

Ботанический журнал, 2019, T. 104, № 7, стр. 1045-1058
ПИРОГЕННЫЕ СУКЦЕССИИ В МОХОВОМ ЯРУСЕ НА МЕЗОТРОФНОМ БОЛОТЕ В ПРИАМУРЬЕ
Т. А. Копотева *
Институт водных и экологических проблем ДВО РАН
680022 г. Хабаровск, ул. Ким Ю Чена, 65, Россия
* E-mail: kopoteva@ivep.as.khb.ru
Поступила в редакцию 19.04.2019
После доработки 29.04.2019
Принята к публикации 16.05.2019
Аннотация
Приведены результаты десятилетних наблюдений за изменениями видового состава и динамики фитомассы мхов после катастрофического пожара на торфяном болоте. Исследования проводились методами геоботанических описаний и отбора с последующей обработкой монолитов на выгоревшем до торфа участке и в акротельме негоревшего участка с ненарушенной дерниной. Выявлены закономерности развития сукцессии Polytrichum strictum. Сделано заключение влияния повторных палов на процесс восстановления сфагнового покрова. Впервые приведены данные фитомассы и продукции Polytrichum strictum и влияние на них пирогенного стресса.
Растительный покров большинства торфяных болот северного полушария – самоорганизующаяся эндогенно развивающаяся экосистема и эта ее способность определяется сфагновыми мхами, благодаря их эдификаторной роли. В условиях Приамурья начальная фаза постпирогенной сукцессии на торфяных болотах является детерминантной (Kopoteva, Kuptsova, 2016а), потому что с самого начала доминируют терминальные виды, являвшиеся доминантами и до пожара (Titlyanova et al., 1993). Но это относится только к кустарничково-травяному ярусу, сукцессии мохообразных развиваются по своим закономерностям и они изучены очень слабо. Мхи – исключительно чувствительный компонент, хорошо отражающий даже небольшие изменения физико-химических условий среды, которые часто не проявляются на уровне сосудистых растений (Proktor, 1982; Schofield, 1985). Мхи способны к репарации фотосинтетической деятельности после весенних и осенних заморозков и сухих периодов летом, что свидетельствует о высокой устойчивости фотосинтетического аппарата, и позволяет им занимать заметное место в сообществах (Shpak, 2008). Сфагновые же мхи относятся к “ключевым видам” (эдификаторам) мезо-олиготрофных болот, так как создают среду, в которой снижаются темпы минерализации опада сосудистых растений и сами по себе являются главным материалом в процессе торфообразования, что доказано в экспериментах. Поэтому считается, что восстановление мезо-олиготрофной болотной экосистемы не может рассматриваться как успешное, до тех пор, пока не восстановлено растительное сообщество с доминированием сфагновых мхов. Но продуктивность сфагновых мхов находится в сильной зависимости от гидрологического режима в летний период. По данным наших исследований доля продукции сфагновых мхов от общей продукции мезотрофных болотных фитоценозов Приамурья колеблется от 8 до 57%. На мезотрофных болотах Европейской части России эти колебания находятся в пределах 30–34% (Elina et al., 1984). В засушливые годы в Приамурье сфагновые мхи образуют лишь 7–9% от собственной фитомассы. Во влажные годы их продукция достигает 90–96% от запасов своей фитомассы (3.2–4.6 т/га). Доля сфагновых мхов в надземной фитомассе периодически выгораемых мезотрофных болот Приамурья варьирует от 45 до 62% (Kopoteva, 1999). Таким образом, они являются самым важным и одновременно наиболее уязвимым компонентом растительного покрова торфяных болот.
В последнее время проявляется большой интерес исследователей к проблеме восстановления напочвенного покрова с моховым ярусом после пожаров и палов в лесных экосистемах, как наиболее уязвимого компонента растительного покрова. Появился целый ряд публикаций, описывающих результаты наблюдений восстановления после пожаров, в том числе после экспериментальных контролируемых выжиганий (Gorshkov et al., 1995; Perevosnikova et al., 2007; Sofronov et al., 2008; Kovaleva et al., 2011; Kovaleva, Ivanova, 2013; Volokitina et al., 2013; Kovaleva, 2015; Ivanova et al., 2017). Результатов же исследований восстановления мохового яруса после пожаров на торфяных болотах и пирогенных сукцессий мхов на торфе крайне мало (Popov, 2000; Kopoteva, 2008; Kosykh, 2011). Между тем, уничтожение мохового покрова приводит к нарушению основной биосферной функции торфяных болот, в результате чего они перестают какое-то время быть стоком углерода (Naumov et al., 2009). По результатам исследований состояния верховых “островных” болот лесостепи Западной Сибири пирогенный фактор играет здесь ключевую роль. Деградация болота проявляется в виде разрастания кустарников и кустарничков, снижения скорости накопления торфа вследствие выгорания сфагновых мхов (Naumov et al., 2009). Те же процессы мы наблюдаем и в наших исследованиях (Kopoteva, Kuptsova, 2016 а).
Мезотрофные болота Приамурья так же, как и на юге Западной Сибири, занимают в регионе южный форпост распространения и тоже находятся в условиях неустойчивого увлажнения. Хотя годовое количество осадков в Приамурье в 2 раза больше, высокая частота повторяемости пожаров (из-за контрастности увлажнения) наносит процессам торфообразования невосполнимый ущерб. Пирогенный фактор в Приамурье считается ландшафтообразующим (Kopoteva, 1999; Burenina, 2006). Крупные лесные пожары, при которых выгорают и болота, за последние десятилетия наблюдались с периодичностью 8 лет (Sokolova et al., 2009). Для сравнения: рассчитанный для заболоченных лиственничников Средней Сибири зоны северной тайги межпожарный интервал составляет от 73 ± 8 до 138 ± 18 лет при ведущей роли природной составляющей в возникновении пожаров (Kharuk et al., 2005).
Все эти данные заставляют задуматься о пересмотре концепции, рассматривающей торфяные болота северного полушария, как абсолютный сток углерода (O’Neyll, 2002). Поскольку многократное воздействие пожаров может трансформировать экосистемы в источники углерода не только за счет прямых его выбросов при сгорании биомассы и косвенных воздействий пожаров на тепловой и водный режим, но и меняет структуру и функционирование экосистем.
Целью нашей работы было: проследить изменения видового состава и динамику фитомассы мхов на начальном этапе пирогенной вторичной сукцессии и выявить особенности восстановления сфагнового покрова в условиях Приамурья.
МАТЕРИАЛЫ И МЕТОДЫ
Изучение хода восстановления растительного покрова проводилось в течение 10 лет на Бичевском болотном массиве (47°48' с.ш., 135°39' в.д.), типичном для Среднеамурской низменности, на котором в середине июня 2008 г., в конце засушливого периода, прошел пожар. До пожара доминантами растительного покрова были: Chamaedaphne calyculata + Ledum palustre + Carex globularis + Sphagnum fuscum. Пожаром было уничтожено более 80% растительного покрова на площади болота, в том числе древесный ярус, состоявший из угнетенной лиственницы Vа класса бонитета. Однако из-за быстрого прохождения огня при сильных порывах ветра на торфяных болотах остаются отдельные редкие неповрежденные огнем участки (Kopoteva, Kuptsova, 2016 b). Прогорание было мозаичным, в основном на глубину сфагнового очеса на 5–10 см в понижениях и 20–30 см на подушках, местами выгорел торф. На пройденных огнем участках были полностью уничтожены: надземные живая фитомасса и мортмасса кустарничково-травяного яруса, живая фитомасса и значительная часть мортмассы (очеса) мохового яруса, обнажились кочки пушицы влагалищной, ранее скрытые моховым покровом, погибли возобновление и подрост лиственницы, 95% взрослых деревьев. На первом этапе восстановления происходило уплотнение верхнего торфяного слоя и сохранившегося сфагнового очеса, их проседание и подъем уровня болотных вод. Это отмечалось после катастрофических пожаров на торфяных болотах и другими исследователями (Tsaregradskaja, Kositsin, 1999).
Моховый ярус до пожара состоял из сфагновых мхов с очень небольшой примесью видов рода Polytrichum. Проективное покрытие (ПП) мохового яруса составляло до пожара 80–90%, в микрорельефе преобладали подушки и ковры, образованные Sphagnum fuscum (Schimp.) Klinggr., с небольшой примесью S. divinum11 Flatb. et K. Hassel. Склоны подушек в смешанных со S. divinum дернинах занимал S. balticum (Russ.) Russ. ex C. Jens., днища понижений были заняты только S. balticum. Более подробное описание растительности болотного фитоценоза до пожара приводится в более ранних наших работах (Kopoteva, Kuptsova, 2016 а, 2016 б).
Торфяная залежь мощностью 2–2.5 м низинно-переходного типа имеет типичное для мезотрофных болот Среднеамурской низменности строение, сложена, главным образом, травяно-древесно-кустарничковыми, кустарничково-травяными торфами, в основном низинными, лишь в верхней части переходными (Kopoteva, 2011).
С 2008 по 2018 г. проводились наблюдения за восстановлением растительного покрова на выгоревших и ненарушенных пожаром (сохранившихся) участках болота. В апреле 2014 г. прошел повторный пал слабой интенсивности, так как моховый ярус был насыщен водой и местами под снегом. Обгорели кустарнички и ветошь травянистых на тех же площадях, которые пострадали от первого пожара, мхи пострадали незначительно. В июне 2018 г. прошел еще один пал. Поскольку уровень болотных вод был ниже, выгорели не только надземные части кустарничков и ветошь, но и значительная часть верхушек моховых дернин, в основном политриховых. Оба пала захватили только территорию уже пройденного пожаром 2008 г. массива болота. Ненарушенные первым пожаром участки были слегка задеты палом 2018 г. только по краям, растительный покров на них не пострадал.
Восстановительные сукцессии бриофитов изучались на 10 учетных площадках, заложенных на обоих участках случайным методом: каждая площадка размером 1 м2 после удаления растений кустарничково-травяного яруса зарисовывалась в масштабе, отмечались видимые на обнаженном торфе мхи и в камеральных условиях определялся процент проективного покрытия по видам. Учитывая высокую степень мозаичности растительности и контагиозности распределения возобновляющихся видов, проводилось также описание 50 площадок по 1 м2 на трансекте с глазомерной оценкой ПП бриофитов без удаления растений кустарничково-травяного яруса. До 2012 г. наблюдения проводились только за надземной частью фитоценоза. Для наблюдений за динамикой процессов в акротельме с 2012 г. вырезались монолиты (10 × 10 см) на глубину 30 см в количестве 10 повторностей на уцелевшем от огня участке и выгоревшем. В камеральных условиях монолиты разбирались, определялась фитомасса мхов в абсолютно-сухом состоянии, у политриховых отделялся годичный прирост – пигментированная фотосинтезирующая часть (Korchagin, 1960; Goncharova, 2008) и боковые столоновидные побеги (Ignatov, Ignatova, 2003). Живая фитомасса Polytrichum strictum Brid. приравнена к годичному приросту. У S. fuscum она рассчитана для длины побега 5 см от вершины головки, у S. divinum – 10 см (Malysheva, 1970).
РЕЗУЛЬТАТЫ И ИХ ОБСУЖДЕНИЕ
В первый год восстановления (2009) количество атмосферных осадков теплого периода (апрель-октябрь) – 693 мм – было в пределах среднемноголетней нормы (685 мм). Последующие 2 года были влажными: на 38 и 13% больше нормы, что отчасти предотвратило внедрение сосудистых растений-эксплерентов и создало благоприятные условия для восстановления растений мохового яруса. В это время на поверхности голого торфа активно развивалась сукцессия Marchantia polymorpha L. и мелких бриевых мхов: Ceratodon purpureus (Hedw.) Brid., и Cynodontium strumiferum (Hedw.) Lindb. Эти мхи занимали более половины поверхности обнаженного торфа (табл. 1).
Таблица 1.
Динамика восстановления растительного покрова после пожара, ПП, % Table 1. Dynamics of recovering of the vegetation cover after a fire, projective cover, %
| Год Year |
Голый торф
с мелкими бриевыми Bare peat with small Bryidae |
Polytrichum strictum | Sphagnum divinum | S. fuscum | S. balticum + + S. fallax + + S. capilifolium | Фитомасса сосудистых растений, г/м2 Phytomass of vascular plants, g/m2 |
|---|---|---|---|---|---|---|
| 2009 | 89.5 ± 2.9 | 12.5 ± 3.4 | 1.4 ± 0.5 | 0.6 ± 0.5 | 0 | 151.0 ± 25.0 |
| 2010 | 61.2 ± 8.1 | 26.8 ± 3.6 | 13.0 ± 5.0 | 2.3 ± 1.9 | 0 | 275.0 ± 26 |
| 2011 | 49.6 ± 5.5 | 27.1 ± 7.6 | 9.5 ± 2.5 | 8.5 ± 4.4 | 5.9 ± 1.6 | 419.0 ± 49 |
| 2012 | 52.3 ± 6.4 | 25.0 ± 7.3 | 11.7 ± 4.1 | 9.7 ± 2.3 | 1.3 ± 0.5 | 456.7 ± 23.0 |
| 2013 | 42.8 ± 5.5 | 17.7 ± 3.2 | 27.2 ± 5.0 | 4.0 ± 2.8 | 7.8 ± 2.6 | 685.2 ± 64.0 |
| 2014 | 71.2 ± 6.4 | 10.7 ± 1.8 | 15.9 ± 5.1 | 0.9 ± 0.5 | 1.3 ± 0.7 | 312.8 ± 29.9 |
| 2015 | 41.7 ± 9.1 | 2.0 ± 1.4 | 35.4 ± 6.1 | 8.4 ± 4.5 | 11.5 ± 5.6 | 378.6 ± 27.0 |
| 2016 | 23.0 ± 6.8 | 16.9 ± 6.0 | 45.0 ± 7.6 | 10.3 ± 2.3 | 4.8 ± 3.0 | 313.0 ± 20.9 |
| 2017 | 23.5 ± 8.5 | 26.9 ± 6.6 | 31.7 ± 6.1 | 14.3 ± 4.3 | 4.2 ± 3.6 | 346.43 ± 29.7 |
| 2018 | 24.5 ± 8.3 | $\frac{{10.8 \pm 3.3{\text{*}}}}{{12.7 \pm 5.4}}$ | $\frac{{27.2 \pm 7.6{\text{*}}}}{{20.8 \pm 6.7}}$ | $\frac{{1.5 \pm 0.7{\text{*}}}}{{5.0 \pm 3.1}}$ | 0 | 258.5 ± 20.4 |
Развивались споровые поселения политриховых, главным образом P. strictum, значительно менее распространены P. commune Hedw. и P. piliferum Hedw. Переувлажнение этих двух лет, по-видимому, способствовало сокращению времени течения сукцессии мелких бриевых: их ПП на голом торфе снизилось с 40% ± 2.6 в 2010 г., до 13% ± 1.4 в 2011 г. При обработке монолитов в 2012 г. выделялось 51.1 ± 13.8 г/м2 их вещества, в 2013 г. – 101.8 ± 33.6 г/м2, а в 2014 – 46.9 ± 35.0 г/м2 соответственно, в 2015 г. их уже почти не было. Таким образом, длительность существования сукцессии мелких бриевых мхов после пожара на торфе не более 5 лет. Споровые возобновления политриховых происходили прямо на сплошном покрытии торфа мелкими бриевыми мхами. Развитие сукцессии P. strictum на обнаженном торфе началось раньше, чем на негоревшем участке с ненарушенным сфагновым покровом. Уже в 2012 г. была одна полностью сформировавшаяся дернина (0–30 см) из 10 повторностей и 3 активно формирующиеся. В 2013 г. тоже одна, возраст растений 4–5 лет, но количество фитомассы в поверхностном покрове снизилось (табл. 2). В 2014 г. тоже только одна дернина и три на стадии активного формирования. В 2015 г. фитомасса P. strictum очень мала, в 2016 г. опять увеличение: 2 сформировавшиеся и 4 формирующиеся дернины. В конце 2017 г. мы фиксировали уже 8 полностью сформированных дернин из десяти повторностей (табл. 2). После пала 2018 г. в дернинах в горизонте 0–10 см начали активно расти боковые столоновидные побеги: нитевидные образования 6–8 см длиной розового цвета. Поскольку поверхности дернин обгорели, и обычный для этих растений верхушечный рост стал невозможен, политриховые мхи таким образом пытаются восстановить дернину (табл. 3). Как видно из данных, приведенных в таблице, на развитие сукцессии P. strictum июньский пал 2018 г. оказал очень сильное влияние: на ненарушенном участке оно положительное, на горевшем ранее пока отрицательное, т.к. обгорели дернины и стал невозможен верхушечный годичный прирост, но в дальнейшем сукцессия может возобновиться с прежней силой.
Таблица 2.
Динамика фитомассы мхов, абс.-сух. вес, г/м2 Table 2. Dynamics of phytomass of mosses, absolute dry weight, g/m2
| Фитомасса/Phytomass | 2012 | 2013 | 2014 | 2015 | 2016 | 2017 | 2018 |
|---|---|---|---|---|---|---|---|
| Ненарушенный участок Undisturbed site |
|||||||
| Sphagnum fuscum живой (0–5 см) S. fuscum alive (0–5 cm) |
289.0 ± 22.9 | 227.9 ± 31.8 | 287.4 ± 30.3 | 342.9 ± 43.6 | 221.1 ± 38.7 | 181.5 ± 44.9 | 243.2 ± 37.8 |
| S. magellanicum живой (0–10 см) S. magellanicum alive (0–10 cm) |
149.1 ± 30.1 | 84.0 ± 14.1 | 90.3 ± 31.0 | 133.0 ± 46.0 | 171.3 ± 43.4 | 250.9 ± 70.5 | 189.1 ± 57.9 |
| S. balticum живой (0–10 см) S. balticum alive (0–10 cm) |
18.2 ± 11.1 | 0 | 2.0 ± 2.0 | 0 | 3.1 ± 3.1 | 7.1 ± 4.1 | 0 |
| Итого сфагновые Total Sphagnum |
456.3 ± 39.4 | 311.9 ± 34.8 | 379.7 ± 43.4 | 475.9 ± 63.4 | 395.5 ± 58.2 | 439.4 ± 83.7 | 432.3 ± 69.2 |
| Polytrichum strictum, 0–10 см P. strictum, 0–10 cm |
1.7 ± 1.6 | 263.6 ± 185.7 | 316.3 ± 161.2 | 334.9 ± 127.4* | 631.3 ± 118.8* | 415.7 ± 126.2 | 670.8 ± 156.3 |
| P. strictum, 0–30 см P. strictum, 0–30 см |
2.9 ± 2.3 | 387 ± 194.9 | 647.1 ± 307.8 | 1028.5 ± 250.1* | 1504.8 ± 335.7* | 1122.4 ± 315.4 | 1852.8 ± 437.6 |
| Выгоревший участок Burnt site |
|||||||
| S. fuscum (0–5 см) S. fuscum (0–5 cm) |
11.0 ± 8.5 | 33.5 ± 12.8 | 79.8 ± 40.1 | 92.6 ± 47.5 | 184.7 ± 72.2 | 254.2 ± 42.3 | 149.3 ± 64.0 |
| S. divinum (0–10 см) S. divinum (0–10 cm) |
26.5 ± 15.8 | 108.9 ± 43.3 | 123.5 ± 66.9 | 231.3 ± 73.1 | 265.6 ± 70.0 | 104.0 ± 24.0 | 195.4 ± 60.7 |
| S. balticum (0–10 см) S. balticum (0–10 сm) |
0 | 0 | 0 | 17.0 ± 12.8 | 2.8 ± 2.8 | 3.0 ± 3.0 | 0.4 ± 0.4 |
| Итого сфагновые Total Sphagnum |
37.5 ± 17.9 | 142.4 ± 45.2 | 203.3 ± 78.0 | 340.9 ± 88.1 | 453.1 ± 100.6 | 361.2 ± 49.5 | 345.1 ± 88.2 |
| P. strictum, 0–10 см P. strictum, 0–10 cm |
249.4 ± 130.1 | 100.0 ± 49.9 | 285.0 ± 101.7 | 7.9 ± 4.1 | 407.3 ± 121.3 | 364.4 ± 88.7 | 346.8 ± 126.1 |
| P. strictum, 0–30 см P. strictum, 0–30 см |
352.0 ± 142.2 | 125.5 ± 53.0 | 449.9 ± 196.7 | 36.4 ± 17.3 | 615.5 ± 200.9 | 969.7 ± 236.1 | 626.8 ± 126.4 |
Таблица 3.
Динамика годичного прироста (ANP) Polytrichum strictum (в дернинах), г/м2 Table 3. Dynamics of annual growth (ANP – annual net weight production) of Polytrichum strictum (in cushions), g/m2
| Фитомасса/Phytomass | 2016 | 2017 | 2018 |
|---|---|---|---|
| Ненарушенный участок/Undisturbed site | |||
| ANP верхушечный рост, г/м2 ANP apical growth, g/m2 |
130.8 ± 27.8 | 96.8 ± 34.5 | 201.1 ± 41.2 |
| Боковые столоны, г/м2 Lateral stolons, g/m2 |
0 | 0 | 0.6 ± 0.6 |
| Итого: Total: |
130.8 ± 27.8 | 96.8 ± 34.5 | 201.7 ± 41.3 |
| Доля ANP от своей фитомассы горизонта 0–10 см, % Proportion of ANP from its phytomass of layer 0–10 cm, % |
21 | 23 | 30 |
| Выгоревший участок/Burnt site | |||
| ANP верхушечный рост, г/м2 ANP apical growth, g/m2 |
101.2 ± 43.7 | 114.9 ± 32.7 | 31.1 ± 9.0 |
| Боковые столоны, г/м2 Lateral stolons, g/m2 |
0 | 0 | 11.2 ± 5.4 |
| Итого: Total: |
101.2 ± 43.7 | 114.9 ± 32.7 | 42.3 ± 10.5 |
| Доля ANP от своей фитомассы горизонта 0–10 см, % Proportion of ANP from its phytomass of layer 0–10 cm,% |
25 | 31 | 12 |
Распределение P. strictum по площади выгоревшего болота очень неравномерное, мозаичное, поэтому по запасам фитомассы трудно судить о причинах такой динамики (мало повторностей из-за большой трудоемкости обработки). Судя по всему развитие сукцессии P. strictum происходит очень быстро, отрицательное влияние оказывает конкуренция со стороны растений кустарничково-травяного яруса: их надземная фитомасса здесь в 2–2.5 раза превышает фитомассу ненарушенного участка (табл. 4). Также отрицательное влияние оказывают повторные палы: слабое в 2014 г. и очень сильное в 2018 г. Кроме того, сказываются, по-видимому, сезонные различия увлажнения: самые низкие показатели фитомассы в горизонте 0–10 см у него в холодном и влажном 2015 г., самые высокие в сухом 2016 г., при среднегодовой норме осадков 736 мм (рисунок 1, табл. 2).
Таблица 4.
Динамика долевого участия сфагновых мхов в составе живой надземной фитомассы негоревшего (А) и горевшего (Б) участков сообщества Table 4. Dynamics of proportion of the Sphagnum mosses in the composition of alive phytomass on undisturbed (A) and burnt (B) sites of community
| Год Year |
Живая фитомасса мхов, г/м2/ Phytomass of alive mosses, g/m2 |
Живая фитомасса сосудистых растений, г/м2/Phytomass of alive vascular plants, g/m2 | Доля мхов в надземной фитомассе, %/Proportion of mosses in underground phytomass, % | |||
|---|---|---|---|---|---|---|
| А | Б | А | Б | А | Б | |
| 2012 | 456.7 | 107.3** | 220.8 | 456.7 | $\frac{{67.3{\text{*}}}}{{0.05}}$ | $\frac{{6.6}}{{12.9}}$ |
| 2013 | 375.2 | 170.4** | 262.9 | 685.2 | $\frac{{48.9}}{{9.9}}$ | $\frac{{16.6}}{{3.3}}$ |
| 2014 | 452.4 | 283.1 | 221.0 | 312.8 | $\frac{{56.4}}{{10.8}}$ | $\frac{{34.1}}{{13.4}}$ |
| 2015 | 559.6 | 343.3 | 215.9 | 378.6 | $\frac{{63.3}}{{10.8}}$ | $\frac{{57.3}}{{0.4}}$ |
| 2016 | 526.3 | 554.3 | 166.0 | 313.0 | $\frac{{57.0}}{{19.0}}$ | $\frac{{52.2{\text{*}}}}{{11.7}}$ |
| 2017 | 536.2 | 476.1 | 108.7 | 347.1 | $\frac{{68.1}}{{15.0}}$ | $\frac{{43.9{\text{*}}}}{{14.0}}$ |
| 2018 | 634.0 | 387.4 | 126.6 | 258.5 | $\frac{{56.8}}{{26.5}}$ | $\frac{{53.4{\text{*}}}}{{6.5}}$ |
Примечание. * – над чертой доля живых сфагновых мхов, под чертой – только живые политриховые мхи. ** – не включена фитомасса мелких бриевых мхов.
Note. * – above the line: the proportion of alive Sphagnum mosses, below the line: proportion of alive Polytrichaceae mosses. ** – phytomass of small Bryidae mosses is not included.
Рис. 1.
Динамика годовых сумм осадков и сумм положительных температур по метеостанции Бичевая (Погода и климат. https:rp.5.ru)
Fig. 1. Variations in annual precipitation and annual positive cumulative temperatures at the Bichevaya settlement area (Weather and climate. https:rp.5.ru)
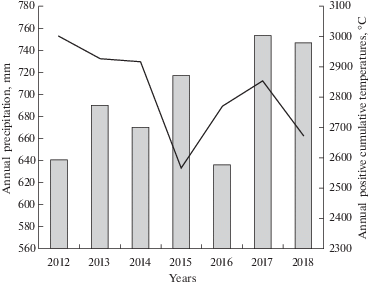
На ненарушенных пожаром участках болота сукцессия политриховых происходит иначе, чем на выгоревшей до торфа поверхности. Во первых, сукцессия P. strictum здесь начала развиваться позже: на пятый год после пожара (табл. 2). Во вторых, растения P. strictum здесь стали прорастать прямо в дернине S. fuscum, постепенно вытесняя доминанта. До пожара P. strictum отмечался в моховом покрове единично (Kopoteva, Kuptsova, 2016а). Еще в 2012 г. его фитомасса ничтожно мала (табл. 2), растения только начинают развиваться из проникших в дернину S. fuscum спор, к концу 2013 г. уже 2 полностью сформированные дернины. А к концу 2016 г. политриховые уже создают мощные дернины более 30 см в 9 из 10 повторностей и их фитомасса в горизонте 0–10 см практически равна общей фитомассе сфагновых в этом горизонте (табл. 2). К концу 2017 г. живая фитомасса (продукция) P. strictum на негоревшем участке немного снизилась (табл. 2, 3), видимо из-за переувлажнения (рисунок), а в 2018 г. – резко увеличилась, вероятно из-за влияния пала.
К концу десятого года наблюдений на ненарушенном участке P. strictum, постепенно накапливая фитомассу с 6.5 т/га, образовал около 26 т/га в акротельме горизонта 0–30 см. На выгоревшем от пожара 2008 г. участке болота его фитомасса в этом слое колебалась с 0.4 до 13 т/га. В отечественной литературе очень мало данных фитомассы и продукции политриховых. Мы нашли их только в работах С.Э. Вомперского c соавторами (Vomperski et el., 1999): фитомасса Polytrichum commune (255 г/м2 год–1) и О.В. Шпак (Shpak, 2008): годичный прирост того же вида (94 г/м2).
Восстановление сфагновых мхов на выгоревшем участке находится в сильной зависимости от глубины прогорания во время пожара и степени увлажнения в постпирогенный период. Возобновление, наряду со споровым, идет также вегетативно из сохранившейся живой ткани очеса, поэтому в первую очередь восстанавливаются мхи, имеющие наиболее длинный стебель с живой тканью. В частности, S. divinum здесь имеет явное преимущество перед S. fuscum на ранних этапах сукцессии. Первые поселения сфагновых мхов на торфе, в основном S. divinum, отмечены уже в 2009 г.: очень редкие головки на наименее обгоревшем очесе подушек. По данным геоботанических описаний в восстанавливающемся сфагновом покрове горевшего участка доля ПП S. divinum в 2012 г. составляла 51%, далее она увеличилась, колеблясь с 63 до 88% (табл. 1). Показатели и ПП, и фитомассы S. divinum за все время наблюдений значительно выше, чем у S. fuscum (табл. 1, 2). На девятый год восстановления величина живой фитомассы S. fuscum уже выше, чем у S. divinum, но от июньского пала 2018 г. первый вид пострадал значительно сильнее и среди мхов появился Aulacomnium palustre (Hedw) Schwāgr., правда в небольшом количестве. Сфагновые мхи здесь довольно медленно восстанавливаются в относительно сухие годы, темпы увеличиваются во влажном 2015 г. и снижаются после сухого 2016 г. Начиная с 2015 г. можно говорить об успешном восстановлении сфагнового покрова – долевое участие сфагнов приближается к показателям на ненарушенном участке в 2012 г. (табл. 4). В 2017 г. в снижении доли сфагнов возможно виновна усилившаяся конкуренция политриховых.
В сфагновом покрове на ненарушенном участке также происходили изменения. Если доля ПП S. divinum в сфагновом покрове в первые 3 года исследований колебалась в пределах 10%, с 2012 г. она начинает увеличиваться. Живая фитомасса S. divinum, учитывавшаяся при отборе монолитов, тоже увеличивается и к концу наблюдений уже практически равна фитомассе S. fuscum (табл. 2). Это вызвано скорее всего активной экспансией P. strictum, который предпочитает фусковые дернины. Началась она с 2013 г., на пятый год после пожара. Возможно, с 2017 г. сукцессия политриховых здесь пошла бы на спад, но ее явно стимулировал июньский пал 2018 года.
Данные, приведенные в таблице 4, позволяют проанализировать динамику долевого участия в живой надземной фитомассе фитоценоза сфагновых мхов и P. strictum. На ненарушенном участке доля P. strictum постоянно возрастает, темпы снижаются во влажные годы: 2015, 2017. Участие сфагновых снижается в 2013 г., в этом году годовая сумма осадков не дотягивает до нормы, а 2012 г. был сухой (рисунок).
Фитомасса P. strictum увеличивается в самые сухие годы (2012, 2014, 2016), исключение составляет 2017 г. Самая низкая фитомасса у P. strictum во влажные 2013, 2015 гг., при высоких показателях фитомассы сосудистых. Можно сказать, что кустарнички, как доминанты кустарничково-травяного яруса, являются конкурентами политриховым мхам, сдерживающими их развитие. В то время как со сфагновыми мхами у них взаимоотношения обратные.
Политриховые мхи – эндогидрильные виды, получают влагу по внутренней системе листьев и других тканей, в отличие от эктогидрильных сфагновых мхов, всасывающих воду всей поверхностью благодаря процессу диффузии (Proktor, 1982; Schofield, 1985). При длительном отсутствии осадков, что весьма характерно для первой половины лета в Приамурье, эктогидрильные мхи теряют в 2–3 раза больше влаги из листьев, чем эндогидрильные (Shpak, 2008). Но проводящая система политриховых еще очень примитивна и они, как и сфагновые, предпочитают компенсировать потери воды в плотных дерновинах, благодаря чему создается дополнительная капиллярная система для поднятия воды (Igumnova, Shamurin, 1965). Кроме того, скорость испарения у плотной подушки Polytrichum в 25 раз меньше, чем у отдельного растения (Stocker, 1956b). И наши наблюдения показывают, что P. strictum образует значительно большую фитомассу в фусковых дернинах, чем на торфе (табл. 2), где для него менее комфортные условия увлажнения.
По результатам геоботанических исследований в лесных сообществах взаимоотношений сфагновых и политриховых в двухвидовых синузиях, первые в конце концов вытесняют политриховые в ходе формирования конечной фазы смен сосняков долгомошных сосняками сфагновыми (Zhuravleva, Ipatov, 2003). На открытых верховых болотах, как считает С.Ю. Попов, олиготрофная (допожарная) гидросерия временно сменяется мезоолиготрофной гидросерией (“пирогенный динамический ряд”), затем восстанавливается допожарная растительность (Popov, 2000). Автор проводил исследования через 20 лет после пожара и на их основе, ссылаясь на Н.Я. Кац (Kats, 1934), делает вывод, что “олиготрофный ряд представляет собой циклическую сукцессию, способную существовать веками”. Некоторые исследователи тоже считают, что “Регрессивная эволюция болотных экосистем, спровоцированная торфяными пожарами, является обратимым процессом” (Efremova, Efremov, 2006). В Приамурье, с его специфическим крайне неравномерным распределением атмосферных осадков и частой повторяемостью пирогенного фактора эта закономерность может нарушаться, приводя к необратимым изменениям природы. У нас имеются факты наблюдений полной деградации кустарничково-сфагновых болот и трансформации их в безмоховые ерники под влиянием пирогенного фактора в Нанайском р-не Хабаровского края. Такие процессы наблюдаются и на юге Западной Сибири (Naumov et al, 2009).
ЗАКЛЮЧЕНИЕ
Таким образом, на первом этапе после катастрофического пожара развивается сукцессия Marchantia polymorpha и мелких бриевых мхов: Ceratodon purpureus и Cynodontium strumiferum. Длительность существования маршанциевой стадии 2 года, мелких бриевых – не более 5 лет. Одновременно, на второй год после пожара начинает развиваться сукцессия политриховых. На выгоревших участках болота их распространение и рост происходит очень быстро, мозаично, отдельными пятнами. В течение 4 лет они способны образовывать дерновину мощностью 30 см с генеративными органами (коробочками), но чаще всего полного развития не происходит. Однако повторные палы слабой и особенно средней интесивности, а также засушливые периоды пролонгируют течение сукцессии политриховых.
Восстановление сфагновых мхов на выгоревших участках происходит со сменой олиготрофных видов на мезоолиготрофные и мезотрофные. Не выгоревшие во время пожара участки болота с сохранившимся сфагновым покровом через некоторое время по мере развития сукцессии политриховых и увеличения банка их спор подвергаются активной экспансии P. strictum. В сфагновом покрове на этих участках также происходит временная смена видов олиготрофной гидросерии на мезоолиготрофную и мезотрофную. Повторные палы средней интенсивности существенно затормаживают восстановление сфагнового покрова.
Подводя итоги десятилетним наблюдениям за восстановлением мохового яруса, можно с уверенностью сказать, что полностью сфагновый покров восстановится только тогда, когда мощность сфагновой дернины на подушках и коврах будет не менее 30 см. С учетом того, что в условиях Приамурья S. divinum прирастает за год в среднем на 3 см, а S. fuscum на 2 см, потребуется еще не менее 10 лет при благоприятных для сфагновых мхов условиях увлажнения.
Список литературы
[Burenina] Буренина Т.А. 2006. Послепожарная динамика в лиственничных лесах Северо-Восточной Азии. – В сб.: Материалы Междунар. конф. “Лесные экосистемы Северо-Восточной Азии и их динамика”. Дальнаука. Владивосток. С. 152–156.
[Efremova, Efremov] Ефремова Т.Т., Ефремов С.П. 2006. Пирогенная трансформация органического вещества почв лесных болот. – Почвоведение. 12: 1441–1450.
[Elina et al.] Елина Г.А., Кузнецов О.Л., Максимов А.И. 1984. Структурно-функциональная организация и динамика болотных экосистем Карелии. Л. 127 с.
[Goncharova] Гончарова И.А. 2008. Особенности структуры дерновины и продуктивности мхов в лесоболотных комплексах Западной Сибири. – Сибирский экологический журнал. 15 (4): 607–616.
[Gorshkov et al.] Горшков В.В., Баккал И.Ю., Ставрова Н.И. 1995. Восстановление нижних ярусов сосновых лесов после пожаров. – Бот. журн. 80 (5): 35–46.
Hassel K., Kyrkjeeide M.O., Yousefi N., Prestø T., Stenøien H.K., Show J.A., Flatberg K.I. 2018. Sphagnum divinum (sp. nov.) and S. medium Limpr. and their relationship to S. magellanicum Brid. – Journal of Bryology. 40 (3): 197–222.
[Ignatov, Ignatova] Игнатов М.С., Игнатова Е.А. 2003. Флора мхов средней части Европейской России. Т. 1. М. 608 с.
[Igumnova, Shamurin] Игумнова З.С., Шамурин В.Ф. 1965. Водный режим лишайников и мхов в тундровых сообществах. – Бот. журн. 50 (5): 702–709.
[Ivanova et al.] Иванова Г.А., Иванов В.А., Ковалева Н.М., Конард С.Г., Жила С.В., Тарасов П.А. 2017. Сукцессия растительности после высокоинтенсивного пожара в сосняке лишайниковом. – Сибирский экологический журнал. 24 (1): 61–71.
[Kats] Кац Н.Я. 1934. К изучению строения и методике учета торфоподстилочного слоя. – Торфяное дело. (1): 35–40.
[Kharuk et al.] Харук В.И., Двинская М.Л., Рэнсон К.Дж. 2005. Пространственно-временная динамика пожаров в лиственничных лесах северной тайги Средней Сибири. – Экология. 5: 334–343.
[Kopoteva, Kuptsova] Копотева Т.А., Купцова В.А. 2016 а. Влияние пожаров на функционирование фитоценозов торфяных болот. – Экология. 1: 14–21.
[Kopoteva, Kuptsova] Копотева Т.А., Купцова В.А., 2016 б. Влияние пожаров на динамику фитомассы и первичной продукции мезотрофного кустарничково-сфагнового болота в Приамурье. – Журнал общей биологии. 77 (5): 397–405.
[Kopoteva] Копотева Т.А. 1999. Пирогенный фактор и динамика растительного покрова болот Приамурья. – В сб.: Материалы Междунар. конф.: “Болота и заболоченные леса в свете задач устойчивого природопользования”. М. С. 58–61.
[Kopoteva] Копотева Т.А. 2008. Трансформация растительности на мезотрофных болотах Приамурья. – В сб.: Материалы Всеросс. конф. “Фундаментальные и прикладные проблемы ботаники в начале ХХI века” Ч. 5. Петрозаводск. С. 145–147.
[Kopoteva] Копотева Т.А. 2011. Роль пожаров в формировании торфяной залежи и динамика надземных фитомассы и продукции в постпирогенный период на мезотрофных кустарничково-сфагновых болотах Среднеамурской низменности. – В сб.: Материалы Междунар. конф. “Резервуары и потоки углерода”. Сыктывкар. С. 81–86.
[Korchagin] Корчагин А.А. 1960. Определение возраста и длительности жизни мхов и печеночников. – Полевая геоботаника. Л. С. 279–314.
[Kosykh] Косых Н.П. Пирогенная сукцессия на болотах лесостепной зоны. – В сб.: Материалы III междунар. полевого симпозиума “Западно-Сибирские торфяники и цикл углерода, прошлое и настоящее”. Новосибирск. С. 183–185.
[Kovaleva et al.] Ковалева Н.М., Иванова Г.А., Кукавская Е.А. 2011. Восстановление напочвенного покрова после низовых пожаров в среднетаежных сосняках. – Лесоведение. 5: 30–35.
[Kovaleva, Ivanova] Ковалева Н.М., Иванова Г.А. 2013. Восстановление живого напочвенного покрова на начальной стадии пирогенной сукцессии. – Сибирский экологический журнал. 20 (2): 203–213.
[Kovaleva] Ковалева. И.М. 2015. Динамика фитомассы живого напочвенного покрова сосняков после низовых пожаров. – Растительные ресурсы. 51 (2): 145–153.
[Malysheva] Малышева Т.В. 1970. К методике разграничения живых и отмерших частей у мхов при учете их фитомассы. – Бот. журн. 55 (5): 704–709.
[Naumov et al.] Наумов А.В., Косых Н.П., Паршина Е.К., Артымук С.Ю. 2009. Верховые болота лесостепной зоны, их состояние и мониторинг. – Сибирский экологический журнал. 16 (2): 251–259.
O’Neill K.P. 2002. Role of bryophyte-dominated ecosystems in the global carbon budget. – Bryophyte Boilogy. Cambridge. P. 344–368.
[Perevosnikova et al.] Перевозникова В.Д., Иванова Г.А., Иванов В.А., Ковалева Н.М. 2007. Трансформация живого напочвенного покрова под воздействием пожаров в сосняках Средней Сибири. – Экология. 6: 476–480.
[Popov] Попов С.Ю. 2000. Пирогенные сукцессии сфагновых мхов в Средней России. – Бот. журн. 85 (2): 89–96.
Proctor M.C.F. 1982. Physiological Ecology: Water relation, Light and temperature responses, carbon balance. – Bryophyte Ecology. P. 333–381.
Schofield W.B. 1985. Introduction to bryology. Department of Botany. University of British Columbia. 23. Physiology. P. 309–329.
[Shpak] Шпак О.В. 2008. Эколого-физиологическая характеристика некоторых видов мхов в Хибинах: Автореф. дис. канд. биол. наук. Кировск. 98 с.
[Sofronov et al.] Софронов М.А., Волокитина А.В., Софронова Т.М. 2008. Пожары и пирогенные сукцессии в лесах Южного Прибайкалья. – Сибирский экологический журнал. 15 (3): 381–388.
[Sokolova et al.] Соколова Г.В., Глаголев В.А., Коган Р.М. 2009. Разработка автоматизированного метода оценки и прогноза пожарной опасности территории по погодным и метеорологическим условиям. – Лесное хозяйство. 4: 43–45.
Stocker O. 1956. Die Transpiration der Tallophyten. Handb. d. Pflanzenphysiologie, III.
[Titlyanova et al.] Титлянова А.А., Афанасьев А.Н., Наумова Н.Б. и др. 1993. Сукцессии и биологический круговорот. Новосибирск. 157 с.
[Tsaregradskaja, Kositsin] Цареградская С.Ю., Косицын В.Н. 1999. Оценка состояния растительного покрова после сильного торфяного пожара. – В сб.: Материалы Междунар. конф. “Болота и заболоченные леса в свете задач устойчивого природопользования”. М. С. 156–158.
[Volokitina et al.] Волокитина А.В., Софронова Т.М., Белянкин А.А. 2013. Восстановление напочвенного покрова после лесных низовых пожаров в Южном Прибайкалье. – Лесной журнал. 2: 58–66.
[Vomperskii et al.] Вомперский С.Э., Вомперская М.И., Валяева Н.А., Зазнобин М.Ю. 1999. Короткопериодные колебания оторфованности заболоченных лесов. – В сб.: Материалы Междунар. конф. “Болота и заболоченные леса в свете задач устойчивого природопользования”. М. С. 42–45.
[Zhuravleva, Ipatov] Журавлева Е.Н., Ипатов B.С. 2003. Взаимоотношения видов рода Sphagnum (Sphagnaceae) и Polytrichum commune (Polytrichaceae) в заболоченных сосновых лесах. – Бот. журн. 88 (8): 20–27.
Дополнительные материалы отсутствуют.
Инструменты
Ботанический журнал