

Ботанический журнал, 2019, T. 104, № 7, стр. 1033-1044
БЕСШОВНЫЕ ДИАТОМОВЫЕ ПРИБРЕЖНОЙ ЗОНЫ БЕРЕЗОВЫХ ОСТРОВОВ ФИНСКОГО ЗАЛИВА
В. А. Степанова *
Ботанический институт им. В.Л. Комарова РАН
197376 С.-Петербург, ул. Проф. Попова, 2, Россия
* E-mail: vera.stepanova@binran.ru
Поступила в редакцию 15.04.2019
После доработки 24.04.2019
Принята к публикации 16.05.2019
Аннотация
В прибрежной зоне ООПТ Березовые острова (Финский залив) было идентифицировано 15 видов из 8 родов бентосных бесшовных диатомовых водорослей. Представлены данные о морфологии и распространении видов на побережье, приведены микрофотографии. Вид Opephora krumbeinii указан впервые для Ленинградской области, Stauroforma inermis – впервые для флоры России и Балтийского моря; все виды впервые приводятся для Березовых остров.
Региональный комплексный заказник “Березовые острова” – одна из крупнейших особо охраняемых природных территорий Ленинградской обл. (Выборгский р-он): с 1976 г. в составе Выборгского заказника, с 1996 г. выделен в отдельный ООПТ. Заказник расположен в северо-восточной части Финского залива Балтийского моря, в 2 км к юго-западу от г. Приморска и включает в себя более 50 островов и 46 000 га акватории. В состав архипелага входят три крупных острова (Большой Березовый, Западный Березовый и Северный Березовый), 23 более мелких острова и более 30 безымянных. Точное число островов неизвестно, поскольку большинство из них являются небольшими каменистыми отмелями, появление которых зависит от колебания уровня моря (Prirodnaya…, 2007).
В данном районе Финского залива проводятся не только разнообразные гидрохимические и гидрологические исследования, но и изучение флоры и фауны, как самих островов, так и прибрежных вод. Но, исследования бентосных диатомовых прибрежной зоны ООПТ ранее никогда не проводились.
Воды Финского залива, где расположены Березовые острова, относятся к солоноватым. Этот район достаточно мелководен, в центральной части глубины варьируют от 30 до 45 м, около берегов – значительно мельче (до 5 м). Соленость поверхностных вод колеблется от 0.5 до 4.0 ‰ (Prirodnaya …, 2007), непосредственно в районе островов значения солености изменяются в зависимости от глубины, направления ветра и времени года, в интервале 2–4 ‰ (Luk’yanov et al., 2003).
МАТЕРИАЛ И МЕТОДЫ ИССЛЕДОВАНИЙ
Материалом для настоящего исследования послужили 66 проб, собранных в августе 2015 г. в прибрежной зоне островов Равица, Волчий, Большой Солнечный, Цепной, Северный Березовый, Западный Березовый и Большой Березовый. Одновременно со сбором материала измерялись температура, соленость и рН воды при помощи кондуктометра HM Digital COM80 и Checker pH Tester HI 98103. В период сбора материала показатели среды в районе исследований составляли: соленость – 1.9–2.2‰, водородный показатель – 7.3–9.6 рН, температура – 19.3–23.9°С., (табл. 1; “BO” – “Berezovye Ostrova”, места сбора проб в ООПТ). Пробы диатомовых собирались на глубине от 20 см до 1.5 м на мелководье с различных субстратов: поверхности грунтов (песок, илистый грунт), камней, деревянных и иных погруженных в воду искусственных сооружений (помосты, сваи, канаты), а также пробы с водных растений и водорослей-макрофитов. Фиксировались пробы 40% раствором формалина.
Предварительный просмотр проб осуществлялся с помощью светового микроскопа Carl Zeiss Jena Amplival. Для изучения морфологии диатомовых водорослей панцири очищали методом холодного сжигания (Balonov, 1975). Полученный осадок с очищенными створками диатомовых помещался в 90% спирт для дальнейшего хранения. Осадок с отмытыми створками использовался для приготовления препаратов для светового и электронного микроскопов. Для определения водорослей использовали сканирующий электронный микроскоп JSM 6390LA (Jeol). Обилие в пробах оценивалось визуально: “единично” – единичные клетки или створки; “редко” – до нескольких десятков клеток в препарате; “часто” – до нескольких сотен клеток в препарате. Названия таксонов высокого ранга даны по системе F.E. Round, R.M. Crawford и D.G. Mann (Round et al., 1990) и E.J. Cox (Cox, 2015). Используемая терминология соответствует “Русскому и английскому терминологическому словарю по морфологии” (Gogorev et al., 2016).
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
Литературные сведения о разнообразии диатомовых водорослей как пресноводных водоемов, так и прибрежной зоны Березовых островов отсутствуют. Всего было выявлено 80 видов диатомовых водорослей из трех классов – Coscinodiscophyceae, Fragilariophyceae и Bacillariophyceae. Особенностью сообществ диатомовых обрастаний является преобладание бесшовных пеннатных диатомовых (Fragilariophyceae) – 15 видов из 8 родов: Ctenophora Grunow ex D.M. Williams et Round, Diatoma Bory, Fragilaria Lyngbye, Opephora P. Petit, Pseudostaurosira D M. Williams et Round, Stauroforma Flower, Jones et Round, Tabularia Kutzing ex D.M. Williams et Round, Ulnaria Compere. Наиболее часто встречаются виды: Ctenophora pulchella, Diatoma moniliformis, Fragilaria vaucheriae и Tabularia fasciculata. Ниже приведены описания и иллюстрации таксонов бесшовных пеннатных диатомовых водорослей. Дана экологическая и географическая характеристика видов, отмечено их распространение в Балтийском море, местонахождения в ООПТ Березовые острова, частота встречаемости.
Класс Fragilariophyceae Round 1990
Порядок Fragilariales Silva 1962
Семейство Fragilariaceae Greville 1833
Ctenophora pulchella (Ralfs ex Kützing) D.M. Williams et Round 1986, Diatom Research, Vol. 1 (2), p. 330, figs 53–61 . (табл. I: 1, 2). Створки от линейных до ланцетных со слабоголовчатыми концами, более длинные створки с более выраженными головчатыми концами, 37–171 мкм дл., 4.5–6.8 мкм шир. Осевое поле узкое, линейное, слегка эксцентричное, как правило, слабозаметное. Среднее поле в виде отчетливой фасции, от округлого до прямоугольного, в единичных случаях эксцентрично расположенное, на внутренней поверхности обычно с центральной депрессией. Редко присутствуют теневые штрихи, или фасция односторонняя. Апикальные поровые поля на внешней поверхности небольшие, типа оцеллюлимбус, располагаются на каждом конце створки. Штрихи однорядные, параллельные, слаборадиальные к концам, заходящие на загиб створки, 13–20 в 10 мкм; штрихи обычно укороченные вокруг среднего поля и у концов створки; ареолы в штрихах от округлых до прямоугольных, вытянутых в апикальном направлении, с лицевой поверхности прикрыты велумом (простыми пороидными пластинками). Две римопортулы на створку.
Таблица I. Ctenophora pulchella (1, 2), Diatoma moniliformis (3, 4), D. tenuis (5, 6), D. problematica (7, 8), Fragilaria capucina (9, 10), F. vaucheriae (11, 12).
1, 3, 5, 7, 9, 11 – створка; 2, 4, 6, 8, 10, 12 – створка с внутренней поверхности. Масштабные линейки, мкм: 1, 5, 7 – 5; 2, 6 – 10; 3, 8–10 – 2; 4, 11, 12 – 1.
Plate I . Ctenophora pulchella (1, 2), Diatoma moniliformis (3, 4), D. tenuis (5, 6), D. problematica (7, 8), Fragilaria capucina (9, 10), F. vaucheriae (11, 12).
1, 3, 5, 7, 9, 11 – valve; 2, 4, 6, 8, 10, 12 – valve interior. Scale bars, μm: 1, 5, 7 – 5; 2, 6 – 10; 3, 8–10 – 2; 4, 11, 12 – 1.
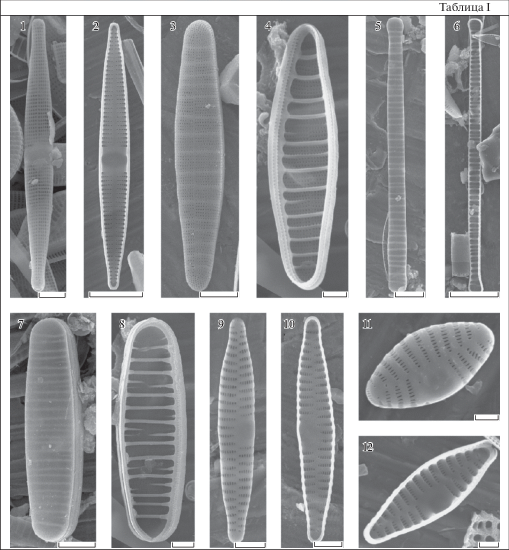
Солоноватоводный эпифитный вид; космополит. В Балтийском море: почти повсеместно (Intercalibration…, 1993; Potapova, 1999; Hällfors, 2004; Balashova et al., 2016). BO-03, BO-04, BO-06, BO-12, BO-16, BO-20, BO-21. Часто.
Diatoma moniliformis (Kützing) D.M. Williams 2012, Nova Hedwigia, Vol. 141, p. 260, figs 3–5 . (Табл. I: 3, 4). Створки от эллиптических до ланцетных, концы от широко закругленных до головчатых, 6.9–60.8 мкм дл., 2.2–7.3 шир., соотношение длины и ширины 2.3–21.7. Осевое поле узкое, линейное, слегка эксцентричное, как правило, отчетливое. Апикальные поровые поля располагаются на загибе створки на обоих концах, заходят на лицевую поверхность. Штрихи однорядные, продолжаются на загиб створки, 43–78 в 10 мкм; иногда не строго параллельны, а слегка искривлены относительно поперечной оси; ареолы в штрихах мелкие округлые. Римопортула одна или две на створку. Широкие первичные ребра располагаются на внутренней поверхности створки, 6–14 в 10 мкм, иногда присутствуют вторичные ребра.
Пресноводно-солоноватоводный эпифитный вид, аркто-бореально-тропический. В Балтийском море: почти повсеместно (Intercalibration…, 1993; Potapova, 1999; Hällfors, 2004; Balashova et al., 2016). BO-04-06, BO-08, BO-10, BO-12, BO-13, BO-15, BO-16, BO-19-21. Часто.
Diatoma problematica Lange-Bertalot 1993, Bibliotheca Diatomologica, Vol. 27, pl. 6, figs 11–15, 21–27 ; pl. 7, figs 1–6 ; pl. 8, figs 1–5 . (Табл. I: 7, 8). Створки линейные, иногда слаболанцетные, с концами от широко закругленных до головчатых, 21–35 мкм дл., 4.4–7 мкм шир., соотношение длины к ширине 3.4–7.6. Осевое поле узкое, линейное, слегка эксцентричное, как правило, отчетливое. Апикальные поровые поля располагаются на обоих концах створки. Штрихи однорядные, продолжаются на загибе створки, 47–64 в 10 мкм; ареолы в штрихах мелкие округлые. Римопортула одна на створку. Первичные ребра на внутренней поверхности створки широкие, 7–14 в 10 мкм, иногда присутствуют вторичные ребра.
Пресноводный эпифитный вид; бореальный (европейский). Для Балтийского моря ранее отмечался в Финском заливе (Potapova, 1999). BO-06, BO-13, BO-16, BO-20. Редко.
Diatoma tenuis Agardh 1812, Algarum decas prima, р. 15. (Табл. I: 5, 6). Створки линейные, концы головчатые, 25.6–51 мкм дл., 3–6.9 мкм шир., соотношение длины и ширины 3.6–13.2. Осевое поле узкое, слегка эксцентричное, как правило, отчетливое. Апикальные поровые поля располагаются на обоих концах створки. Штрихи однорядные, не строго параллельные друг другу, продолжаются на загиб створки, 41–49 в 10 мкм; ареолы в штрихах мелкие округлые. Римопортула одна на створку. Первичные ребра 7–11 в 10 мкм, вторичные ребра иногда присутствуют.
Солоноватоводно-морской, эпифитный вид; аркто-бореально-тропический. В Балтийском море: повсеместно (Intercalibration…, 1993; Potapova, 1999; Hällfors, 2004; Balashova et al., 2016). BO-06, BO-13, BO-15, BO-16, BO-20. Редко.
Fragilaria capucina Desmazières 1830, Plantes Cryptogames du nord de la France, p. 200, tab. 4 , icon 3, No. 453. (Табл. I: 9, 10). Створки линейные, линейно-ланцетные, с оттянутыми, головчатыми концами, 25–31.6 мкм дл., 3.2–4.2 мкм шир. Осевое поле узкое, линейное, слегка эксцентричное. Среднее поле прямоугольное, образованное укороченными штрихами с одной стороны створки, типа фасция. Апикальные поровые поля небольшие, округло-прямоугольные. Штрихи однорядные, чередующиеся, 15–18 в 10 мкм; ареолы в штрихах от округлых до овальных, вытянутых в апикальном направлении. На концах створки могут располагаться несколько округлых пор. Римопортула одна на створку.
Пресноводный бентосный вид; бореальный (европейский). В Балтийском море: повсеместно (Intercalibration…, 1994; Potapova, 1999; Hällfors, 2004; Balashova et al., 2016). BO-03, BO-05, BO-19-21. Единично.
Fragilaria vaucheriae (Kützing) J. B. Petersen 1938, Botaniska Notiser, p. 167, fig. 1a–g . (Табл. I: 11, 12). Створки линейно-эллиптические, с закругленными или слегка оттянутыми концами, 6.1–42.7 мкм дл., 1.9–4.6 мкм шир. Осевое поле узкое, линейное, слегка эксцентричное. Среднее поле одностороннее, типа субфасция. Апикальные поровые поля с внешней поверхности небольшие, округло-прямоугольные. Штрихи однорядные, чередующиеся, 11–24 в 10 мкм; ареолы от округлых до овальных. Римопортула одна на створку.
Пресноводный вид, эпифит, эпилит; аркто-бореально-тропический. В Балтийском море: почти повсеместно (Intercalibration…, 1995; Potapova, 1999; Hällfors, 2004; Balashova et al., 2016). BO-03-05, BO-10, BO-16, BO-19-21. Часто.
Opephora krumbeinii A. Witkowski, Witak et K. Stachura in H. Lange-Bertalot et S. I. Genkal 1999, Iconographia Diatomologica. Vol. 6, p. 80, pl. 3: figs 1–17 , pl. 4: figs 1–3. (Табл. II: 1–3). Створки округлые, 2.1–3.8 мкм дл., 1.8–3.7 мкм шир. Апикальные поровые поля состоят из нескольких (1–3) округлых или вытянутых пор. Штрихи однорядные, параллельные или слегка радиальные, 20–30 в 10 мкм; ареолы округло-прямоугольные. Римопортула отсутствует. На границе загиба и лицевой поверхности створки на штрихах располагаются удлиненные шипы, иногда шипы отсутствуют.
Таблица II. Opephora krumbeinii (1–3), O. mutabilis (4–6), Pseudostaurosira brevistriata (7), P. trainorii (8–10), Stauroforma atomus (11, 12).
1, 4, 7, 8, 11 – створка; 9, 12 – створка с внутренней поверхности; 2, 3, 5, 6, 10 – панцирь, вид с пояска. Масштабные линейки, мкм: 1 – 0.5; 2–4, 6–12 – 1; 5 – 2.
Plate II . Opephora krumbeinii (1–3), O. mutabilis (4–6), Pseudostaurosira brevistriata (7), P. trainorii (8–10), Stauroforma atomus (11, 12).
1, 4, 7, 8, 11 – valve; 9, 12 – valve interior; 2, 3, 5, 6, 10 – frustule, girdle view. Scale bars, μm: 1 – 0.5; 2–4, 6–12 – 1; 5 – 2.

Морской бентосный вид, эпилит; бореальный (североамериканско-европейский). В Балтийском море: Гданьский залив (Lange-Bertalot, Genkal, 1999). BO-05, BO-06, BO-13, BO-15, BO-16, BO-20. Часто.
Opephora mutabilis (Grunow) Sabbe et Wyverman 1995, European Journal of Phycology, Vol. 30, p. 241, figs 13–28 . (Табл. II: 4–6). Створки гетеропольные, с зауженным базальным концом, 9.6–11.2 мкм дл., 3.4–4.7 мкм шир. Осевое поле узкое. Штрихи однорядные, чередующиеся, слаборадиальные, 7–10 в 10 мкм, состоят из нескольких апикально вытянутых ареол. На каждом конце створки располагается апикальное поровое поле различной формы и структуры. Римопортула отсутствует. На границе лицевой поверхности и загиба створки на штрихах расположены крупные плоские лопастевидные шипы.
Примечание: M.D. Guiry и G.M. Guiry (Guiry, Guiry, 2019) указывают, что базионим названия вида недействительно обнародован, поскольку, по их мнению, в первоописании отсутствует диагноз на латинском языке. “Invalid: proposed basionym invalid [INA]. Invalid in its own right: lacking Latin description – (12 Jan 2018) – M.D. Guiry”. K. Sabbe и W. Vyverman (Sabbe, Wyverman, 1995) отмечают, что P.T. Cleve и J.D. Möller (Cleve, Möller, 1879) опубликовали список видов образца No. 255, в этом списке содержится следующая надпись: “Sceptroneis (marina var. ?) mutabilis Grunow (Fragilaria mut. form. 10 str. in 0.01 mm) common, var. minuta Grunow. 15 str. in 0.01 mm”. По нашему мнению, такого описания достаточно, чтобы считать название вида действительно обнародованным.
Солоноватоводно-морской вид, эпифит, эпипсаммит; космополит. В Балтийском море: Рижский и Финский заливы, южная Балтика (Intercalibration…, 1993; Hällfors, 2004; Balashova et al., 2016). BO-13, BO-16. Единично.
Pseudostaurosira brevistriata (Grunow) D.M. Williams et Round 1988, Diatom Research, Vol. 2, p. 276, figs 28–31 . (Табл. II: 7). Створки ланцетные, с закругленными концами. дл. от 7.4 мкм, шир. до 3.9 мкм. Апикальное поровое поле прямоугольное, расположено на каждом конце створки. Штрихи короткие, состоят из двух овальных ареол, 16 в 10 мкм. Римопортула отсутствует. Шипы разветвленные, полые, расположены по краю створки на штрихах.
Пресноводно-солоноватоводный вид, эпифит; космополит. В Балтийском море: Финский и Рижский заливы, южная Балтика (Intercalibration…, 1998; Hällfors, 2004). BO-13, BO-15, BO-16, BO-20. Единично.
Pseudostaurosira trainorii E.A. Morales 2001, Proceedings of the Academy of Natural Sciences of Philadelphia, Vol. 151, p. 113, fig. 6 a–l . (Табл. II: 8–10). Створки от круглых до эллиптических, 3.8–4.8 мкм дл., 3.6–4.3 мкм шир. Осевое поле от узкого до широколанцетного. Апикальные поровые поля отсутствуют. Штрихи однорядные прямые, 15–18 в 10 мкм, состоят из нескольких круглых или округло-прямоугольных, крупных ареол. Римопортула отсутствует. На границе поверхности и загиба створки располагается ряд шипов на штрихах. Шипы неразветвленные, иногда слегка уплощенные на концах.
Примечание: Н.Б. Балашова (Balashova et al., 2016) отмечает сходство P. trainorii с видом P. elliptica (Schumann) Edlund, Morales et Spaulding, в результате чего может быть перепутан с последним. Однако стоит отметить, что P. trainorii, хоть и обладает сходными размерными параметрами, имеет значительные отличия по структуре панциря. Одним из важных диагностических признаков является расположение шипов на штрихах у P. trainorii, в то время как у вида P. elliptica шипы расположены между штрихами, кроме того, у последнего шипы полые, чего не наблюдается у P. trainorii, все эти параметры можно оценить только с использованием СЭМ.
Пресноводный бентосный вид; бореальный (североамерикано-европейский). В Балтийском море отмечался в Финском заливе (Balashova et al., 2016). BO-03, BO-08, BO-12, BO-15, BO-16, BO-20. Редко.
Stauroforma atomus (Hustedt) D. Talgatti, C.E. Wetzel, E. Morales et L.C. Torgan in Talgatti et al. 2014, Phytotaxa, Vol. 158(1), p. 45. (Табл. II: 11, 12). Створки от эллиптических до ланцетных, изо- или гетеропольные, с широко закругленными, иногда тупо усеченными концами, 3.3–5.2 мкм дл., 2.7–3.5 мкм шир. Осевое поле узкое, линейное или слегка искривленное, может быть расположено эксцентрично. Апикальные поровые поля состоят из вертикальных рядов округлых пор. Штрихи однорядные, преимущественно параллельные в центре и слегка радиальные к концам, 20–26 в 10 мкм; ареолы от округлых до эллиптических. Шипы и римопортула отсутствуют.
Солоноватоводный вид, эпипсаммон; бореальный (европейский). В Балтийском море: Каттегат, Ботнический, Рижский и Финский заливы (Intercalibration…, 1993; Potapova, 1999; Hällfors, 2004). BO-03, BO-05, BO-10, BO-13, BO-16, BO-19, BO-20. Часто.
Stauroforma inermis R.J. Flower, V.J. Jones et Round 1996, Diatom Research, Vol. 11(1), p. 54, figs 23–33 . (Табл. III: 1–3). Створки от эллиптических до ланцетных, с тупо-, иногда узкозакругленными концами, 6.7–19.6 мкм дл., 4–5 мкм шир. Осевое поле узкое, линейное. Апикальное поровое поле расположено на каждом конце створки. Штрихи однорядные, параллельные или слегка радиальные, 19–21 в 10 мкм, ареолы в штрихах мелкие, от округлых до овальных, вытянутых апикально. Римопортула и шипы отсутствуют.
Таблица III. Stauroforma inermis (1–3), Tabularia fasciculata (4–7), T. tabulata (8–12), Ulnaria ulna (13, 14).
1, 2, 4, 10, 13 – створка; 3, 5, 11 – створка с внутренней поверхности; 6, 8 – середина створки с наружной поверхности; 7, 9 – кончик створки с внутренней поверхности; 12 – увеличенный фрагмент створки с внутренней поверхности, штрихи из макроареол, закрытых велумом; 14 – кончик створки с внешней поверхности. Масштабные линейки, мкм: 1 – 2; 2, 3, 6–9, 12, 14 – 1; 4, 5 – 10; 10, 11, 13 – 20.
Plate III. Stauroforma inermis (1–3), Tabularia fasciculata (4–7), T. tabulata (8–12), Ulnaria ulna (13, 14).
1, 2, 4, 10, 13 – valve; 3, 5, 11 – valve interior; 6, 8 – middle part of valve exterior; 7, 9 – valve apex interior; 12 – enlarged fragment of valve interior, striae with macroareolae closed by velum; 14 – valve apex exterior. Scale bars, μm: 1 – 2; 2, 3, 6–9, 12, 14 – 1; 4, 5 – 10; 10, 11, 13 – 20.

Примечание: под световым микроскопом этот вид легко перепутать с S. exiguiformis (Lange-Bertalot) R.J. Flower, V.J. Jones et Round, поскольку размерные характеристики видов перекрываются. Основные различия заключаются в наличии или отсутствии шипов и апикальных поровых полей. У вида S. exiguiformis шипы на поверхности створки присутствуют, но нет поровых полей; S. inermis отличается обратным: шипов нет, а поровые поля развиты на обоих концах створки.
Пресноводный бентосный вид; североамерикано-антарктический. В Балтийском море ранее не отмечался. BO-05, BO-13, BO-16. Редко.
Tabularia fasciculata (Agardh) D.M. Williams et Round 1986, Diatom Research, Vol. 1(2), p. 326, figs 46–52 . (Табл. III: 4–7). Створки линейно-ланцетные, ланцетные, со слегка головчатыми концами, 37–96 мкм дл., 3.4–6.3 мкм шир. Осевое поле широколанцетное, отчетливое. Апикальные поровые поля типа оцеллюлимбус. Штрихи однорядные, короткие, параллельные, 11–17 в 10 мкм, форма ареол в штрихах меняется от апикально вытянутых у края створки до более мелких полуэллиптических вблизи осевого поля. Две римопортулы на створку. На концах створки рядом с римопортулой располагается небольшая группа округлых мелких пор, продолжающихся на загиб створки.
Эвригалинный вид, эпифит, бентос; аркто-бореально-тропический. В Балтийском море: почти повсеместно (Intercalibration…, 1993; Hällfors, 2004; Balashova et al., 2016). BO-03, BO-05, BO-10, BO-13, BO-16, BO-19, BO-20. Часто.
Tabularia tabulata (Agardh) Snoeijs 1992, Diatom Research, Vol. 7(2), р. 343, figs 38–48 . (Табл. III: 8–12). Створки линейно-ланцетные, ланцетные, с широкозакругленными концами, 124–225.8 мкм дл., 5.6–10.6 мкм шир. Осевое поле широкое, слегка ланцетное, отчетливое. Апикальные поровые поля типа оцеллюлимбус. Штрихи однорядные, короткие, параллельные, 12–14 в 10 мкм, состоят из макроареол (Табл. III: 12), переходящих на загиб створки с разрывом на границе лицевой поверхности и загиба. Ареолы вытянуты в трансапикальном направлении. Римопортул две на створку. На концах створки рядом с римопортулой располагаются небольшие группы округлых мелких пор, продолжающихся на загиб створки.
Примечание: M.D. Guiry (Guiry, Guiry, 2019) вид T. tabulata рассматривают в качестве синонима T. fasciculata. Мы придерживаемся мнения, что эти виды самостоятельны. Во-первых, T. tabulata значительно отличается от T. fasciculata по размеру: длина 100–540 мкм против 22–176 мкм, ширина 6–9 мкм против 4.5–8 мкм. Кроме того, существенно различается структура штрихов: у T. fasciculata они состоят из апикально вытянутых ареол, разделенных тонкими виминами (Табл. III: 6), у T. tabulata штрихи, за исключением нескольких апикальных, состоят из макроареол (Табл. III: 8, 12). Также различна форма створок: для T. fasciculata характерны оттянутые концы (Табл. III: 7), тогда как у T. tabulata они широкозакругленные (Табл. III: 9). Двугубые выросты у последнего крупные, у T. fasciculata – небольшие. P. Snoeijs (Snoeijs, 1992) также отмечает различие в форме римопортул: у вида T. tabulata выросты с внутренней стороны створки имеют прямоугольную форму, у T. fasciculata – овальную; в числе хлоропластов – 20–35 против 8–20, соответственно; и значениях солености, при которых оба вида были обнаружены в Балтийском море: T. tabulata при солености 4–7‰ и T. fasciculata при солености 5–>11‰.
Солоноватоводно-морской вид, эпифит бентос; бореально-тропически-антарктический. В Балтийском море: повсеместно (Intercalibration…, 1993; Hällfors, 2004). BO-03, BO-04, BO-13, BO-15. Часто.
Ulnaria ulna (Nitzsch) Compère in Jahn et al. 2001, Lange-Bertalot Festschrift. Studies on diatoms dedicated to Prof. Dr. Dr. h.c. Horst Lange-Bertalot on the occasion of his 65th birthday, р. 100. (Табл. III: 13,14). Створки линейные, с закругленными концами, 142–177 мкм дл., 5.2–6.8 мкм шир. Осевое поле узкое, линейное, слабозаметное. Среднее поле прямоугольное. Апикальные поровые поля крупные, округло-прямоугольные. Штрихи однорядные, параллельные, 9–16 в 10 мкм, ареолы от эллиптических, вытянутых апикально, до округлых. Римопортула на каждом конце створки. На краю загиба и лицевой поверхности кончика створки располагаются два шипика (Табл. III: 14).
Пресноводный вид, эпифит, планктон; космополит. В Балтийском море: повсеместно (Intercalibration…, 1995; Potapova, 1999; Hällfors, 2004; Balashova et al., 2016). BO-13. Единично.
Большинство обнаруженных видов широко распространены по всему Балтийскому морю, включая Финский залив, и характерны для опресненных участков Балтики. Вид Stauroforma atomus ранее был отмечен для пролива Каттегат, Ботнического, Финского и Рижского заливов (Intercalibration…, 1993), Pseudostaurosira trainorii – только для Финского залива (Balashova et al., 2016). Наибольший интерес представляет вид впервые указанный для Балтийского моря, – Stauroforma inermis; новым для флоры Ленинградской обл. является Opephora krumbeinii, для России – Stauroforma inermis.
Список литературы
[Balashova et al.] Балашова Н.Б., Киселев Г.А., Степанова В.А., Тобиас А.В. 2016. Диатомовые водоросли бентоса южного побережья Финского залива (заказник “Лебяжий”). – Вестник СПбГУ. Серия 3. Биология. 4: 9–25. https://doi.org/10.21638/11701/spbu03.2016.401
[Balonov] Балонов И.М. 1975. Подготовка водорослей к электронной микроскопии. – В кн.: Методика изучения биогеоценозов внутренних водоемов. М. С. 87–90.
Cleve P.T., Möller J.D. 1879. Diatoms. Uppsala: Esatas Edquists Boktrykeri. V. 217–276.
Cox E.J. 2015. Coscinodiscophyceae, Mediophyceae, Fragilariophyceae, Bacillariophyceae (Diatoms). Syllabus of Plant Families. Adolf Engler’s Syllabus der Pflanzenfamilien. Part 2/1. Photoautotrophic eukaryotic Algae: Glaucocystophyta, Cryptophyta, Dinophyta/Dinozoa, Haptophyta, Heterokontophyta/Ochrophyta, Chlorarachniophyta/Cercozoa, Euglenophyta/Euglenozoa, Chlorophyta, Streptophyta. Berlin. P. 64–103.
[Gogorev et al.] Гогорев Р.М., Чудаев Д.А., Степанова В.А., Куликовский М.С. 2018. Русский и английский терминологический словарь по морфологии диатомовых водорослей. – Новости систематики низших растений. 52(2): 265–309. https://doi.org/10.31111/nsnr/2018.52.2.265
Guiry M.D., Guiry G.M. 2019. AlgaeBase. World-wide electronic publication, National University of Ireland. Galway. http://www.algaebase.org (08.04.2019).
Hällfors G. 2004. Checklist of Baltic Sea Phytoplankton Species (including some heterotrophic protistan groups). Baltic Sea Environment Proceedings No. 95. Helcom. Helsinki. 208 p.
Intercalibration and distribution of diatom species in the Baltic Sea. (eds. Snoeijs P.) 1993. The Baltic Marine Biologist Publication. N 16a. Opulus Press Uppsala. Vol. 1. 129 p.
Intercalibration and distribution of diatom species in the Baltic Sea. (eds. Snoeijs P., Vilbaste S.) 1994. The Baltic Marine Biologist Publication. N 16b. Opulus Press Uppsala. Vol. 2. 126 p.
Intercalibration and distribution of diatom species in the Baltic Sea. (eds. Snoeijs P., Potapova M.) 1995. The Baltic Marine Biologist Publication. N 16c. Opulus Press Uppsala. Vol. 3. 126 p.
Intercalibration and distribution of diatom species in the Baltic Sea. (eds. Snoeijs P., Balashova N.) 1998. The Baltic Marine Biologist Publication. N 16e. Opulus Press Uppsala. Vol. 5. 144 p.
Lange-Bertalot H., Genkal S.I. 1999. Diatoms from Siberia I – Islands in the Arctic Ocean (Yugorsky‑Shar Strait). In: Lange-Bertalot, H. (ed.) Iconographia Diatomologica. Annotated Diatom Micrographs. Vol. 6. Phytogeography-Diversity-Taxonomy. Königstein. 303 p.
[Luk’yanov et al.] Лукьянов С.В., Пнюшков А.В., Степанов О.В. 2003. Распределение гидрологических показателей по акватории пролива Бьёркёзунд. – В кн.: Природная среда побережья и акватории Финского залива (район порта “Приморск”). СПб. С. 81–86.
[Potapova] Потапова М.Г. 1999. Диатомовые водоросли песчаной литорали в восточной части Финского залива. – Бот. журн. 84 (1): 59–65.
[Prirodnaya…] Природная среда и биологическое разнообразие архипелага Березовые острова (Финский залив). 2007. СПб. 368 с.
Round F.E., Crawford R.M., Mann D.G. 1990. The diatoms. Biology and morphology of the genera. Cambridge. 747 p.
Sabbe K., Vyverman W. 1995. Taxonomy, morphology and ecology of some widespread representatives of the diatom genus Opephora. – European Journal of Phycology. 30: 235–249.
Snoeijs P. J. M. 1992. Studies in the Tabularia fasciculata complex. – Diat om Research. 7 (2): 313–344.
Дополнительные материалы отсутствуют.
Инструменты
Ботанический журнал