

Ботанический журнал, 2019, T. 104, № 7, стр. 1090-1109
ИЗМЕНЧИВОСТЬ МОРФОЛОГИЧЕСКИХ ПРИЗНАКОВ ВИДОВ DELPHINIUM (RANUNCULACEAE) ЮГО-ВОСТОКА ЕВРОПЕЙСКОЙ ЧАСТИ РОССИИ
А. С. Кашин 1, *, А. В. Богослов 1, И. В. Шилова 1, Т. А. Крицкая 1, А. С. Пархоменко 1, Л. В. Гребенюк 1, Н. А. Петрова 1
1 Саратовский государственный университет им. Н.Г. Чернышевского
410012 Саратов, ул. Астраханская, 83, Россия
* E-mail: kashinas2@yandex.ru
Поступила в редакцию 01.03.2019
После доработки 26.06.2019
Принята к публикации 02.07.2019
Аннотация
На основе дисперсионного, кластерного и факторного анализов 61 качественных и количественных морфологических признаков образцов 16 популяционных выборок из пяти таксонов Delphinium L. Нижнего Поволжья и прилегающих территорий выделены группы, на которые подразделяются образцы, и основные признаки, по которым таксоны дифференцируются. Все указанные методы анализа достоверно подразделяют исследованные образцы на две группы. Одна группа включает все образцы D. puniceum Pall. и D. sergii Wissjul., вторая – все образцы трех прочих таксонов: D. duhmbergii Huth, D. litwinowii Sambuk и D. pubiflorum (DC.) Turcz. ex Huth. Внутри каждой из этих двух групп выделение таксонов видового ранга не столь отчетливо. Наименьшее число различий имеют образцы D. litwinowii и D. pubiflorum. Разделение образцов таксонов D. puniceum и D. sergii одних и тех же популяций в существенной степени зависит от погодных условий конкретного года наблюдений. При сравнении распределения облаков рассеяния точек выборочных совокупностей в пространстве главных компонент и географического распределения мест произрастания популяций исследованных таксонов показано, что распределение облаков рассеяния точек в факторном пространстве соответствует градиенту континентальности климата на исследованной территории. Сделан вывод о том, что выявленная морфологическая вариабельность среди исследованных таксонов рода связана, скорее всего, с градиентом показателей регионального климата юго-востока европейской части России.
Дельфиниум (Delphinium L.) – род преимущественно многолетних (реже однолетних) травянистых растений семейства лютиковые (Ranunculaceae), насчитывающий около 300 видов, распространенных в умеренно теплых и субтропических областях Северного полушария, а отчасти в горных районах тропиков (Tzvelev, 2001).
Отдельные таксоны рода, такие как D. elatum L., D. dictyocarpum DC. или D. cuneatum Steven ex DC., имеют весьма обширные ареалы, как естественные, так и культурные. Это служит причиной высокой морфологической изменчивости растений у них в пределах ареалов (Malyutin, 1973, 1984). В таком случае целая группа близкородственных таксонов с не всегда четкой экологической и географической дифференциацией, образующих, как полагают, на стыке ареалов переходные формы, рядом исследователей принимаются за один вид (Tutin et al., 1993; Fedorov, 2003, Dіdukh et al., 2004; Davydov, 2014). Например, Н.И. Фёдоров (Fedorov, 2003) характеризует такие группы как сингамеоны в понимании В. Гранта (Grant, 1957, 1981) или Р.В. Камелина (Kamelin, 2009). Однако другие авторы (Tzvelev, 1996, 2001; Maevsky, 2014), напротив, считают таксоны подобных комплексов самостоятельными видами, объединяя в группы, называя последние “агрегатами”.
Для Восточной Европы Н.Н. Цвелёв (Tzvelev, 1996, 2001) указывает 19 видов. Из них 15 он относит к секции Delphinastrum DC. и четыре вида – к секции Diedropetala Huth. В секции Delphinastrum им выделены четыре группы: 1. D. aggr. elatum L. с пятью видами (D. elatum s. str., D. alpinum Waldst. et Kit., D. cryophilum Nevski, D. nacladense Zapal., D. villosum Stev. ex DC.), 2. D. aggr. cuneatum Stev. ex DC. с четырьмя видами (D. cuneatum s. str., D. subcuneatum Tzvel., D. litwinowii Sambuk, D. duhmbergii Huth), 3. D. aggr. dictyocarpum DC. с четырьмя видами (D. pubiflorum (DC.) Turcz. ex Huth, D. dictyocarpum s. str., D. cyananthum Nevski, D. uralense Nevski), 4. D. aggr. cheilantum Fisch. с двумя видами (D. middendorffii Trautv., D. grandiflorum L.). В секции Diedropetala Huth он объединяет в группу D. aggr. fissum Waldst. et Kit. три вида (D. leuocarpum Huth, D. pallasii Nevski, D. sergii Wissjul.), оставляя самостоятельным, хорошо от них отличимый D. puniceum Pall.
При таком понимании объема видов в Среднем и Нижнем Поволжье (Ульяновская, Пензенская, Самарская, Саратовская, Волгоградская, Астраханская области, Республика Калмыкия) и прилегающих регионах (Республика Мордовия, Тамбовская, Воронежская, Ростовская, Оренбургская области) встречается 10 видов из рода Delphinium. Из секции Delphinastrum полностью представлена группа D. aggr. cuneatum: D. cuneatum s. str., D. subcuneatum, D. litwinowii, D. duhmbergii; более половины видов из группы D. aggr. dictyocarpum: D. pubiflorum, D. dictyocarpum, D. uralense; один вид из группы D. aggr. elatum: D. elatum s. str. Из секции Diedropetala встречаются два вида – D. sergii из группы D. aggr. fissum и D. puniceum (Tzvelev, 2001).
Однако при работе с гербарными сборами из Саратовской обл. мы столкнулись с целым рядом сложностей и разночтений в интерпретации их видовой принадлежности (Shilova et al., 2016). По результатам анализа было установлено, что D. pubiflorum распространен по территории Саратовской обл. гораздо шире, чем считалось ранее, причем многие из сборов определялись ранее как D. elatum, D. dictyocarpum, D. cuneatum или D. duhmbergii. Единичные образцы D. dictyocarpum и D. elatum имеют сомнительное происхождение и, вероятно, по этим соображениям А.Г. Еленевский с соавт. в своих работах (Elenevsky et al., 2008, 2009) проигнорировали их. Не меньше противоречий и нестыковок обнаружено и при определении гербарных сборов из соседних с Саратовской обл. регионов (Волгоградской, Воронежской, Пензенской, Самарской, Тамбовской, Ульяновской областей и Республики Мордовия) и примерно с тем же характером разночтений и многочисленных ревизий (Shilova et al., 2019). Все это указывает на то, насколько запутанной оказывается ситуация с определением видовой принадлежности и встречаемости тех или иных таксонов во флоре Саратовской области и прилегающих территорий из-за отсутствия четких морфологических критериев отнесения образцов к тому или иному таксону, присутствия отдельных таксономически значимых признаков разных таксонов у одного образца и неоднозначности представлений о таксономической структуре рода, по крайней мере, в отдельных ее составляющих.
В этой связи целью нашего исследования было изучение морфологической изменчивости популяций пяти таксонов рода Delphinium, произрастающих в пределах Ульяновской, Саратовской, Волгоградской, Астраханской, Воронежской, Пензенской и Тамбовской областей (рис. 1). В задачи исследования входило: выявление диапазона изменчивости морфологических (прежде всего, считающихся в отечественной флористике видоспецифичными) признаков у растений в популяциях таксонов; уточнение взаимоотношений между таксонами рода в пределах вышеуказанной территории; определение характера влияния погодно-климатических и экотопических факторов на совокупность морфологических признаков, характеризующих тот или иной таксон.
МАТЕРИАЛ И МЕТОДИКА
Исследования проведены в 2017–2018 гг. в пределах вышеуказанных регионов в 16 популяциях пяти таксонов рода, выделяемых в отечественной флористике в ранге вида (D. litwinowii, D. duhmbergii, D. pubiflorum, D. sergii, D. puniceum) (Tzvelev, 1996, 2001; Maevskiy, 2014) (рис. 1, табл. 1). Ограниченное число популяций, задействованных в исследовании, связано с редкой их встречаемостью во флорах указанных регионов.
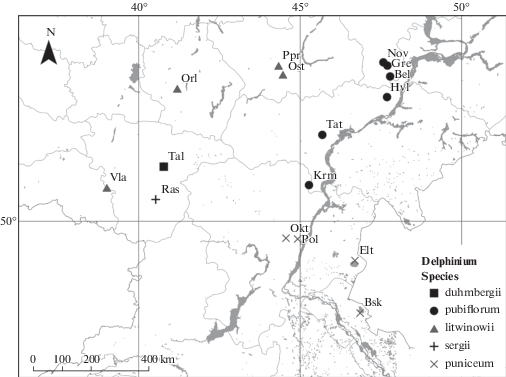
Таблица 1.
Перечень популяционных выборок исследованных таксонов Delphinium L., включенных в морфологический анализ Table 1. List of the Delphinium L. populations included in morphological analysis
| Условное обозначение популяции Population abbreviation |
Таксон Taxon |
Место произрастания Locality |
|---|---|---|
| Bsk | D. puniceum | Астраханская обл., Ахтубинский р-н, ЮВ склон г. Б. Богдо Astrakhan Region, Akhtubinsky district, southeastern slope of Bol’shoy Bogdo Mt. |
| Elt | D. puniceum | Волгоградская обл., Палласовский р-н, окр. оз. Эльтон Volgograd Region, Pallasovka district, Elton Lake environs |
| Pol | D. puniceum | Волгоградская обл., Дубовский р-н, окр. х. Полунино Volgograd Region, Dubovka district, farm Polunino environs |
| Okt | D. puniceum | Волгоградская обл., Ольховский р-н, окр. пос. Октябрьский Volgograd Region, Olkhovka district, Oktyabrsky village environs |
| Krm | D. pubiflorum | Саратовская обл., Красноармейский р-н, окр. с. Каменка Saratov Region, Krasnoarmeysk district, Kamenka village environs |
| Tat | D. pubiflorum | Саратовская обл., Татищевский р-н, окр. д. Ильиновка Saratov Region, Tatishchevo district, Ilyinovka village environs |
| Hvl | D. pubiflorum | Саратовская обл., Хвалынский р-н, окр., с. Акатная Маза Saratov Region, Khvalynsk district, Akatnaya Maza village environs |
| Bel | D. pubiflorum | Ульяновская обл., Радищевский р-н, окр. д. Белогоровка Ulyanovsk Region, Radishchevo district, Belogorovka village environs |
| Gre | D. pubiflorum | Ульяновская обл., Радищевский р-н, окр. п. Гремячий Ulyanovsk Region, Radishchevo district, Gremyachiy village environs |
| Nov | D. pubiflorum | Ульяновская обл., Новоспасский р-н, окр. с. Новая Лава Ulyanovsk Region, Novospasskoye district, Novaya Lava village environs |
| Tal | D. duhmbergii | Воронежская область, Таловский р-н, окр. ст. Таловая Voronezh Region, Talovaya district, Talovaya village environs |
| Ras | D. sergii | Воронежская обл., Павловский р-н, окр. х. Рассоховатое Voronezh Region, Pavlovsk district, Rassokhovatoye village environs |
| Vla | D. litwinowii | Воронежская обл., Острогожский р-н, окр. д. Владимировка Voronezh Region, Ostrogozhsk district, Vladimirovka village environs |
| Orl | D.litwinowii | Тамбовская обл., Тамбовский р-н, окр. с. Орловское Tambov Region, Tambov district, Orlovskoye village environs |
| Ost | D. litwinowii | Пензенская обл., Колышлейский р-н, окр. х. Островцы Penza Region, Kolyshley district, Ostrovtsy farm environs |
| Ppr | D. litwinowii | Пензенская обл., Пензенский р-н, окр. с. Поперечное Penza Region, Penza district, Poperechnoye village environs |
Качественные и количественные морфологические признаки оценивались только у растений средневозрастного онтогенетического состояния. В анализе по каждой популяционной выборке участвовало от (3) 10 до 30 образцов (малое число образцов связано с малочисленностью некоторых популяций и малочисленностью особей средневозрастного онтогенетического состояния в них). Суммарный объем всех популяционных выборок составил 530 образцов.
В качестве рабочей гипотезы виды понимались в объеме, предлагаемом Цвелёвым (Tzvelev, 2001).
В анализе использованы признаки вегетативных и генеративных органов, а также их возможные состояния. Всего для каждого образца каждой популяции был измерен и описан 61 признак: 29 количественных (табл. 2) и 32 качественных. Качественным признакам присваивался балл, согласно их состоянию. Кроме принятых в систематике рода таксономически значимых качественных признаков (Tzvelev, 2001; Maevskiy, 2014) дополнительно анализировались такие признаки, как тип ветвления соцветия, форма чашелистиков, форма верхушки чашелистиков, форма шпорца у цветка из середины соцветия, характер поверхности шпорца, форма отгиба лепестков-нектарников, форма верхушки лепестков-нектарников, соотношение длины лепестков-нектарников и чашелистиков, характер опушения черешка и шпорца.
Таблица 2.
Основные количественные признаки, дифференцирующие таксоны рода Delphinium. Приведены лимиты (в круглых скобках), 95%‑ный доверительный интервал среднего значения и величина F-критерия Table 2. Basic quantitative parameters used to differentiate between Delphinium taxa. The limits (in parentheses), 95% confidence interval of average value, and F-criterion value are given
| № признака Parameter number |
Признак Parameter | Таксон/Taxon | F, при p ≤ 0.05 F, at p ≤ 0.05 |
||||
|---|---|---|---|---|---|---|---|
| D. pubiflorum | D. duhmbergii | D. litwinowii | D. sergii | D. puniceum | |||
| 1 | Высота растения, см Plant height, cm | (40.0)–109.4– 116.4–(200.0) |
(23.0)–157.2– 214.6–(425.0) |
(85.0)–139.1– 148.5–(200.0) |
(43.0)–89.5– 102.0–(140.0) |
(25.0)–45.4– 49.3–(73.0) |
241.0372 |
| 2 | Диаметр куста, см Diameter of bush, cm |
(15.0)–66.3– 75.0–(183.0) |
(15.0)–73.6– 146.8–(450.0) |
(25.0)–70.6– 85.3–(200.0) |
(6.0)–19.3– 24.8–(63.0) |
(3.0)–19.6– 22.2–(40.0) |
74.4237 |
| 3 | Число генеративных побегов, шт. Number of generative shoots, pcs. |
(2.0)–4.9– 6.2–(27.0) |
(2.0)–5.6– 10.3–(24.0) |
(2.0)–4.1– 6.1–(44.0) |
(1.0)–1.0– 1.1–(2.0) |
(1.0)–1.5– 2.0–(8.0) |
30.3141 |
| 4 | Длина генеративного побега, см Generative shoot length, cm |
(45.0)–121.3– 129.4–(210.0) |
(23.3)–166.3– 211.0–(300.0) |
(85.0)–142.3– 152.0–(200.0) |
(43)–96.6– 111.4–(160.0) |
(25.0)–45.4– 49.3–(73.0) |
261.4764 |
| 5 | Число листьев на генеративном побеге, шт. Number of leaves on generative shoot, pcs. |
(13.0)–26.9– 28.7–(52.0) |
(14.0)–29.1– 36.0–(54.0) |
(16.0)–31.0– 33.9–(53.0) |
(1.0)–3.33– 4.5–(9.0) |
(4.0)–5.4– 5.7–(8.0) |
499.3876 |
| 6 | Длина междоузлия, см Internode length, cm |
(0.5)–4.3– 4.8–(10.0) |
(2.0)–4.7– 6.3–(12.0) |
(1.0)–4.0– 4.8–(20.0) |
(7.0)–14.0– 16.8–(28.0) |
(1.0)–5.0– 5.9–(12.0) |
209.3425 |
| 7 | Диаметр стебля, мм Stem diameter, mm |
(2.0)–5.7– 6.2–(11.6) |
(2.4)–6.7– 9.8–(23.6) |
(4.2)–9.1– 10.0–(16.9) |
(1.9)–5.0– 5.8–(9.9) |
(1.9)–3.9– 4.4–(7.8) |
118.8373 |
| 8 | Число цветков в соцветии, шт. Number of flowers in inflorescence, pcs. |
(6.0)–46.5– 55.2–(188.0) |
(5.0)–72.4– 112.8–(194.0) |
(16.0)–68.4– 81.3–(189.0) |
(5)–64.4– 92.4–(283.0) |
(6.0)–32.4– 39.5–(113.0) |
33.3480 |
| 9 | Длина соцветия, см Inflorescence length, cm |
(4.5)–23.4– 26.34–(60.0) |
(7.0)–36.5– 52.8–(93.0) |
(9.0)–32.7– 38.9–(105.0) |
(7.0)–46.1– 57.3–(115.0) |
(10.0)–22.5– 25.8–(54.0) |
55.7927 |
| 10 | Число паракладиев, шт. Number of paracladia, pcs. |
(0)–5.2– 6.1–(18.0) |
(1.0)–5.6– 8.5–(15.0) |
(1.0)–5.9– 6.8–(16.00) |
(0)–1.9– 2.5–(6.0) |
(0)–1.0– 1.4–(4.0) |
81.9217 |
| 11 | Длина наиболее развитого паракладия, см Length of the most developed parakladium, cm |
(1.5)–7.3– 8.2–(20.1) |
(5.0)–14.5– 18.7–(26.0) |
(4.0)–14.0– 16.3–(35.0) |
(7.0)–21.4– 30.3–(80.0) |
(2.5)–10.5– 13.2–(35.0) |
75.4321 |
| 12 | Длина чашелистика, мм Sepal length, mm |
(5.81)–10.8– 11.4–(19.7) |
(5.2)–8.0– 9.4–(12.8) |
(5.6)–9.3– 10.0–(15.6) |
(5.5)–7.2– 8.0–(12.4) |
(5.3)–7.3– 7.7–(11.1) |
88.6051 |
| 13 | Ширина чашелистика, мм Sepal width, mm |
(3.6)–7.1– 7.5–(12.8) |
(3.6)–5.7– 6.9–(9.9) |
(3.7)–6.6– 7.1–(11.5) |
(2.5)–3.7– 4.1–(6.0) |
(2.5)–4.0– 4.3–(6.0) |
156.3523 |
| 14 | Длина шпорца, мм Spur length, mm |
(6.2)–12.4– 13.0–(19.5) |
(2.4)–11.4– 13.3–(16.8) |
(6.9)–12.6– 13.5–(22.0) |
(8.3)–10.4– 11.0–(13.0) |
(5.0)– 9.5– 10.0–(13.0) |
48.8500 |
| 15 | Ширина шпорца, мм Spur width, mm |
(1.6)–3.1– 3.3–(8.8) |
(1.71)–3.3– 4.6–(7.02) |
(1.21)–3.0– 3.3–(9.74) |
(1.1)–2.2– 2.5–(3.9) |
(1.4)–2.5– 2.7–(4.2) |
25.3046 |
| 16 | Длина листовой пластинки, мм Length of leaf blade, mm |
(51.6)–88.1– 92.7–(144.7) |
(47.1)–83.9– 99.5–(128.0) |
(57.1)–89.5– 95.3–(135.2) |
(23.6)–57.8– 68.8–(104.2) |
(12.9)–41.4– 46.7–(73.3) |
160.5050 |
| 17 | Длина нерасчлененной части листовой пластинки, мм Length of unsegmented part of leaf blade, mm |
(0)–14.6– 16.4–(42.5) |
(6.3)–13.5– 17.7–(32.9) |
(7.8)–19.4– 21.4–(37.7) |
(0)–0.1– 0.8–(8.75) |
(0)–0– 0.2–(3.9) |
274.2276 |
| 18 | Длина центрального сегмента средней лопасти, мм Length of central segment of middle lobe, mm |
(15.23)–43.5– 46.3–(78.5) |
(26.0)–39.4– 48.7–(67.3) |
(26.2)–43.8– 47.5–(77.5) |
(17.2)–36.3– 43.6–(65.3) |
(11.8)–31.1– 35.4–(61.8) |
22.9057 |
| 19 | Ширина листовой пластинки, мм Width of leaf blade, mm |
(70.4) –126.7– 132.9–(184.0) |
(89.0)–126.6– 147.5–(192.0) |
(71.2)–124.9– 133.7–(196.0) |
(30.6)–94.7– 116.0–(196.0) |
(19.3)–73.0– 82.0–(139.1) |
86.9524 |
| 20 | Ширина основания центрального сегмента средней лопасти, мм Base width of central segment of middle lobe, mm |
(2.6)–7.6– 8.2–(15.4) |
(5.6)–9.0– 10.5–(14.1) |
(6.5)–11.2– 12.2–(20.3) |
(0.86)–2.5– 3.5–(6.6) |
(0.9)–2.2– 2.6–(5.7) |
296.9968 |
| 21 | Максимальная ширина средней лопасти, мм Maximum width of middle lobe, mm |
(16.2)–42.5– 46.0–(79.35) |
(22.6)–40.4– 48.1–(67.1) |
(18.1)–40.2– 43.9–(71.2) |
(7.7)–34.8– 45.4–(86.2) |
(6.3)–30.6– 36.0–(65.9) |
12.1432 |
| 22 | Ширина основания средней лопасти, мм Base width of middle lobe, mm |
(2.0)–6.9– 7.6–(15.9) |
(4.1)–8.2– 10.3–(15.7) |
(4.8)–10.6– 11.6–(17.9) |
(0.6)–1.0– 1.3–(3.0) |
(0.5)–1.4– 1.6–(3.4) |
323.5889 |
| 23 | Длина черешка, мм Petiole length, mm |
(14.6)–53.2– 58.1–(119.4) |
(46.7)–79.6– 101.7–(162.5) |
(13.3)–69.7– 78.6–(164.0) |
(10.0)–45.4– 60.1–(157.4) |
(10.0)–53.2– 63.2–(137.3) |
25.6060 |
| 24 | Ширина черешка, мм Petiole width, mm |
(0.8)–1.5– 1.6–(2.6) |
(1.2)–1.6– 1.8–(2.3) |
(1.2)–1.8– 2.0–(2.9) |
(1.1)–2.2– 2.5–(4.1) |
(0.7)–1.6– 1.8–(3.1) |
43.1780 |
| 25 | Длина прицветника, мм Bract length, mm |
(4.9)–12.4– 14.1–(35.7) |
(5.2)–8.4– 10.0–(14.5) |
(5.6)–9.2– 10.1–(18.4) |
(2.9)–5.3– 6.7–(12.3) |
(2.7)–5.3– 5.8–(8.6) |
75.5001 |
| 26 | Ширина прицветника, мм Bract width, mm |
(0.2)–0.6– 0.7–(1.6) |
(0.9)–1.3– 1.6–(2.6) |
(1.1)–2.3– 2.5–(3.8) |
(0.2)–0.3– 0.4–(0.6) |
(0.2)–0.4– 0.5–(1.0) |
604.3150 |
| 27 | Длина прицветничка, мм Bracteole length, mm |
(3.5)–6.4– 6.8–(11.5) |
(4.5)–5.8– 6.7–(8.4) |
(3.7)– 6.3– 6.8–(10.8) |
(1.5)–3.1– 3.5–(4.8) |
(1.7)–3.8– 4.1–(6.4) |
138.2966 |
| 28 | Ширина прицветничка, мм Bracteole width, mm |
(0.2)–0.5– 0.6–(1.3) |
(0.6)–1.1– 1.3–(1.66) |
(1.2)–2.1– 2.3–(3.8) |
(0.2)–0.3– 0.4–(0.5) |
(0.2)–0.3– 0.4–(0.6) |
825.7933 |
| 29 | Длина цветоножки, мм Pedicle length, mm |
(4.8)–13.6– 14.7–(30.4) |
(3.0)–10.1– 13.2–(19.2) |
(8.2)–15.9– 17.6–(36.2) |
(5.4)–10.2– 12.2–(19.3) |
(5.2)–9.9– 10.8–(16.6) |
44.5257 |
Примечания. * – ноль вместо минимального значения означает отсутствие морфологической единицы у некоторых растений в выборке. Полужирным шрифтом выделены значения признаков, дифференцирующих таксон, и значения Fф > Fст. (4.8).
Note. * – zero instead of a minimum value indicates the absence of morphological feature in some specimens. Bold typeface marks the taxon-differentiating parameter values and Ff > Fst values (4.8).
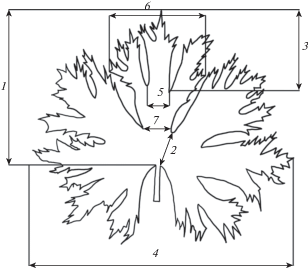
При измерении параметров листа следовали рекомендациям Д.М. Анатова и соавт. (Anatov et al., 2015) (рис. 2).
Рис. 2.
Учтенные морфологические признаки листьев видов Delphinium (по: Anatov et al., 2015): 1 – длина листовой пластинки; 2 – длина нерасчлененной части листовой пластинки; 3 – длина центрального сегмента средней лопасти; 4 – ширина листовой пластинки; 5 – ширина основания центрального сегмента средней лопасти; 6 – максимальная ширина средней лопасти; 7 – ширина основания средней лопасти.
Fig. 2. Recorded morphological traits of leaves of the Delphinium species (after: Anatov et al., 2015): 1 – the length of the leaf blade; 2 – the length of the unsegmented part of the leaf blade; 3 – the length of the central segment of the middle lobe; 4 – the width of the leaf blade; 5 – the base width of the central segment of the middle lobe; 6 – the maximum width of the middle lobe; 7 – the base width of the middle lobe.
Индивидуальная изменчивость растений в популяциях Delphinium по 23 качественным признакам предварительно оценивалась визуально с помощью метода пиктограмм StarPlot (Chambers et al., 1983; Markov, 2012).
Статистический анализ внутрипопуляционной и внутривидовой изменчивости морфологических признаков проводили с использованием дисперсионного, кластерного и факторного анализов в программах PAST 3.17 (Hammer et al., 2001) и STATISTICA 13 For Windows (STATISTICA, 2018).
При построении пиктограмм и при кластерном анализе взято 224 образца (по 10 случайных растений из каждой популяции, отобранных с помощью генератора случайных чисел, кроме белогоровской (Bel, по 4 и 3 образца в 2017 и 2018 гг., соответственно) и владимировской (Vla, 6 образцов).
Изменчивость количественных признаков оценивали по их лимитам и 95%-ному доверительному интервалу среднего арифметического. Достоверность различия выборок оценивали с помощью однофакторного дисперсионного анализа (табл. 2).
Кластерный анализ проведен методом невзвешенного попарного среднего (UPGMA) по всем промеренным и описанным качественным и количественным признакам. Средняя таксономическая дистанция рассчитывалась с использованием дистанции Говера, т.к. в анализе используются качественные признаки. Количество перестановок в бутстреп-тесте составило 1000.
Факторный анализ выполнен методом главных компонент (PCA) с использованием матрицы корреляций. Анализ проведен по 427 образцам, у которых отсутствовали пропущенные данные в исходной матрице. Всего в анализе участвовало 178 образцов D. pubiflorum, 26 образцов D. duhmbergii, 109 образцов D. litwinowii, 32 образца D. sergii, 82 образца D. puniceum.
С целью снижения эффектов разного масштаба измерений для разных признаков перед проведением кластерного и факторного анализа данные стандартизировались.
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
В предварительной оценке индивидуальной изменчивости растений по 23 качественным признакам с использованием метода StarPlot (не показано) выявлено, что пиктограммы разных популяций одного вида, полученные в один год наблюдений, более сходны между собой, чем пиктограммы одной и той же популяции, полученные в разные годы наблюдений. Это отражает особенности роста и развития растений в популяции, связанные с межсезонными различиями погодных условий. В то же время растения, произрастающие в популяции в один и тот же сезон, в случае, когда они давали более или менее заметно различающиеся пиктограммы, отражали именно фенотипические внутрипопуляционные различия между растениями.
Полученные пиктограммы позволили визуально по качественным признакам предварительно разделить выборку на три группы. В одну вошли образцы из популяций D. puniceum и D. sergii, во вторую – D. duhmbergii. Третья группа объединила менее отчетливо различающиеся между собой D. pubiflorum и D. litwinowii.
Растения первой группы – D. puniceum и D. sergii – отличались от прочих бархатисто-войлочным опушением стеблей; эллиптическими чашелистиками; как правило, гладкой поверхностью шпорца; нектарниками, как правило, равными по длине чашелистикам (однако данное выражение признака зачастую наблюдалось также у D. duhmbergii и в одной из популяций D. litwinowii); волосистыми листьями (в отдельных популяциях D. puniceum – до бархатисто-войлочных, а в популяции D. sergii часть растений имела листья, опушенные лишь по краям и жилкам, как и в популяциях прочих видов). В первой группе растения D. puniceum отличались от D. sergii и других видов пурпурным цветом чашелистиков, всегда до половины надрезанной верхушкой лепестков-нектарников, бархатисто-войлочным опушением черешка; более интенсивно, чем у других видов, опушенными прицветничками; расположением прицветничков выше середины цветоножки.
Растения второй группы – D. duhmbergii – отличались от остальных видов голой осью соцветия; голыми или опушенными малочисленными ресничками прицветничками; голыми или опушенными редкими волосками цветоножками и чашелистиками снаружи; голыми завязями и плодиками. С D. litwinowii эту группу растений объединяет черно-бурый цвет нектарников и ланцетная или ланцетно-яйцевидная форма прицветничков.
Растения третьей группы – D. pubiflorum и D. litwinowii – по большей части параметров были сходны между собой. Различались они цветом лепестков-нектарников, как указано выше; у растений D. pubiflorum, в отличие от D. litwinowii, были густо опушены завязи и плодики (в этом растения D. pubiflorum схожи с D. puniceum и D. sergii), а также чашелистики не только с наружной, но и с внутренней стороны, у D. litwinowii завязи и плодики по швам были реснитчатые или голые, чашелистики были опушены лишь снаружи.
Результаты анализа изменчивости количественных признаков, оцененной по их лимитам и 95%-ному доверительному интервалу среднего арифметического, представлены в табл. 2. С помощью однофакторного дисперсионного анализа показано, что при разделении исходной выборки на пять групп, соответствующих рассматриваемым таксонам, вычисленное значение F-критерия для всех рассматриваемых признаков при p ≤ 0.05 (Fст = 4.8) указывает на достоверное различие между группами. Из данных табл. 2 видно, что лимиты и доверительные интервалы подавляющего большинства из рассматриваемых признаков у всех исследуемых таксонов перекрываются. Единственным признаком, абсолютно достоверно отличающим два таксона из секции Diedropetala от таксонов из секции Delphinastrum, является число листьев на генеративном побеге. Кроме того, в пределах доверительных интервалов, D. sergii и D. puniceum имеют минимальный диаметр куста, самые короткие и узкие чашелистики и прицветники и самые узкие прицветнички. D. sergii отличается в пределах доверительных интервалов от всех прочих видов самыми длинными междоузлиями, соцветиями (по этому признаку частично перекрываясь с D. duhmbergii) и паракладиями; широкими черешками (частично перекрываясь по лимитам с остальными видами); самыми малочисленными побегами (частично перекрываясь с D. puniceum), малочисленными листьями, узкими шпорцами, узким основанием средней лопасти, наиболее узкими прицветниками (соприкасаясь по верхнему пределу с нижним пределом D. puniceum). D. puniceum отличается (в пределах доверительных интервалов) от остальных видов низкорослостью, короткими генеративными побегами, тонкими стеблями, короткими соцветиями с малочисленными паракладиями и малочисленными цветками, короткими шпорцами, короткими и узкими листовыми пластинками, в том числе – лопастями и их сегментами. D. duhmbergii выделяется (в пределах доверительных интервалов) по максимальной величине следующих признаков: высота и диаметр куста, длина генеративного побега, ширина шпорца (соприкасаясь нижним пределом с верхним пределом данного признака у D. pubiflorum и D. litwinowii), длина черешка. D. pubiflorum отличается (в пределах доверительных интервалов) от прочих исследованных видов короткими паракладиями, длинными и широкими чашелистиками (частично перекрываясь по последнему признаку с D. litwinowii), узкими черешками, длинными прицветниками. Из всех исследованных видов D. litwinowii выделяется (в пределах доверительных интервалов) наибольшей длиной нерасчлененной части листовой пластинки, наиболее широким основанием центрального сегмента средней лопасти и самой средней лопасти, наиболее широкими прицветниками и прицветничками, длинными цветоножками.
В кластерном анализе, проведенном по 29 количественным и 32 качественным морфологическим признакам (рис. 3) образцов пяти исследуемых таксонов Delphinium из 16 популяционных выборок методом невзвешенного попарного среднего (UPGMA), кофенетическая корреляция матрицы расстояний и полученного дерева была достигнута на уровне 0.83, что указывает на хорошее совпадение результатов с матрицей данных (Rohlf, 1993).
Рис. 3.
Фенограмма изученных в 2017–2018 гг. популяций видов рода Delphinium, полученная при обработке матрицы 61 морфологического признака (табл. 2) методом невзвешенного попарного среднего (UPGMA). Способ объединения – дистанция Говера.
Fig. 3. Phenogram of Delphinium populations studied in 2017–2018, obtained from UPGMA treatment of the matrix of 61 morphological parameters (Table 2). The Gower distance is used as a clustering method.
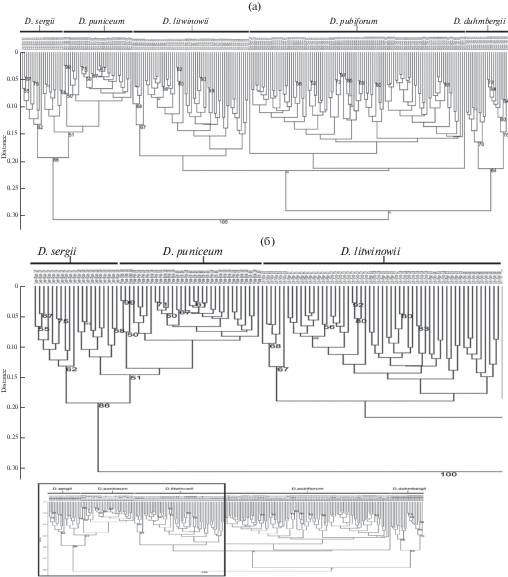
Рис. 3.
Окончание.
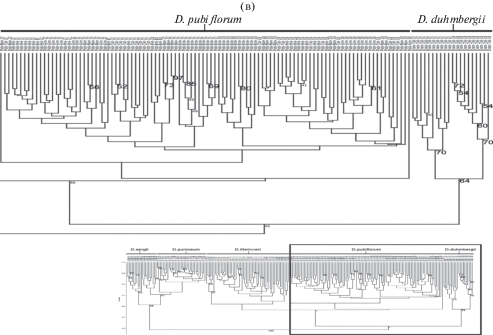
При этом анализ выявил две основные группы с уровнем связи 0.31. Кластер I включает образцы всех четырех популяционных выборок D. puniceum и обеих выборок D. sergii. С высокой поддержкой (индекс бутстрепа 86%) этот кластер распадается на два подкластера. При этом в первый подкластер попала группа образцов D. sergii, исследованных в 2017 г. Во второй подкластер попали все образцы D. puniceum и группа образцов D. sergii, исследованных в 2018 г. Популяция D. sergii в 2017 г. была многочисленной, с высокой жизненностью. В 2018 г., по-видимому, в связи с поздней и холодной весной, большая часть особей в данной популяции находилась в покое, а вегетировавшие имели более низкую жизненность и были более близкими с D. puniceum, чем с особями своей же популяции предыдущего года наблюдения. Следует отметить, что во втором подкластере межпопуляционная и внутрипопуляционная изменчивость была несколько выше, чем межвидовая.
Второй кластер с невысокой поддержкой (индекс бутстрепа 43%) распадается на два подкластера. Один объединяет все образцы D. duhmbergii. Второй подкластер тоже с невысокой поддержкой (индекс бутстрепа 46%), в свою очередь, распадается на два подкластера II порядка. Один из них включает все образцы D. pubiflorum, второй – все образцы D. litwinowii. При этом наблюдается зачастую более высокая межпопуляционная и внутрипопуляционная изменчивость по сравнению с межвидовой и образцы второго кластера фактически не разделяются этим методом анализа.
Таким образом, кластерный анализ по совокупности количественных и качественных признаков позволил подразделить образцы на кластеры, включающие образцы D. puniceum и D. sergii, – с одной стороны, – и D. duhmbergii, D. pubiflorum и D. litwinowii, – с другой. Таксоны второго кластера этим методом не разделяются, а разделение образцов таксонов D. puniceum и D. sergii одних и тех же популяций в существенной степени зависит от погодных условий конкретного года наблюдений.
Результаты анализа методом главных компонент изученных в 2017–2018 гг. образцов по 29 количественным и 32 качественным морфологическим признакам позволили выявить пять групп, каждая из которых включает образцы одного из исследованных таксонов (рис. 4). При этом первая главная компонента объясняет 30.05% вариаций в матрице данных, вторая – 10.84%.
Рис. 4.
Диаграмма рассеяния всех изученных представителей рода Delphinium методом главных компонент (PCA) по 61 морфологическому признаку.
Fig. 4. Scatterplot of all the studied representatives of Delphinium by the principal component method (PCA) for 61 morphological characters.
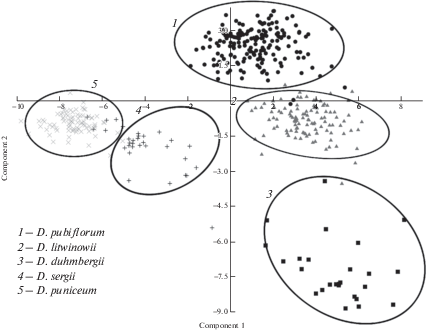
Максимальные по модулю значения факторной нагрузки (0.70–0.99) по первой главной компоненте имеют следующие параметры: высота растения, длина генеративного побега, число листьев на побеге, число паракладиев, длина листовой пластинки, длина нерасчлененной части листа, ширина листовой пластинки, ширина основания центральной доли листа, ширина основания средней лопасти, форма основания листовой пластинки, характер опушения листа, степень рассечения листа, опушение черешка, расположение прицветничков. По второй главной компоненте максимальные факторные нагрузки (0.60–0.99) имеют параметры: опушение чашелистиков снаружи и внутри и опушение шпорца.
Таким образом, первая компонента в большей степени отражает изменения по: а) размеру растения, б) числу, форме и размеру листьев, включая прицветники и прицветнички, в) степени рассечения листовой пластинки, г) степени разветвленности соцветий, д) опушению всей вегетативной и частично генеративной сферы, т.е., прежде всего, она отражает изменения размерности и опушенности вегетативных частей растения. Вторая компонента отражает изменения по опушенности цветка.
В первую группу входит подавляющее большинство образцов D. pubiflorum, во вторую – подавляющее большинство образцов D. litwinowii, в третью – подавляющее большинство образцов D. duhmbergii. При этом облако рассеяния точек, соответствующих образцам D. pubiflorum частично перекрывается с облаком рассеяния точек, соответствующих образцам D. litwinowii, которое, в свою очередь, в меньшей мере, но все же перекрывается с облаком рассеяния точек, соответствующих D. duhmbergii. Четвертая группа объединяет подавляющее большинство образцов D. sergii, а пятая – объединяет подавляющее большинство образцов D. puniceum. Облака рассеяния точек, соответствующих образцам этих двух таксонов, также перекрываются. То есть, по результатам факторного анализа нет полного разделения между облаками рассеяния точек, соответствующих образцам D. duhmbergii, D. litwinowii и D. pubiflorum, а облака рассеяния точек, соответствующих образцам D. sergii и D. puniceum, перекрываются в еще большей степени.
Таким образом, и кластерный, и факторный анализы по совокупности количественных и качественных признаков разделяют исследованные популяционные выборки только на группы, включающие образцы D. puniceum и D. sergii, – с одной стороны, – и D. duhmbergii, D. pubiflorum и D. litwinowii, – с другой. Таксоны внутри каждой из этих групп до конца не разделяются, что указывает на недостаточную правомочность выделения их в качестве самостоятельных видов.
Обращает на себя внимание тот факт, что D. pubiflorum, D. litwinowii и D. duhmbergii, образовали облака рассеяния точек, разнесенные в пространстве главных компонент, прежде всего, по второй, – вертикальной, – компоненте, отражающей изменения по опушению цветка, т.е. основных таксономически значимых признаков, отличающих эти таксоны. В то же время D. sergii и D. puniceum образовали облака рассеяния точек, разнесенные в пространстве главных компонент, прежде всего, по первой, горизонтальной, – компоненте, отражающей изменения размерности и опушенности вегетативных частей растения, в меньшей степени вносящие вклад в таксономически значимые отличия между этими таксонами.
При сравнении распределения облаков точек выборочных совокупностей в пространстве главных компонент (рис. 4) и географического распределения мест произрастания популяций исследованных таксонов (рис. 1) обращает на себя внимание сходный характер трендов. А именно, тренд распределения облаков рассеяния точек, соответствующих образцам отдельных таксонов, фактически идентичен тренду географического распределения мест произрастания популяций этих таксонов. При этом образцы популяций D. puniceum и D. sergii, произрастающие на юго-востоке и юго-западе исследованной территории, соответственно, образовали облака рассеивания точек в левой части пространства главных компонент, причем образцы юго-восточных популяций (D. puniceum) образовали самое левое по расположению в пространстве главных компонент облако рассеивания. Образцы популяций D. pubiflorum, произрастающих на востоке и северо-востоке исследованной территории, образовали облако рассеяния точек в центральной верхней части пространства главных компонент, а образцы популяций D. duhmbergii и D. litwinowii, произрастающих на северо-западе исследованной территории, образовали облака рассеивания точек в правой нижней части пространства главных компонент. Очевидно, что пространственное географическое расположение мест произрастания популяций исследованных таксонов и расположение облаков рассеяния точек, соответствующих образцам этих таксонов, совпадает с градиентом континентальности климата на исследованной территории (Pryachina et al., 2006; Rulev, 2012; Pryachina, Ormeli, 2017; Kireeva-Genenko et al., 2017). Таким образом, выявленная морфологическая вариабельность среди исследованных таксонов рода связана, скорее всего, с градиентом показателей регионального климата юго-востока европейской части России.
Та же самая закономерность в целом прослеживается и при сравнении распределения точек, соответствующих образцам различных популяций D. pubiflorum, в пространстве главных компонент (рис. 5а, б, в) и географического распределения этих популяций с юга на север по Приволжской возвышенности в пределах Саратовской и Ульяновской областей (рис. 1) и дифференцировано по каждому из лет исследования (2017 и 2018 гг.). По результатам наблюдения 2017 г. (рис. 5а) в левой верхней части пространства главных компонент образовала облако рассеяния точек самая южная по географическому положению места произрастания популяция из Красноармейского р-на Саратовской обл. В правой верхней и центральной части пространства главных компонент оказалось облако рассеяния точек, соответствующих образцам популяции, произрастающей в центре исследованной части Приволжской возвышенности – из Татищевского р-на Саратовской обл. В центральной части пространства главных компонент образовали рыхлое облако рассеяния образцы самой северной из саратовских популяций – из Хвалынского р-на, а в нижней части пространства главных компонент – образцы популяций из Ульяновской обл. (Bel, Nov, Gre). Между последними популяциями эта закономерность менее отчетлива в силу их близкого пространственного расположения относительно друг друга (в пределах 15–30 км, в то время как между местами произрастания популяций из Красноармейского и Хвалынского р-нов Саратовской обл. расстояние превышает 300 км, а между популяцией из Хвалынского р-на и популяциями из Ульяновской обл. – порядка 40–70 км. По результатам наблюдения 2018 г. (рис. 5б) облако рассеяния точек, соответствующих образцам самой южной из саратовских популяций (Krm), распределилось в верхней, – преимущественно левой, – части пространства главных компонент. Облако рассеяния точек, соответствующих центральной саратовской популяции (Tat), расположилось в центре левой половины пространства главных компонент, северной саратовской популяции (Hvl) – в центре правой половины пространства главных компонент, а образцы ульяновских популяций (Bel, Nov, Gre) образовали облако рассеяния точек в нижней половине пространства главных компонент.
Рис. 5.
Диаграмма рассеяния образцов различных популяций D. pubiflorum в пространстве главных компонент: а – в 2017 г., б – в 2018 г., в – в 2017 и 2018 гг. Условные обозначения популяций соответствуют указанным в табл. 1.
Fig. 5. Scatterplot of the samples of different D. pubiflorum populations in the space of principal components: a – in 2017, б – in 2018, в – in 2017 and 2018. For the population abbreviatons see Table 1.
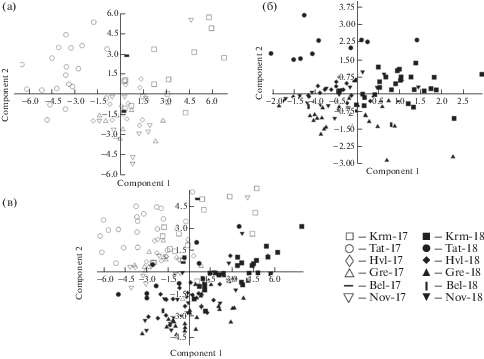
При объединенном анализе популяционных выборок D. pubiflorum за два года наблюдения (2017–2018 гг.) в пространстве главных компонент (рис. 5в) показано, что все популяции таксона, исследованные в 2018 г., образовали облака рассеяния точек правее и ниже облаков рассеяния точек, соответствующих образцам тех же популяций, исследованных в 2017 г. Это дополнительно указывает на роль погодных условий в дифференциации их по совокупному набору анализируемых признаков.
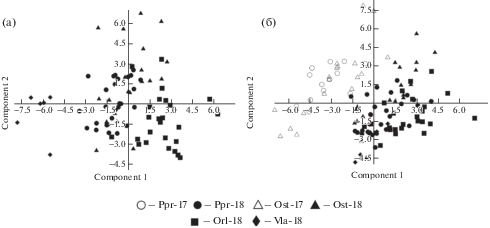
Аналогичная закономерность в целом прослеживается и при сравнении распределения точек, соответствующих образцам различных популяций D. litwinowii, в пространстве главных компонент и их географического распределения в северо-западной части исследованной территории (в пределах Воронежской, Тамбовской и Пензенской обл.) (рис. 1), что на примере наблюдений за 2018 г. показано на рис. 6a. Популяции Ppr и Ost, произрастающие северо-восточнее остальных популяций этого таксона, образовали рыхлые не разделяющиеся облака в центральной части пространства главных компонент, северо-западная (Orl) – в правой, а юго-западная (Vla) – в левой части пространства главных компонент. При этом, так же как в случае распределения облаков рассеяния точек, соответствующих образцам D. pubiflorum, образцы D. litwinowii при объединенном анализе популяционных выборок за два года наблюдения в факторном пространстве все популяции таксона, исследованные в 2018 г., образовали облака рассеяния точек правее и ниже облаков рассеяния точек, соответствующих образцам тех же популяций, исследованных в 2017 г. (рис. 6б).
Рис. 6.
Диаграмма рассеяния образцов различных популяций D. litwinowii в пространстве главных компонент: а – в 2018 г., б – в 2017 и 2018 гг. Условные обозначения популяций соответствуют указанным в табл. 1.
Fig. 6. Scatterplot of the samples of different D. litwinowii populations in the space of principal components: a – in 2018, б – in 2017 and 2018. For the population abbreviatons see Table 1.
На роль экологических условий произрастания в морфологической изменчивости исследованных таксонов косвенно указывает и то, что морфологически сходные между собой растения D. pubiflorum и D. litwinowii произрастают в сходных экотонных (разреженные заросли кустарников) или открытых остепненных сообществах; растения D. duhmbergii произрастают в густых кустарниковых зарослях; D. sergii – в открытых степных сообществах, а D. puniceum – в наиболее засушливых местообитаниях.
Межвидовую и межпопуляционную изменчивость в различной степени зависимости от условий произрастания растений таксонов рода Delphinium отмечали ранее для других частей их ареалов (Malyutin, 1973, 1984; Fedorov, 2003).
ЗАКЛЮЧЕНИЕ
По результатам проведенного анализа следует признать, что таксономическая структура рода Delphinium в пределах юго-востока европейской части России не вполне отчетлива. Хорошо обособлены морфологически только секции Diedropetala и Delphinastrum. Для более уверенного выделения видов необходимо привлекать кроме применяемых в отечественной систематике (форма основания листовой пластинки, степень рассечения листа, характер опушения листовой пластинки, расположение прицветничков, опушение чашелистиков снаружи и внутри) и другие качественные (опушение шпорца, опушение черешка), а также некоторые количественные (высота растения, длина побега, число листьев на побеге, число паракладиев, длина листовой пластинки, длина нерасчлененной части листовой пластинки, ширина листовой пластинки, ширина основания центральной доли, ширина основания средней лопасти) признаки.
Приведенный выше анализ по совокупности количественных и качественных признаков позволил подразделить образцы на кластеры, включающие D. puniceum и D. sergii, – с одной стороны, – и D. duhmbergii, D. pubiflorum и D. litwinowii, – с другой. Таксоны второго кластера использованными методами не разделяются, а разделение образцов таксонов D. puniceum и D. sergii одних и тех же популяций в существенной степени зависит от погодных условий конкретного года наблюдений.
Для установления окончательного статуса таксонов рода Delphinium на исследованной части их ареалов требуется дополнительное уточнение с использованием секвенирования пластидной и ядерной ДНК большого количества образцов из большего числа популяционных выборок. В этой связи следует отметить, что в большинстве случаев заключения о морфологической разделенности или неразделенности принятых в современной отечественной флористике таксонов носит описательный умозрительный характер. Представление о том, что переходные формы в агрегатах по границам ареалов имеют гибридное происхождение, является также чисто умозрительным заключением, основанным на встречаемости у одних и тех же образцов признаков разных таксонов или промежуточном выражении ряда признаков, считающихся таксономически значимыми. Однако гибридность этих переходных форм не доказана генетически.
Выявленная в ходе проведенного анализа морфологическая вариабельность как между, так и внутри исследованных таксонов рода связана, скорее всего, с градиентом показателей регионального климата юго-востока европейской части России.
Список литературы
[Anatov et al.] Анатов Д.М., Амирова Л.А., Раджабов Г.К. 2015. Внутривидовая изменчивость природных популяций Delphinium crispulum Rupr. по морфологическим признакам и суммарному содержанию флавоноидов и антоцианов в условиях Дагестана. – Ботанический вестник Северного Кавказа. 1: 8–17.
Chambers Jo.M., Cleveland W.S., Kleiner B., Tukey P.A. 1983. Graphical Methods for Data Analysis. California: Wadsworth. 395 p.
[Davydov] Давидов Д.А. 2014. Номенклатурні проблеми, що стосуються деяких видів рослин, занесених до “Червоної книги України”. – У зб.: Рослинний світ у Червоній книзі України: провадження Глобальної стратегії збереження рослин. Матеріали III Міжнародної наукової конференції. Львів. С. 27–28.
[Dіdukh et al.] Дідух Я.П., Зиман С.М., Бурда Р.І., Четверних І.С. 2004. Delphinium cuneatum Stev. ex DC. – Дельфіній клиноподібний. – У кн.: Екофлора України. Київ. 2: 115–116.
[Elenevskii et al.] Еленевский А.Г., Буланый Ю.И., Радыгина В.И. 2008. Конспект флоры Саратовской области. Саратов. 232 с.
[Elenevskii et al.] Еленевский А.Г., Буланый Ю.И., Радыгина В.И. 2009. Определитель сосудистых растений Саратовской области. Саратов. 248 с.
[Fedorov] Фёдоров Н.И. 2003. Род Delphinium L. на Южном Урале: экология, популяционная структура и биохимические особенности. Уфа. 149 с.
Grant V. 1957. The plant species in theory and practice. – In: The Species Problem, ed. E. Mayr, Amer. Assoc. Adv. Sci. Washington, D.C. P. 39–80.
Grant V. 1981. Plant speciation. New York. 563 p.
Hammer O., Harper D.A.T., Ryan P.D. 2001. PAST: Paleontological statistics software package for education and data analysis. – Palaeontol. Electron. 4 (1): 9.
[Kamelin] Камелин Р.В. 2009. Особенности видообразования у цветковых растений. – Труды ЗИН РАН. 313 (S1): 141–149.
[Kireeva-Genenko et al.] Киреева-Гененко И.А., Новикова Е.П., Чумейкина А.С. 2017. Анализ и оценка индекса континентальности климата в Центрально-Черноземном районе за последние 30 лет. – Успехи современного естествознания. 7: 76–80.
[Maevskii] Маевский П.Ф. 2014. Флора средней полосы европейской части России. 11 изд. М. 635 с.
[Malyutin] Малютин Н.И. 1973. Филогения и систематика рода Delphinium L. – Бот. журн. 58 (12): 1710–1722.
[Malyutin] Малютин Н.И. 1984. Дельфиниумы. М. 80 с.
[Malyutin] Малютин Н.И. 1987. Система рода Delphinium (Ranunculaceae), основанная на морфологических признаках семян. – Бот. журн. 72 (5): 683–693.
[Markov] Марков М.В. 2012. Популяционная биология растений. М. 387 с.
[Priachina et al.] Пряхина С.И., Фридман Ю.Н., Васильева М.Ю. 2006. Мониторинг климата Саратовской области. – Известия Саратовского ун-та. Новая серия. Серия Науки о Земле. 6 (1): 15–18.
[Priachina, Оrmeli] Пряхина С.И., Ормели Е.И. 2017. Расчет индексов континентальности для Среднего и Нижнего Поволжья. – Известия Саратовского ун-та. Новая серия. Серия Науки о Земле. 17 (1): 17–19.
Rohlf F.J. 1993. Relative warp analysis and an example of its application to mosquito. – Contributions to morphometrics. 8: 1–131.
[Rulev] Рулев А.С. 2012. Компьютерное картографирование пространственного распределения градиентов показателей регионального климата юго-востока Европейской части России. – Вестник ВолГУ. Серия 11. 1 (3): 72–77.
[Shilova et al.] Шилова И.В., Петрова Н.А., Ермолаева Н.Н., Кашин А.С., Архипова Е.А. 2016. О распространении видов рода Delphinium L. (Ranunculaceae) на территории Саратовской области. – Бот. журн. 101 (7): 842–849.
[Shilova et al.] Шилова И.В., Богослов. А.В., Крицкая Т.А., Кашин А.С. 2019. О трактовке видовой принадлежности гербарных сборов Delphinium (Ranunculaceae) Юго-Востока Европейской России. – Бюлл. бот. сада СГУ: 17 (2–3): 73–89.
STATISTICA (data analysis software system), version 13 / StatSoft, Inc., 2018. http://www.statsoft.com.
The Plant List: http://www.theplantlist.org/tpl1.1/search?q=Delphinium. Дата создания: 2012. Дата обращения: 05.02.2019.
Tutin T.G., Burges N.A., Chater A.O., Edmondson J.R., Heywood V.H., Moore D.M., Valentine D.H., Walters S.M., Webb D.A. 1993. Flora Europaea. Vol. 1: Psilotaceae to Platanaceae. Second Edition. New York. 629 p.
[Tzvelev] Цвелёв Н.Н. 1996. О некоторых родах семейства Лютиковых (Ranunculaceae) в Восточной Европе. – Бот. журн. 81 (12): 112–122.
[Tzvelev] Цвелёв Н.Н. 2001. Род 10. Живокость – Delphinium. – В кн: Флора Восточной Европы. Т. 10. СПб. С. 66–74.
Дополнительные материалы отсутствуют.
Инструменты
Ботанический журнал