора. 5, 10, 15 – скульптура поверхности. 1, 2, 6, 7, 11, 12 – СМ; 3–5, 8–10, 13–15 – СЕМ. Для каждого пыльцевого зерна показан верхний и нижний фокус (CМ). Масштабные линейки, мкм: 1–4, 6–9, 11–14 – 10; 5, 10, 15 – 1.
Plate III . Pollen morphology of the Nicotiana species (LM, SEM).
1–5 – N. noctiflora var. noctiflora; 6–10 – N. clevelandii; 11–15 – N. acuminata. 1, 3, 6, 8, 11, 13 – polar view of pollen grain. 2, 4, 7, 9, 12, 14 – equatorial view of pollen grain. 5, 10, 15 – exine ornamentation. 1, 2, 6, 7, 11, 12 – LM; 3–5, 8–10, 13–15 – SEM. The upper and lower foci are given for each pollen grain (LM). Scale bars, μm: 1–4, 6–9, 11–14 – 10; 5, 10, 15 – 1.
Анализ изображений поверхности пыльцевых зерен (СЭМ) позволил выделить 2 основных варианта скульптуры: сетчато-струйчатую, с короткими неориентированными струйками, переплетенными в нерегулярную сетку и перфорациями между струйками (табл. I, 7), и струйчатую, с короткими, иногда очень короткими плотно лежащими струйками, ориентированными в одном направлении (наиболее ярко выражена у пыльцы N. аcuminata, табл. III, 15). Однако, четко распределить изученные виды по этим группам оказалось затруднительно из-за большого числа переходных вариантов скульптуры у пыльцы разных видов (табл. I, 15; табл. II, 5).
Ряды форм пыльцы, индивидуальная изменчивость.
В образцах 8 из 16 изученных видов кроме 3-бороздно-оровой пыльцы, были обнаружены разнообразные атипичные формы пыльцевых зерен, отклоняющиеся от типичной по числу и расположению апертур (табл. IV). Были обнаружены следующие атипичные формы: циклоапертурная с кольцевой апертурой, лежащей в экваториальной плоскости (табл. IV, 1; рис. 1, А) или слегка согнутой, субэкваториальной (табл. IV, 2; рис. 1 Б, В); циклоапертурная, с кольцевой апертурой, закрученной как шов на теннисном мяче (табл. IV, 3; рис. 1, Ж); 4-бороздно-оровая с W-образно попарно скошенными апертурами (табл. IV, 4); 4-бороздно-оровая с меридионально ориентированными бороздами (табл. IV, 7, 8; рис. 1, К); 4-бороздно-оровая с W-образно попарно скошенными апертурами и двумя накрест ориентированными дополнительными ругами на противоположных полюсах (табл. IV, 9; рис. 1, З); 3-бороздно-оровая с ассиметричным расположением борозд, промежуточным между 3-бороздной и 4-бороздной W-формой (табл. IV, 5; рис. 1, Е) 6-руговая с апертурами, расположенными по ребрам тетраэдра (табл. IV, 6, 10–12; рис. 1, И).
Рис. 1.
Схематическое изображение морфологического ряда форм расположения апертур пыльцы (а) и распределение форм по числу видов рода Nicotiana (б).
а – последовательность форм расположения апертур (А–К). А – циклоапертурная форма с кольцевой апертурой, лежащей в плоскости экватора пыльцевого зерна; Д – 3-бороздная форма (типичная); Ж – цикло-апертурная форма с кольцевой апертурой, закрученной, как шов на теннисном мяче (уменьшенное изображение внизу справа – 4-бороздная форма с W-образно попарно скошенными бороздами); И – 6-руговая тетраэдр-форма; К – 4-бороздная форма с меридиональными бороздами. Уменьшенные изображения внизу справа от форм Г, Д, Ж, И и К показывают эту форму в другой проекции. Б, В, Г, Е, З – промежуточные формы.
б – цифры на диаграмме указывают число видов, в которых обнаружена обозначенная форма.
Fig. 1. Schematic view of the pollen form series (a) and distribution of the forms among the studied Nicotiana species (б).
а – pollen form series (А–К). А – cycloaperturate form with flat circular aperture situated in equatorial plate of pollen grain; Д – 3-colpate (typical) form; Ж – cycloaperturate form with circular aperture twisted like a seam on a tennis ball (smaller image to the lower right of the form is 4-colpate form with W-arranged colpi); И – 6-rugate tetrahedron form; К – 4-colpate form with meridian colpi. Smaler images to the lower right of the forms Г, Д, Ж, И and К depict these forms in another projection. Б, В, Г, Е, З – intermediate forms.
б – the figures on the diagram indicate the number of the species possessing the deviant forms.
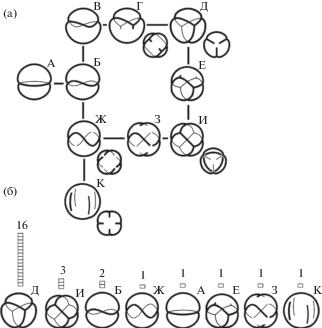
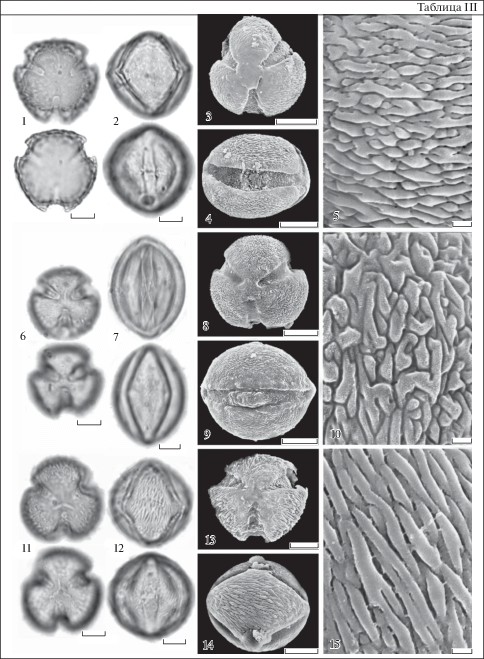
Таблица IV. Атипичные пыльцевые зерна представителей рода Nicotiana (СМ).
1 – N. bonariensis; 2 – N. acuminata; 3, 4 – N. noctiflora var. noctiflora; 5, 6 – N. alata; 7, 8 – N. tabacum; 9, 10 – N. attenuata; 11 – N. tabacum; 12 – N. noctiflora var. noctiflora. 1 – форма А; 2 – форма Б; 3, 4 – форма Ж; 5 – форма Е; 6, 10–12 – форма И; 7, 8 – форма К; 9 – форма З. Буквенные обозначения форм см. на рис. 1. Для каждого пыльцевого зерна показан верхний и нижний фокус. Масштабные линейки, мкм: 1–12 – 10.
Plate IV . Atypical pollens of the Nicotiana species (LM).
1 – N. bonariensis; 2 – N. acuminata; 3, 4 – N. noctiflora var. noctiflora; 5, 6 – N. alata; 7, 8 – N. tabacum; 9, 10 – N. attenuata; 11 – N. tabacum; 12 – N. noctiflora var. noctiflora. 1 – form А; 2 – form Б; 3, 4 – form Ж; 5 – form Е; 6, 10–12 – form И; 7, 8 – form К; 9 – form З. For the letter symbols of the pollen forms see Fig. 1. The upper and lower foci are given for each pollen grain. Scale bars, μm: 1–12 – 10.
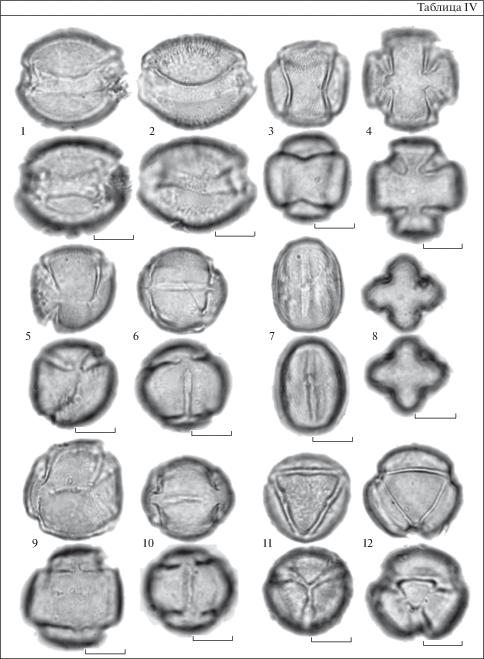
Оказалось, что изображенные в некоторой стандартной проекции, все формы (и отклоняющиеся, и типичная) могут быть упорядочены в единую непрерывную последовательность (рис. 1), в которой расположение борозд изменяется плавно и постепенно, строго подчиняясь некоей простой геометрической закономерности. Существование геометрической закономерности расположения апертур пыльцы Nicotiana описано впервые. Индивидуальная изменчивость расположения апертур неожиданно, с теоретической точки зрения, оказывается высоко упорядоченной, а форма всех отклоняющихся форм геометрически закономерной (все отклонения, как бы криво и ассиметрично они ни выглядели, подчиняются одной той же геометрической упорядоченности).
Хотя отклоняющиеся формы пыльцы во всех образцах Nicotiana редки и даже единичны, само это явление (пыльца с отклоняющейся формой расположения апертур) широко распространено у цветковых. Отклоняющиеся формы были обнаружены в 8 из 16 образцов исследованных видов.
Обнаруженные у изученных видов Nicotiana отклоняющиеся формы пыльцы (рис. 1) и полный параллелизм их рядов были ранее описаны для многих неродственных таксонов цветковых растений: Acer (Pozhidaev, 1993, fig, 3), Aesculus (Pozhidaev, 1995, Fig, 7), Scrophularia (Sheludyakova et al., 2017, рис. 1) и многих других (Pozhidaev, 1998).
Описанный ряд форм пыльцы, обнаруженный у изученных видов рода Nicotiana и существующий во многих других таксонах, является фрагментом целостного (непрерывного и упорядоченного) многообразия (Pozhidaev, 2015, рис. 1, 2 ; табл. 1), в котором плавными переходами удается объединить практически все типы расположения апертур (полярно-субполярные, экваториальные, меридиональные, глобальные) и их формы (типичные и отклоняющиеся), существующие у современных цветковых. Закономерности многообразия форм пыльцы оказываются в высокой степени транзитивны (таксон-неспецифичны и рангово независимы), а само многообразие симметричным и упорядоченым периодически (Pozhidaev, 2015).
Однако в подавляющем большинстве палиноморфологических исследований конкретных таксонов существование многообразных отклоняющихся форм пыльцы и упорядоченность их рядов обычно напрочь игнорируется авторами и вовсе не фиксируется в публикациях, либо подробно не описывается. Эти атипичные отклоняющиеся формы (несмотря на упорядоченность их многообразия, очевидную при корректном изучении) рассматриваются как уродства, терраты, нарушения, случайные аберрации, мутации, аномалии, по-видимому, в силу того, что эти отклонения всегда редки, выглядят необычно, несимметричны и устойчиво не наследуются. Наиболее же распространенные, типичные формы рассматриваются как физиологически нормальные, наследуемые и адаптивные, а их признаки как существенные, таксоноспецифичные, диагностические (сущностные); остальная изменчивость объявляется случайной и несущественной (акциденции). В результате такой типологической редукции создается искаженное представление о естественном полиморфизме признаков пыльцы в изучаемых таксонах, после чего содержательное обсуждение причин возникновения наблюдаемых свойств индивидуальной изменчивости не представляется возможным и все сводится к перечислению разнообразных частных случаев, не складывающихся ни в какую общую картину. Для причинного объяснения наблюдаемого полиморфизма необходимо более тщательное описание свойств естественного многообразия, интерпретация которых наталкивается на значительные теоретические трудности (Pozhidaev, 2015).
Вариабельность формы пыльцы Nicotiana.
Вариабельность пыльцы по числу и расположению апертур у отдельных видов Nicotiana (Mignot et al., 1994; Till-Bottraud et al., 1995) и некоторых других родов из семейства Solanaceae (Ferguson, Colidg, 1932; Henry, 1958; Airapetyan, 1991 и др.) описана в литературе.
J.W. Gish (2000) отмечал высокую степень изменчивости формы пыльцы у двух изученных им видов N. tabacum, N. rustica L. В образцах этих видов наряду с 3-бороздными пыльцевыми зернами встречается большое число 4-бороздно-оровых зерен и зерен с числом апертур, по определению автора, 4+2. Такое многообразие J.W. Gish считает следствием окультуривания этих двух видов табака, отмечая, что у изученных им диких видов подобного многообразия не было выявлено.
Для ряда видов было показано, что число атипичных пыльцевых зерен коррелирует с плоидностью растения, их продуцирующего. Диплоидные растения производят меньше атипичных зерен по сравнению с полиплоидными. Например, у растений N. tabacum особи 2n продуцируют 63.65% 3-апертурной пыльцы и 35.98% 4-апертурной, а растения 6n всего 5.03% 3-апертурных и 90.08% 4-апертурных пыльцевых зерен. Сходная закономерность характерна и для N. sylvestris Speg.& Comes, и для Petunia hybrida Vilmorin ( Mignot et al., 1994).
Анализ пыльцы 8 сортов табака, полученных в результате скрещивания N. sylvestris и N. tomentosiformis L. (Till-Bottraud et al., 1995), показал, что большинство гибридов продуцирует 3- и 4-апертурную пыльцу, тогда как родительские формы только 3-бороздную. При этом существует значительное различие в пропорциях этих двух форм пыльцы среди сортов, в пределах сорта между растениями и даже пыльниками одного цветка. Авторы рассматривают такой гетероморфизм пыльцы как следствие полиплоидизации.
У Physalis pruinosa L. в препаратах пыльцы аутотетраплоидных растений зафиксировано 6% 4-апертурной пыльцы, тогда как у контрольных диплоидных образцов вся пыльца 3-апертурная (Henry, 1958). Сходные данные приводятся и для видов Petunia (Ferguson, Colidg, 1932). Связь между плоидностью и ростом числа пыльцевых зерен с большим по сравнению с типичными числом апертур отмечали и в других семействах цветковых растений. Например, у полиплоидов трех видов Trifolium (Fabaceae) преобладает пыльца с числом апертур больше 3 (Najčevska, Speckmann, 1968). У тетраплоидной Oenothera (Onagraceae) преобладают 4-апертурные пыльцевые зерна, а у гексаплоидной – 5-апертурные, тогда как диплоидные виды продуцируют 3-апертурную пыльцу (Laws, 1965).
Однако, подобная закономерность свойственна не всем таксономическим группам. Так диплоидные растения из рода Viola продуцируют 3- и 4-апертурные или 4- и 5‑апертурные пыльцевые зерна (в зависимости от вида). По мнению авторов, преобладание тех или других зерен связано с условиями произрастания популяций. Также было показано, что пыльца фиалок с большим числом апертур прорастает быстрее, но при этом и быстрее теряет жизнеспособность, пыльца с меньшим числом апертур прорастает медленнее, но дольше сохраняет способность к прорастанию (Dajoz et al, 1991, 1993; Mignot et al., 1994: Till-Bottraud et al., 1999), что, по мнению авторов, обеспечивает разные стратегии выживания.
Также существует предположение, что техногенные загрязнения окружающей среды, в том числе повышенный уровень радиации, приводят к появлению и увеличению числа аномальных (в том числе по признаку числа и расположения апертур) пыльцевых зерен у ряда древесных и травянистых растений (Dzyuba, 2006; Dzyuba et al., 2006; Vasilevskaya, Petrova, 2014). Для Arabidopsis thaliana (L.) Heynh. описаны 3 мутантные линии (Prieu et al., 2016), которые характеризуются разным, отличным от дикого вида типом пыльцы (с безапертурными пыльцевыми зернами, с преобладанием 4-бороздной пыльцы и небольшим числом 3-бороздных зерен и мутант, продуцирующий одновременно 6- и 8-апертупные пыльцевые зерна).
Таким образом, причины появления в одном образце (пыльнике, цветке или растении) пыльцевых зерен c атипичным числом и расположением апертур пока остаются не ясны.
Таксономический анализ вариабельности признаков пыльцы представителей рода Nicotiana.
Анализ палиноморфологических данных показал, что такие признаки пыльцы, как геометрическая форма пыльцевого зерна, очертания и размер ор, особенности скульптуры могут служить дополнительными признаками для характеристики отдельных видов рода Nicotiana. Полученные нами данные не позволяют согласиться с мнением, что признаки пыльцы мало эффективны для систематики Nicotiana на видовом уровне (Collao-Alvarado et al., 2016). Авторы, на наш взгляд, недооценили такой признак как скульптура поверхности. Хотя даже на опубликованных фотографиях пыльцы, сделанных при небольших увеличениях СЭМ, характерный облик и отличия пыльцы разных видов очевидны.
Полученные палиноморфологические данные были сопоставлены с систематическим делением рода Nicotiana по T.H. Googspeed (1954). Подрод Rustica включает 3 секции: Paniculatae (7 видов), Thyrsiflorae (1 вид), Rusticae (1 вид). Нами изучена пыльца трех видов из секции Paniculatae (N. benavidesii, N. glauca, N. paniculata) и одного вида из секции Rusticae (N. rustica). У пыльцы всех изученных видов из секции Paniculatae длина полярной оси почти равна длине экваториального диаметра, зерна почти сфероидальные, самые мелкие из исследованных видов, со сходным очертанием мезокольпиумов. Пыльца всех трех видов хорошо различается характером скульптуры, а N. paniculata очень крупными, четко очерченными орами. Пыльца N. rustica отличается от видов секции эллипсоидальной формой, большими размерами. По характеру скульптуры секция неоднородна.
Подрод Tabacum включает 2 секции: Tomentosae (5 видов), Genuinae (1 вид). Нами изучена пыльца одного вида N. tabacum из монотипной секции Genuinae. Пыльца этого вида гетероморфна, наряду с типичными 3-бороздно-оровыми пыльцевыми зернами встречается пыльца с другим числом и расположением апертур. По характеру скульптуры пыльца N. tabacum похожа на пыльцу N. benavidesii и N. rustica из подрода Rustica. Межвидовые различия хорошо видны при исследовании с помощью свето-оптического микроскопа. Пыльца N. tabacum отличается эллипсоидальной формой (соотношение P/E значительно больше 1) и более четкими бороздами.
Подрод Petunioides – самый большой в роде, включает 45 видов, разделенных на 6 секций: Undulatae, Trigonophyllae, Alatae, Noctiflorae, Acuminatae, Suaveolentes. Изучена пыльца 8 видов из 4 секций: Acuminatae (2), Alatae (3), Bigelovianae (2), Noctiflorae (1). Пыльца всех изученных видов из этого подрода характеризуется струйчатой скульптурой поверхности. Характер стручатости (длина, ширина струй, их форма, расположение) являются хорошими диагностическими признаками для разграничения большинства изученных видов. Однако сходный струйчатый характер скульптуры имеют пыльцевые зерна N. paniculata из подрода Rusticae. Пыльца других изученных видов этого подрода по характеру скульптуры сходна с пыльцой N. tabacum из подрода Tabacum. Признаки пыльцы в разных секциях подрода Petunioides перекрываются.
Таким образом, сравнительное исследование морфологии пыльцевых зерен видов рода Nicotiana показало, что по комплексу морфологических признаков пыльцы не удается выделить группы, совпадающие с установленными по макроморфологическим признакам подродам и секциям. При этом данные палинологии могут быть использованы для характеристики отдельных видов или групп видов Nicotiana.
Список литературы
[Airapetyan] Айрапетян А.М. 1991. Апертурный полиморфизм пыльцы у вида Physalis maxima L. (Solanaceae). – Флора, растительность и растительные ресурсы Армении. 13: 107–115.
Borisjuk N.V., Davidjuk Y.M., Kostishin S.S., Miroshnichenco G.P., Velasco R., Hemleben V., Volkov R.A. 1997. Structural analysis of rDNA in the genus Nicotiana. – Plant Molecular Biology. 35 (5): 655–660.
Bryant V.M., Kampbell S.M., Hall J.L. 2012. Tobacco pollen: archaeological and forensic applications. – Palynology. 36 (2): 208–223.
Chase M.W., Knapp S., Cox A., Clarkson J., Butsko Y., Joseph J., Savolainon V., Parokonny A.S. 2003. Molecular Systematics, GISH and the Origin of Hybrid Taxa in Nicotiana (Solanaceae). – Ann. Bot. 92 (7): 107–127.
Clarkson J.J., Knapp S., Garcia V.F., Olmstead R.G., Leitch A.R. 2004. Phylogenetic relationships in Nicotiana (Solanaceae) inferred from multiple plastid DNA regions. – M. Phylogenet. Evol. 33 (1): 75–90.
Dajoz I., Till-Bottraud I., Gouyon P.H. 1991. Evolution of pollen morphology. – Science. 253 (5015): 66–68.
Dajoz I., Till-Bottraud I., Gouyon P.H. 1993. Pollen aperture polymorphism and gametophyte performance in Viola diversifolia. – Evolution. 47 (4): 1080–1093.
[Dzyuba O. F] Дзюба О.Ф. 2006. Палиноиндикация качества окружающей среды. СПб. 197 с.
Dzyuba O.F., Shrekova O.V., Tokarev P.I. 2006. On the natural polymorphism of pollen grains of Acer tataricum L. – Paleont. Journ. 40 (5): 590–594.
Erdtman G. 1952. Pollen morphology and taxonomy. Angiosperms. Stockholm. 539 p.
Ferguson M.C., Coolidge E.B. 1932. A cytological and a genetical study of Petunia. IV. Pollen grains and method of studying them. – Amer. J. Bot. 19 (7): 644–658.
Finot V.L., Marticorena C., Marticorena A. 2018. Pollen grain morphology of Nolana L. (Solanaceae: Nolanoideae: Nolaneae) and related genera of southern South American Solanaceae. – Grana. https://doi.org/10.1080/00173134.2018.1458897
Googspeed T.H. 1947. On the evolution of the genus Nicotiana. – Proc. Natt. Acad. Sci. USA. 33 (6): 15–171.
Googspeed T.H. 1954. The genus Nicotiana – Chronica Botanica. Waltham. Massachusetts. 536 p.
Goodward M.B.E., Pell F.L.S., Pell K. 1994. Inheritance of exine pattern in Nicotiana sanderae (Solanaceae). – Bot. J. Lin. Soc. 115 (2): 145–159.
Gish J.W. 2000. Morphological distinctiveness of Nicotiana pollen and the potential for identifying prehistoric Southwest tobacco use through pollen analysis. – In: Tobacco use by native north Americans: Sacred smoke and silent killer. – Norman. P. 223–261.
Henry D.R. 1958. Morphological studies of diploid and autotetraploid plants of Physalis pruinosa L.: PhD Thesis. Ohio. 90 p.
Holloway R.G., Dean G. 2000. Morphological studies of New Mexico Solanaceae pollen. – In: Winter J. C. Tabacco use by native North Americans: sacred smoke and silent killer. – Norman. P. 211–222.
[Kupriyanova, Aleshina] Куприянова Л.А., Алешина Л.А. 1967. Палинологическая терминология покрытосеменных растений. Л. 84 с.
[Kupriyanova, Aleshina] Куприянова Л.А., Алешина Л.А. 1972. Пыльца и споры растений флоры европейской части СССР. Т. 1. Л. С. 48–51.
Laws H.M. 1965. Pollen grain morphology of polyploid Oenotheras. – J. Heredity. 56 (1): 18–21.
Leopold E.B. 1969. Miocene pollen and spore flora of Eniwetok Atoll, Marshall Islands. Washington. P. 1144.
Mignot A., Hoss C., Dajoz I., Leuret C., Henry J.-P., Dreuillaux J.-M., Heberle-Bors E., Till-Bottraud I. 1994. Pollen aperture polymorphism in the Angiosperms: importance, possible causes and. – Acta Bot. Gallica. 14 (2): 109–122.
Najčevska C.M., Speckmann G.J. 1968. Nambers of chloroplasts and pollen grain pores in diploid and tetraploid varieties of some Trifolium species. – Euphytica. 17 (3): 357–362.
Perveen A., Qaiser M. 2007. Pollen morphology of family Solanaceae from Pakistan. – Pak. J. Bot. 39 (7): 2243–2256.
Pozhidaev A.E. 1993. Polymorphism of pollen in the genus Acer (Aceraceae). Isomorphism of deviant forms of Angiosperm pollen. – Grana. 32 (2): 79–85.
Pozhidaev A.E. 1995. Pollen morphology of the genus Aesculus (Hippocastanaceae). Patterns in the variety of morphological characteristics. – Grana. 34 (1): 10–20.
Pozhidaev A.E. 1998. Hypothetical way of pollen aperture patterning. 1. Formation of 3-colpate patterns and endoaperture geometry. – Rev. Palaeobot. Palynol. 104 (1): 67–83.
[Pozhidaev] Пожидаев А.Е. 2015. Рефренная структура биологического многообразия и теория филогенеза. Палеоботанический временник. Приложение к журналу “Lethaea rossica Российский палеоботанический журнал”. 2: 115–127.
Prieu C., Matamoro-Vidal A., Raquin C., Dobritsa A., Mercier R., Gouyon R., Albert B. 2016. Aperture number influences pollen survival in Arabidopsis mutants. – Amer. J. Bot. 103 (3): 452–459.
Särkinen T., Bohs L., Olmstead R.G., Knapp S. 2013. A phylogenetic framework for evolutionary study of the nightshades (Solanaceae): a dated 1000-tip tree. – BMC Evolutionary Biology. 13:214. http://www.biomedcentral.com/1471-2148/13/214
[Sheludyakova, Grigoryeva, Pozhidaev] Шелудякова М.Б., Григорьева В.В., Пожидаев А.Е. 2017. Морфология пыльцевых зерен представителей рода Scrophularia (Scrophulariaceae). – Бот. журн. 102 (3): 361–379.
Ślasarczyk J., Tykarska T., Wierzbicki A., Jerzmanowski A., Kuraś M. 2006. Tapetum development in transgenic tobacco (Nicotiana tabacum L.) plants with modified level of histone H1 variants.– Acta soc.bot. pol. 75 (2): 95–105.
Tiii-Bottraud I, Gouyon P. H., Dajoz I.,Venabie D.L. 1991. Evolution of pollen morphology: game theory models. – Am. Nat. 144 (3): 395–411.
Till-Bottraud I., Mignot A., de Paepe R., Dajoz I. 1995. Pollen heteromorphism in Nicotiana tabacum (Solanaceae). – Amer. J. Bot. 82 (8): 1040–1048.
Till-Bottraud I., Vincent M., Dajoz I., Mignot A. 1999. Pollen aperture heteromorphism:
Variation in pollen type proportions along altitudinal transects in Viola calcarata (Violaceae).
Comptes Rendus de l’Académie des Sciences Paris. Life Sciences. 322 (7): 579–589.
[Vasilevskaya, Petrova] Василевская Н.В., Петрова Н.В. 2014. Морфологическая изменчивость пыльцы Pinus sylvestris L. в условиях промышленного города (на примере Мончегорска). – Уч. записки Петрозаводского гос. ун-та. 141 (4): 7–12.
Warner B.G. 2012. Paleoecology of floating bogs and landscape change in the Great Lakes drainage basin of North America. – In: Climate change and human impact on the Lundscape . Kelle. P. 238–246.
Дополнительные материалы отсутствуют.
Инструменты
Ботанический журнал