

Ботанический журнал, 2019, T. 104, № 9, стр. 1446-1459
КСЕРОФИЛЬНЫЕ РЕЛИКТЫ ВО ФЛОРЕ СЕВЕРНОГО КАВКАЗА
М. А. Тайсумов 1, 2, *, Р. С. Магомадова 2, М. У. Умаров 1, М. А.-М. Астамирова 1, 2, Л. В. Багмет 3, **, А. С. Абдурзакова 2
1 Академия наук Чеченской Республики
364024 г. Грозный, пр-т М. Эсамбаева, 13, Россия
2 Чеченский государственный педагогический университет
364037 г. Грозный, ул. Киевская, 33, Россия
3 Федеральный исследовательский центр Всероссийский институт генетических ресурсов растений имени Н.И. Вавилова
190000 г. Санкт-Петербург, ул. Б. Морская, д. 42, Россия
* E-mail: musa_taisumov@mail.ru
** E-mail: herbar@yandex.ru
Поступила в редакцию 11.12.2018
После доработки 31.08.2019
Принята к публикации 10.09.2019
Аннотация
На основе данных о распространении эндемичных видов, входящих в состав моно и олиготипных таксонов, и установлении их флорогенетических связей, высказано предположение о гетерохронном проникновении на территорию Кавказа анцестральных ксерофильных элементов, мигрировавших из отдаленных центров образования ксерофильной флоры (среднеазиатского и средиземноморского) и вторичных, прилежащих к Кавказу – иранского и малоазиатского. Хронологически начало становления флоры ксерофитов Кавказа датируется верхним миоценом – периодом значительного горообразования, особенно возвышения Большого Кавказа.
ОБЪЕКТ И МЕТОДЫ ИССЛЕДОВАНИЯ
Ксерофиты – это часть биологического разнообразия, особая группа растений, сформировавшаяся в результате адаптивной эволюции под действием одного из важнейших экологических факторов внешней среды – дефицита влаги. Этот фактор вызвал целый веер адаптаций, следствием которых явилось появление большого разнообразия приспособительных анатомо-морфологических структур и физиологических особенностей среди систематически разных групп видов, которые позволили переносить неблагоприятные условия дефицита влаги во внешней среде от почти полного пересыхания и впадения в анабиоз до крайне короткого сокращения вегетационного периода в онтогенезе. Эти виды в настоящее время составляют около четверти флоры Российского Кавказа. Одной из нерешенных до конца проблем является история формирования флоры ксерофитов на этой территории.
Для решения этой проблемы в качестве объектов исследования взяты положение в системе таксонов и пространственная локализация эндемичных ксерофильных видов флоры Российского Кавказа, относящихся к категории систематических реликтов, а также установление их родственных связей.
Основные методы исследования – метод исторической реконструкции, метод морфолого-эколого-географического анализа, включая метод эволюционных рядов, а также метод флорогенетического анализа и синтеза (Каmеlin, 2004). Информация о генетических и географических связях обсуждаемых таксонов получена путем анализа положения вида в системе рода (в случае монотипных родов – положения в системе семейства), что позволило выявить ближайших родственников и определить их географическую локализацию, предположить время и направления миграционных потоков анцестральных видов. Подобная методика была использована при анализе флоры западной части Центрального Кавказа (Galushko, 1976), анализе флоры Предкавказья (Ivanov, 1998).
Анализ распространения эндемичных видов позволил различать автохтонные (аборигенные) элементы, видообразование которых происходило в определенных локусах северного макросклона Большого Кавказа, и аллохтонные (миграционные) элементы, анцестральные типы которых формировались вне пределов северного макросклона. Становление последних сочетало миграционные процессы, с одной стороны, и процессы вымирания и сокращения ареалов видов, с другой. По времени происхождения эндемики подразделяются на две группы:
– палеоэндемики, имеющие регрессивные (сокращающиеся), локальные или сильно дизъюнктивные ареалы, далеко отстоящие от распространения близкородственных видов, также, в свою очередь, обитающих на ограниченных территориях;
– неоэндемики, обладающие прогрессивными (расширяющимися) ареалами, во многих случаях относимые к географическим расам.
Эти аналитические данные позволяют проводить флорогенетические построения, в том числе и корректировать имеющиеся модели флорогенеза.
Для выяснения начальных этапов флорогенеза целесообразно рассмотреть филогенетические связи особой категории эндемиков, относящейся к систематическим реликтам, стоящим обособленно в системе родов определенного семейства и не имеющих ближайших родственников на видовом уровне. Сам факт наличия в составе флоры подобного типа эндемиков является следствием длительных эволюционных процессов ее формирования, связанных, в первую очередь, с древними миграциями анцестральных форм, вызванных изменениями климата и палеогеографии в пределах Древнего Средиземья и прилегающих территорий.
ОБСУЖДЕНИЕ
В ксерофильной флоре Кавказа насчитывается три монотипных эндемичных рода: Chiastophyllum (Ledeb.) Stapf ex Berger (Crassulaceae), Muehlbergella Feer (Campanulaceae), Pseudobetckea (Hueck) Lincz. (Valerianaceae), а также один, условно олиготипный род, Cladochaeta DC. (Asteraceae); представители монотипных секций (подсекций) – Psephellus annae Galushko (Asteraceae), Primula juliae Kusn. (Primulaceae), Campanula hypopolia Trautv. (Campanulaceae); виды, входящие в состав монотипных рядов (14 видов) – Pyrethrum galushkoi Prima (Asteraceae), Thymus pulchellus C.A. Mey. (Lamiaceae), Scabiosa gumbetica Boiss. (Dipsacaceae), Jurinea woronowii Iljin (Asteraceae), Psephellus boissieri (Sosn.) Sosn. (Asteraceae) и др.; представители олиготипных секций и рядов (11 видов) – Potentilla ghalgana Juz. (Rosaceae), Psephellus schistosus (Sosn.) Alieva (Asteraceae), Cephalaria charadzeae Schchian (Dipsacaceae), Campanula ossetica Bieb. (Campanulaceae), Salvia daghestanica Sosn. (Lamiaceae) и др. Все они являются ключевыми таксонами, родственные связи которых позволяют проследить ранние этапы флорогенеза ксерофильной флоры.
Chiastophyllum oppositifolium (Ledeb.) Berger (Crassulaceae) является эндемиком Западного Кавказа и Западного Закавказья и субэндемиком исследуемой территории. Большая часть ареала его расположена от верховий бассейна р. Мзымты до верховий бассейна р. Риони, а на северном макросклоне Кавказского хребта – от верховий бассейна р. Белой до верховий бассейна р. Уруп. Это суккулент, обитающий под пологом леса на известняковых скалах. Представитель монотипного кавказского рода, выделенный из обширного, преимущественно средиземноморского рода Umbilicus DC., от видов которого отличается следующими признаками: ползучее корневище (а не клубневидно утолщенная или округлая подземная часть); накрест супротивное листорасположение (а не очередное); плоские листья, к основанию вдруг суженные в черешок (а не щитовидные или сердцевидные, в середине ворончато-углубленные) (Borisova, 1939, 1969). Результаты молекулярно-филогенетического анализа, на основании которого выделяются клады – группы близкородственных родов, показали (Goncharova, 2012), что род Umbilicus s.l. (incl. Chyastophyllum (Ledeb.) Stapf ex Berger) занимает обособленное положение в системе семейства Crassulaceae, он монокладовый (клада Umbilicus включает только один род), наиболее близок к многородовым кладам Rhodiola и Hylotelephium. Следовательно, следует искать его родственные связи с анцестральным средиземноморским предком, отделившимся от Umbilicus DC., очевидно, в верхнем палеогене, и в виде условного Praechiastophyllum попавшим на Кавказ с первой волной миграции.
Cladochaeta candidissima (Bieb.) DC. (Asteraceae) – субэндемик, эндемик центральной и восточной частей Большого Кавказа. Основная часть ареала вида расположена на северном макросклоне Большого Кавказа от верховий Баксана в Кабардино-Балкарии до верховий Ахтычая в Дагестане, в среднегорьях известен в долинах рек Аргун и Сулак. На южном макросклоне распространен в бассейнах рек Б. Лиахви и Алазани, а также р. Куры. В Азербайджане его ареал доходит до западной оконечности Апшеронского полуострова. В состав рода включается еще и C. caspica Sosn. ex Grossh., однако ареал этого вида находится в переделах ареала C. candidissima (Bieb.) DC., и, по мнению А.Г. Борисовой (Borisova, 1959), является его экологической расой. Ю.Л. Меницкий (Menytzky, 2008) также считает этот род монотипным, единственный вид которого близок к южнозакавказским видам рода Helichrysum Mill. ряда Callyсhisa Kirp.
Muehlbergella oweriniana (Rupr.) Feer (Campanulaceae) – локальный эндемик Внутригорного Дагестана, распространенный в нижнем течении Аварского Койсу и на хребте Салатау. Габитуально сходен с видами рода Edraianthus A. DC., которые распространены в Средиземноморье и на Балканском полуострове. Ан.А. Федоров при обработке рода во “Флоре СССР” (Fedorov, 1957b) не признавал самостоятельности рода Muehlbergella Feer и сближал Edraianthus owerinianus (Rupr.) Feer с E. pumilio A. DC. – эндемиком хребта Биоково в Хорватской области Далмации. Однако А.А. Колаковский (Kolakovsky, 1995) на основе анализа карпологических признаков семейства Campanulaceae пришел к заключению, что Muehlbergella Feer и Edraianthus A.DC. не являются близкими родами, они резко отличаются карпологическими признаками (или строением плода). Плод видов рода Edraianthus A.DC. аксикорновый11, цветоложе-разрывной, коробочки сплошь окружены жесткими прицветниками, конические, тонкостенные, но с сетью жилок, не поддающихся быстрому разрушению. Аксикорны апикальные, сросшиеся основаниями, со свободными носиками, передвигающиеся по стенкам конической чашечки, после разрыва очень тонкой осевой колонки полностью разрывают тонкостенный диск. Плод же Muehlbergella Feer безаксикорновый, створчато-разламывающийся – тип очень мелкой коробочки, раскрывание которой происходит в результате раздвигания толстостенных створок до самого основания коробочки, и в период ее раскрывания опадают зубцы чашечки и венчик, а створки отгибаются наружу. По типу вскрывания плода Muehlbergella Feer ближе всего к родам Jasione L. и Wahlenbergia Schumah. По карпологическим и ряду других признаков, в том числе и по подушковидной жизненной форме, вид проявляет заметное сходство с ксерофильным кальцефилом Pseudocampanula dzaaku (Albov) Kolak. – эндемичным абхазским видом, также представителем монотипного кавказского рода, обитающим на известняковых скалах (Kolakowcky, 1995). Оба вида древние, палеогеновые.
Pseudobetckea caucasica (Boiss.) Lincz. (Valerianaceae) – субэндемик, эндемик восточной части Большого Кавказа, распространенный на северном макросклоне от верховий р. Шаро-Аргун до верховий Самура и его правых притоков, на южном макросклоне встречается на горном массиве Бабадаг в Азербайджане. И А. Линчевский, обрабатывавший этот род для “Флоры СССP” (Linchevsky, 1958), высказал предположение, что “этот вид происходит от какого-то из видов рода Valeriana L., и что он производит впечатление сравнительно молодого видообразования в кавказской флоре” (с. 682). Что касается первой части цитаты, то действительно, Pseudobetckea (Hueck) Lincz. по некоторым признакам имеет много общего с видами рода Valeriana L., а именно в строении гинецея и плода, который отличается только отсутствием перистого хохолка (и соответственно редукцией отгиба чашечки в цветке). Относительно возраста этого вида (молодости) это утверждение можно поставить под сомнение. О том, что этот вид древний, свидетельствует его систематическая обособленность. Экологически он принадлежит к рефугиоксерофитам22 с эфемерным типом онтогенеза во второй половине лета (июль-август). Такими же эфемерами являются представители рода Valerianella Mill. с весеннее-летним периодом онтогенеза (май-июнь), но от Pseudobetckea (Hueck) Lincz. они резко отличаются трехгнездной завязью и строением плода, на котором формируются различные по форме придатки, производные чашечки. Кроме того, он отличается и экологически от всех Valerianaceae, это наиболее высокогорный вид, обитающий на осыпях субнивального пояса. Следует предположить, что анцестральная форма Pseudobetckea (Hueck) Lincz. обособилась до первой миграционной волны ксерофитов, и ее становление происходило уже в условиях высокогорий Восточного Кавказа.
Такую же значимость, как и монотипные для флорогенетических построений имеют олиготипные роды, насчитывающие 2–3 вида (соответственно ди- и тритипные). По времени формирования они такие же, как и монотипные. Такая категория эндемиков относится к т.н. “схизоэндемикам”. Этот термин введен Favarger, Contandriopoulus (1961) и относится к типу географического видообразования, при котором эндемики формируются путем медленной и прогрессивной дифференциации анцестрального таксона в разных частях его ареала (Favarger, Contandriopoulus, 1961). Подобный способ видообразования отмечен в единственном роде среди изученных нами ксерофитов – Limoniopsis Lincz. Схизоэндемиками также следует считать представителей олиготипных секций и рядов.
Limoniopsis owerinii (Boiss.) Lincz. (Plumbaginaceae) – представитель олиготипного (дитипного) рода, локальный эндемик Центрального Дагестана, ареал которого находится в низовьях рек Андийское и Аварское Койсу. Второй представитель рода L. davisii Bochary распространен в Восточной Анатолии. В целом род занимает обособленное положение в семействе Plumbaginaceae, близок, с одной стороны, к центральноазиатскому олиготипному (тритипному) роду Chaetolimon (Bunge) Lincz. и монотипному среднеазиатскому роду Ikonnikovia Lincz., с другой (Linchevsky, 1952).
Ареалы четырех из пяти вышеприведенных родов расположены в высокогорьях и среднегорьях центральной и восточной частей Большого Кавказа с доминированием в восточной части (Muehlbergella oweriniana (Rupr.) Feer, Pseudobetckea caucasica (Boiss.) Lincz., Limoniopsis owerinii (Boiss.) Lincz). Ареал Cladochaeta candidissima (Bieb.) DC. занимает также и центральную часть (рис. 1). Chiastophyllum oppositifolium (Ledeb.) Berger распространен на Западном Кавказе и в Западном Закавказье, но участок его ареала есть и на Малом Кавказе, что может быть следствием более поздних миграционных процессов из первичного видообразовательного центра.
Рис. 1.
Ареалы монотипных родов флоры ксерофитов Северного Кавказа.
Fig. 1. Ranges of monotypic genera of the xerophytic flora of the Northern Caucasus.
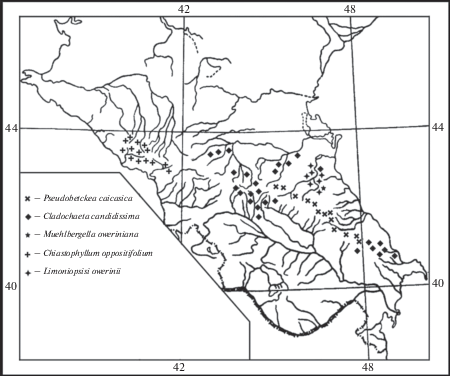
Исходя из вышесказанного, родственные связи монотипных и одного олиготипного рода проявляются как с восточной (Limoniopsis Lincz.), так и с западной частями Древнего Средиземья (Muehlbergella Feer, Chiastophyllum (Ledeb.) Stapf ex Berger, Cladochaeta DC.). Для рода Pseudobetckea (Hueck) Lincz. такие связи не прослеживаются, что заставляет предположить его гибридогенное происхождение. Анцестральные формы видов этих родов являются древнейшими мигрантами в область Большого Кавказа.
Не менее важное значение для теоретического обоснования флорогенетических построений имеют внутриродовые эндемичные таксоны – представители монотипных секций (подсекций) и рядов, которые наряду с монотипными родами являются древнейшими обособленными систематически представителями флоры. Они также проявляют локальный эндемизм, есть среди них эвриэндемики и субэндемики, значение которых для флорогенетических построений также значимо, как и статус монотипных родов. Таких видов три (рис. 2):
Рис. 2.
Ареалы представителей монотипных секций флоры ксерофитов Северного Кавказа.
Fig. 2. Ranges of representatives of monotypic sections of the xerophytic flora of the Northern Caucasus.
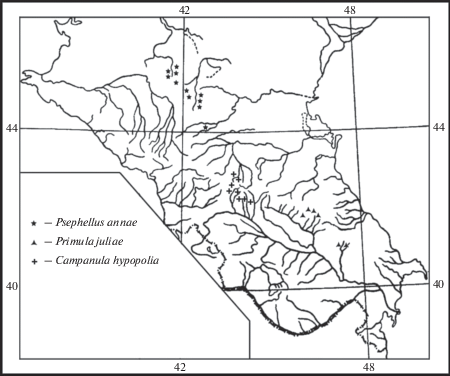
Psephellus annae Galushko (Asteraceae) – эндемик Ставропольской возвышенности (Ставропольские, Бешпагирские, Прикалаусские высоты) и лакколитов Кавказских минеральных вод (г. Машук) (монотипная секция Heterolophus). Близок к видам эукавказской секции Leucophylli Sosn. – P. leucophyllus (Bieb.) C.A. Mey. и P. declinatus (Bieb.) C. Koch (Galushko, 1979).
Primula juliae Kusn. (Primulaceae) – эфемероидный рефугиоксерофит, эндемик восточной части Большого Кавказа, на северном макросклоне распространенный в верховьях рек Джурмут и Самур, на южном – на сопредельной территории Азербайджана. Представитель монотипной кавказской секции Juliana Fed. et Losinsk. Проявляет родственные связи, с одной стороны, со среднеазиатско-сибирско-дальневосточным видом ряда Carthusiformes Fed. секции Cortusoides Balf. f., с другой – с кавказско-малоазиатским видом секции Carolinella (Hemsl.) Pax (Fedorov, 1952).
Campanula hypopolia Trautv. (Campanulaceae) – эндемик центральной части Большого Кавказа, известный из верховий р. Ардон на северном макросклоне и верховий притоков рек Риони и Б. Лиахви на южном. Представитель монотипной кавказской подсекции Hypopolion Fed. Проявляет родство со среднеазиатскими видами подсекции Rupestres (Boiss.) Fed. (ряд Saxicolae Boiss.) и европейско-сибирско-дальневосточными видами подсекции Heterophylla (Nym.) Fed. (ряд Vulgares Witasek) (Fedorov, 1957а).
Среди представителей монотипных секций обнаружены один узколокальный эндемик и два субэндемика. Ареалы их расположены в Центральном и Восточном Кавказе. Один вид – эндемик Предкавказья (Ставропольская возвышенность и г. Машук). Ближайшие родственники Psephellus annae Galushko – кавказские виды, два других вида проявляют связи с видами восточной части Древнего Средиземья. Можно предположить, что представители монотипных и дитипных родов и секций, как древнейшие типы, являются остатками первой миграционной волны анцестральных ксерофитов на Кавказ, родственные связи которых прослеживаются в большей степени со среднеазиатским древним центром образования ксерофитов и в меньшей степени со средиземноморским.
Виды монотипных рядов (в понимании таксономической концепции В.Л. Комарова, принятой авторами “Флоры СССР”), также являются древними типами, обособившимися в процессе эволюции, не имеющими близкого родства внутри рода. Таких видов 14 (рис. 3):
Рис. 3.
Ареалы представителей монотипных рядов флоры ксерофитов Северного Кавказа.
Fig. 3. Ranges of representatives of monotypic series of xerophytic flora of the Northern Caucasus.
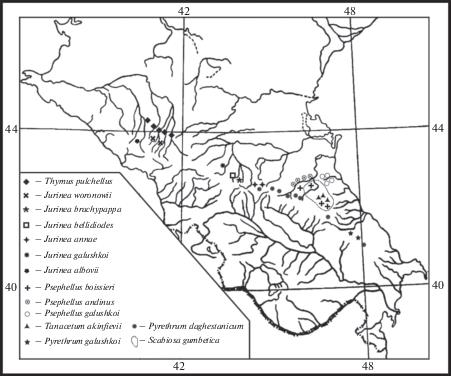
Pyrethrum galushkoi Prima (Asteraceae) – локальный эндемик гор Шалбуздаг (Южный Дагестан) и Тфандаг (Азербайджан); P. daghestanicum (Rupr. ex Boiss.) Fler. (P. aromaticum Tzvel.) – эвриэндемик, распространенный в восточной части северного макросклона, основная часть ареала которого простирается от верховий р. Армхи до верховий р. Дюльты-чай узкой полосой по Боковому хребту. Имеется указание на его нахождение на г. Бабадаг в Азербайджане (Prima, 1974). Входят в состав дитипного ряда Daghestanica Tzvel. секции Pyrethrellum Tzvel. По мнению Н.С. Ханджян (Khanjyan, 2008), этот ряд можно считать монотипным, представленным P. daghestanicum, остальные виды (P. galushkoi и P. aromaticum) – его синонимы. Родственные связи прослеживаются с представителями дитипного закавказского ряда Daucifolia Tzvel. (P. chamaemelifolium (Somm. et Levier) Sosn. и P. silaifolium Stev.), а также с монотипнорядным эндемиком восточной части Северного Кавказа P. tricholobum Sosn. eх Manden. (Tsvelev, 1961а).
Thymus pulchellus C.A. Mey. (Lamiaceae) – эвриэндемик, распространенный на Скалистом хребте от ущелья р. Кизинчи (бассейн Б. Лабы) до ущелья р. Кубань. Относится к монотипному ряду Pulchelli Klok. et Schost. подсекции Goniotrichi Borb. Родственные связи прослеживаются с карпатским эндемиком Th. enervius Klok., также стоящим в системе обособленно (Klokov, 1954).
Scabiosa gumbetica Boiss. (Dipsacaceae) – эндемик Внутригорного Дагестана, ареал которого ограничен бассейнами низовий рек Койсу. Монотипный ряд Gumbeticae Bobr. систематически обособлен и близок, с одной стороны, к западнозакавказскому эндемику S. olgae Albov (также монорядному), с другой, – к представителям дитипного ряда Argenteae Bobr. – закавказско-малоазиатскому S. argentea L. и понтическому S. ucranica L. (Bobrov, 1957а).
Tanacetum akinfievii (Alexeenko) Tzvel. (Asteraceae) – локальный эндемик известняковых массивов среднегорного Дагестана. В секции Xantoglossa (DC.) Sch. Bip. образует монотипный ряд Integrifolia Sosn. По мнению Н.Н. Цвелева (Tsvelev, 1961б), вид не имеет близкого родства с другими кавказскими видами рода и заслуживает выделения в особую монотипную секцию. Близок к малоазиатскому T. pectinatum Hausskn., также имеющему желтые язычковые цветки и слабо рассеченные листья.
Jurinea woronowii Iljin (Asteraceae) – эндемик хребтов Абишира-Ахуба, Чилик в верховьях реки Малый Уруп. Относится к монотипному ряду Woronowianae Nemirova подсекции Mamillosa Nemirova, все виды которой – локальные эндемики Северного Кавказа. Родство – с эукавказским J. pseudoiljinii Galushko et Nemirova – эндемиком верховий Большой Лабы (река Кислинка) и J. iljinii Grossh. – эндемиком г. Ацгара (Nemirova, 1999).
J. bellidioides Boiss. (Asteraceae) – эндемик ущелья р. Ардон в области Скалистого хребта (монотипный ряд Micrachaeniae Iljin ex Nemirova); J. brachypappa Nemirova – эндемик ущелья р. Фиагдон в области этого же хребта (монотипный ряд Brachypappae Nemirova); J. annae Sosn. – эндемик также Скалистого хребта в восточной части Центрального Кавказа (ущелья рек Терек и Асса, монотипный ряд Annae Nemirova); J. galushkoi Nemirova – эндемик Скалистого хребта, ущелье р. Урух (монотипный ряд Galushkoanae Nemirova); J. albovii Galushko et Nemirova – эндемик Фишт-Оштенского горного массива (монотипный ряд Albovianae Nemirova). Эти пять представителей монотипных рядов подсекции Coronopifoliae Nemirova, обособленные в системе рода, близкородственны другим представителям подсекции, а также локальным эндемикам Северного Кавказа; ареалы лишь небольшой части видов выходят на южный макросклон (Немирова, 1999).
Psephellus boissieri (Sosn.) Sosn. (Asteraceae) – эндемик известняковых массивов Внутригорного Дагестана, ареал его простирается от Ботлиха до Гуниба; P. galushkoi Alieva – эндемик среднегорного Дагестана, верховий бассейна р. Халагорк; P. andinus Galushko et Alieva – эндемик Андийского хребта. Виды входят в состав кавказской секции Psephellus, где образуют монотипные ряды, проявляя родство с другими видами этой секции, и, в большинстве своем, локальными эндемиками Восточного Кавказа (Aliyeva, 1979, 1983).
Как видно из рис. 3, ареалы большинства эндемиков, представителей монотипных рядов, расположены в центральной и восточной частях Северного Кавказа и не выходят за пределы северного макросклона, в западной части всего три вида. Также большинство видов (11) имеют ближайших родственников на Большом Кавказе. Это виды рода Jurinea Cass., Psephellus Cass. и два вида рода Pyrethrum Zinn. Ареалы родственников остальных видов расположены в прилегающих территориях – Закавказье, Малой Азии, Карпатах.
Схизоэндемики олиготипных (дитипных) секций и рядов, имеющие не менее важное значение для установления родственных связей и начальных этапов флорогенеза, также относятся к древнейшим типам. Таких видов 11 (рис. 4):
Рис. 4.
Ареалы представителей дитипных секций и рядов флоры ксерофитов Северного Кавказа.
Fig. 4. Ranges of representatives of ditypic sections and series of xerophytic flora of the Northern Caucasus.
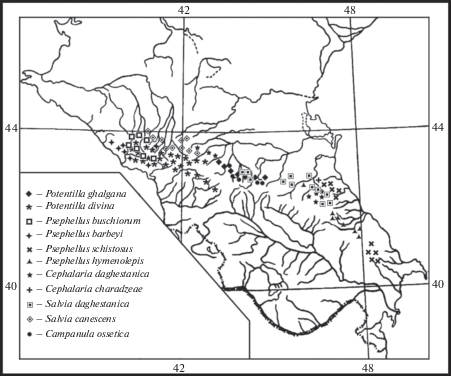
Potentilla ghalgana Juz. (Rosaceae) – эндемик высокогорий Скалистого хребта от бассейна р. Чегем до верховий р. Ассы. Входит в состав дитипной эукавказской секции Speciosae Th. Wolf, где близок к эукавказскому эндемику Potentilla divina Albov. Ближайшие родственники – представители монотипных секций P. elatior Willd. ex Schlecht. и P. brachypetala Fisch. et C.A. Mey. В целом подрод Fragariastrum Ser., куда входят перечисленные виды, является в основном кавказским, за исключением двух представителей еще одной тритипной секции Campestres Poeverl., ареалы которых захватывают Среднюю и Южную Европу (P. alba L.), Средиземноморье и Закавказье (P. micrantha Ramon) (Yuzepchuk, 1941).
Psephellus schistosus (Sosn.) Alieva (Asteraceae) – субэндемик, эндемик восточной части Северного Кавказа, распространенный в северо-восточной части Дагестана и Северном Азербайджане; P. hymenolepis (Trautv.) Boiss. – эндемик сланцевого Дагестана. Виды дитипной эндемичной восточнокавказской секции Schistosi Alieva, (наиболее древней в роде, поскольку ее виды имеют двойной хохолок), по этому признаку близки к представителям переднеазиатской секции Psephelloideae (Boiss.) Sosn. подрода Jacea (Juss.) Hayek. рода Centaurea L. По всем остальным признакам они вполне сходны с видами рода Psephellus Cass. Что касается хохолка, то он опадающий, а это характерно только для рода Psephellus Cass. Виды близки к представителям кавказской секции Psephellus, в первую очередь, к эндемику Внутригорного Дагестана P. boissieri (Sosn.) Sosn. (Aliyeva, 1979).
Psephellus buschiorum Sosn. (Asteraceae) – субэндемик, основной ареал которого расположен в ущельях Малой и Большой Лабы, часть ареала заходит в Абхазию; P. barbeyi Albov – также субэндемик, основной ареал которого сосредоточен в Западном Закавказье от ущелья р. Шахе до ущелья р. Бзыби. На северный макросклон ареал заходит в верховья рек Малая и Большая Лаба, Уруп. Представители дитипной кавказской секции Buschiani Mikheev, близки к представителям секции Hypoleuci Sosn., все виды которой – кавказские эндемики (Mikheev, 2008).
Cephalaria charadzeae Schchian (Dipsacaceae) – эндемик замкнутого ущелья р. Гимри в Центральном Дагестане; C. daghestanica Bobr. – эвриэндемик, основной ареал которого находится во Внутригорном Дагестане, отдельные фрагменты ареала имеются в ущельях рек Урух, Баксан, Малка. А.Д. Михеев (Mikheev, 2008) не признает самостоятельности C. charadzeae Schchian, считая его экологическим типом, находящимся внутри ареала C. daghestanica Bobr. Однако, по мнению А.С. Шхиян (Shkhiyan, 1965), этот вид довольно резко отличается от последнего прутьевидными, тонкими, многочисленными стеблями, опушенными только у основания (в отличие от одиночных, толстых, густо опушенных по всей длине. Оба вида входят в дитипный ряд Simplices Bobr., близкий к видам тритипного ряда Microcephalae Bobr., из которых один – закавказский эндемик (C. armeniaca Bordz.), один – армяно-иранский (C. pilosa Boiss. et Huet) и один – североирано-копетдагский (C. sublanata (Bornm) Szabo) (Bobrov, 1957б).
Campanula ossetica Bieb. (Campanulaceae) – эндемик Скалистого хребта (от ущелья р. Урух до ущелья р. Ассы). Входит в состав дитипного ряда Finitimae Fed., подсекции Symphyandriformes (Fomin) Fed. вместе с малоазиатским C. finitima Fomin. Эти два вида близки к видам рода Symphiandra A.DC., их объединяющим признаком является строение чашечки, у которой наблюдается складка между зубцами, иногда переходящая в более или менее длинный отросток; чашечка отстоит от венчика, с отвисающими вниз долями. Исходя из этого, представителей Symphyandriformes (Fomin) Fed. относят к древнейшим типам рода Campanula L. (Kharadze, 1949). Родственные связи прослеживаются с представителями ряда Raddeanae Fed. этой же подсекции, все виды которой – закавказские эндемики (Fedorov, 1957а).
Salvia canescens C.A. Mey. (Lamiaceae) – эвриэндемик центральной части Северного Кавказа, распространенный на Боковом и Скалистом хребтах от ущелья р. Большая Лаба до ущелья р. Баксан, заходит на меловые хребты окрестностей Кисловодска; S. daghestanica Sosn. – эвриэндемик, ареал которого охватывает Скалистый хребет в ущельях рек Ардон-Фиагдон, Шароаргун, далее Внутригорный и Среднегорный Дагестан. Образуют дитипный ряд Canescentes Pobed. подсекции Homalosphaceae (Bunge) Briq., где близки к видам памиро-алайского эндемичного дитипного ряда Seravschanicae Pobed. (Pobedimova, 1954).
ЗАКЛЮЧЕНИЕ
Таким образом, наиболее обособленные систематические реликты – представители монотипных родов, секций и рядов – проявляют родственные связи не с видами восточного и западного Древнего Средиземья, а с наименее обособленными видами, входящими в состав дитипных секций и рядов, распространенными на территориях, прилегающих к Кавказу. Все они относятся к категории палеоэндемиков, анцестральные формы которых мигрировали на территорию Северного Кавказа в первую волну миграции в верхнем миоцене-нижнем плиоцене. Часть видов сохранили родственные связи со среднеазиатским и средиземноморским видообразовательными центрами, где сегодня обитают их родственные виды, другая часть эти связи утратила. Это можно объяснить разной скоростью протекания видообразовательных процессов у разных таксонов: одни виды эволюционировали медленнее и сохранили отдаленные родственные связи, другие развивались более интенсивно и эти связи утратили, особенно это касается представителей таких родов, как Psephellus Cass., Jurinea Cass. и, отчасти, Campanula L., сформировавшие кавказские видообразовательные центры, давшие большое количество локальных эндемиков, родственные связи которых не выходят за пределы Кавказа. Эти процессы происходят в условиях высокогорий, эту парадоксальную особенность отмечал еще С.С. Харкевич (Kharkevitch, 1954).
Следует также отметить, что таксономические категории вышеупомянутых таксонов заставляют предположить разновозрастность формирования видов: самыми древними являются представители монотипных (дитипных) родов, затем секций и рядов. Это прослеживается в проявлении родственных связей: монотипные роды и секции проявляют таковые с отдаленными среднеазиатским и средиземноморским центрами формирования ксерофитов, а монотипные ряды – со вторичными иранским и малоазиатским центрами. Исключение составляет пара видов Salvia canescens C.A. Mey. – Salvia daghestanica Sosn. (дитипный ряд), проявляющие среднеазиатские связи.
Исходя их вышесказанного можно сделать вывод о том, что начальные этапы флорогенеза ксерофильной флоры Российского Кавказа проходили на основе двух миграционных волн: следы первой волны остались в виде систематических реликтов ранга рода и представителей монотипных секций и рядов, предки которых мигрировали из древнесредиземноморских отдаленных центров образования ксерофитов, источником второй волны миграции явились прилежащие к территории Кавказа вторичные центры образования ксерофильной флоры, которая стала основой формирования систематически менее обособленных таксонов – представителей олиготипных секций и рядов.
Список литературы
[Aliyeva] Алиева А.А. 1979. Новая секция рода Psephellus Cass. (Asteraceae). – В кн.: Флора Северного Кавказа и вопросы ее истории. Вып. 3. Ставрополь. С. 63–69.
[Aliyeva] Алиева А.А. 1983. Систематический обзор восточнокавказских видов рода Psephellus Cass. (Asteraceae Dum.). – В кн.: Флора Северного Кавказа и вопросы ее истории. Вып. 4. Ставрополь. С. 17–55.
[Bobrov] Бобров Е.Г. 1957а. Род Скабиоза – Scabiosa L. – В кн.: Флора СССР. Т. XXIV. М., Л. С. 56–91.
[Bobrov] Бобров Е.Г. 1957б. Род Головчатка – Cephalaria Schrad. – В кн.: Флора СССР. Т. XXIV. М., Л. С. 25–49.
[Borisova] Борисова А.Г. 1939. Род Умбиликус – Umbilicus DC. – В кн.: Флора СССР. Т. IX. М., Л. С. 13–14.
[Borisova] Борисова А.Г. 1959. Род Кладохета – Cladochaeta DC. – В кн.: Флора СССР. Т. XXV. М., Л. С. 430–432.
[Borisova] Борисова А.Г. 1969. Конспект системы сем. Crassulaceae DC. – Новости систематики высших растений. Т. 6. С. 112–121.
Favarger C. 1961. Contandriopoulus J. Essai sur l’endemisme. – Bull. Soc. Bot. Suisse. 71: 384–408.
[Fedorov] Федоров Ан.А. 1952. Род Первоцвет – Primula L. – В кн.: Флора СССР. Т. XVIII. М., Л. С. 111–202.
[Fedorov] Федоров Ан.А. 1957а. Род Колокольчик – Campanula L. – В кн.: Флора СССР. Т. XXIV. М., Л. С. 133–323.
[Fedorov] Федоров Ан.А. 1957б. Род Эдрайант – Edrajanthus A. DC. – В кн.: Флора СССР. Т. XXIV. М., Л. С. 442–444.
[Galushko] Галушко А.И. 1976. Анализ флоры западной части Центрального Кавказа. – В кн.: Флора Северного Кавказа и вопросы ее истории. Вып. 1. Ставрополь. С. 5–130.
[Galushko] Галушко А.И. 1979. Новый вид рода Psephellus Cass. (Asteraceae) с Северного Кавказа. – В кн.: Флора Северного Кавказа и вопросы ее истории. Вып. 3. Ставрополь. С. 58–62.
[Goncharova] Гончарова C.Б. 2006. Подсемейство Sedoideae (Crassulaceae) флоры Сибири и Российского Дальнего Востока (систематика, биоморфология, филогения). Дис…. докт. биол. наук. Владивосток. 301 с.
[Ivanov] Иванов А.Л. 1998. Флора Предкавказья и ее генезис. Ставрополь. 204 с.
[Kamelin] Камелин Р.В. 2004. Лекции по систематике растений. Главы теоретической систематики растений. Барнаул. 226 с.
[Khanjyan] Ханджян Н.С. 2008. Tanacetum L. – В кн.: Конспект флоры Кавказа. Т. 3 (1). СПб., М. С. 206–212.
[Kharadze] Харадзе А.Л. 1949. Опыт систематики кавказских видов рода Campanula L. секции Medium A. DC. – Заметки по систематике и географии растений. Вып. 15. Тбилиси. С. 14–35.
[Kharkevitch] Харкевич С.С. 1954. Роль четвертичного эпейрогенеза в формировании высокогорной флоры Большого Кавказа. – Бот. журн. 39(4): 498–514.
[Klokov] Клоков М.В. 1954. Род Тимьян – Thymus L. – В кн.: Флора СССР. Т. XXI. М., Л. С. 470–591.
[Kolakovsky] Колаковский А.А. 1995. Семейство Колокольчиковых. М. 92 с.
[Linchevsky] Линчевский И.А. 1952. Род Кермековидка. – Limoniopsis Lincz. – В кн.: Флора СССР. Т. XVIII. М., Л. С. 376–378.
[Linchevsky] Линчевский И.А. 1958. Род Ложнобецкея – Pseudobetckea (Hock) Lincz. – В кн.: Флора СССР. Т. XXIII. М., Л. С. 681–682.
[Menitsky] Меницкий Ю.Л. 2008. Cladochaeta DC. – В кн.: Конспект флоры Кавказа. Т. 3(1). СПб., М. С. 186.
[Mikheev] Михеев А.Д. 2008. Psephellus Cass. – В кн.: Конспект флоры Кавказа, Т. 3(1). СПб., М. С. 287–296.
[Nemirova] Немирова Е.С. 1999. Род Jurinea Cass. флоры Северного Кавказа. Ставрополь. 183 с.
[Pobedimova] Победимова Е.Г. 1954. Род Шалфей – Salvia L. – В кн.: Флора СССР. Т. XXI. М., Л. С. 244–374.
[Prima] Прима В.М. 1974. О некоторых видах рода Pyrethrum Zinn с Восточного Кавказа. – Новости систематики высших растений. 11: 277–287.
[Shkhiyan] Шхиян А.С. 1965. Новые данные к изучению рода Cephalaria Schrad. – Заметки по систематике и географии растений. Вып. 25. Тбилиси. С. 46–48.
[Tsvelev] Цвелев Н.Н. 1961а. Род Пиретрум (Ромашник, Поповник) – Pyrethrum Zinn. – В кн.: Флора СССР. Т. XXVI. М., Л. С. 185–261.
[Tsvelev] Цвелев Н.Н. 1961б. Род Пижма – Tanacetum L. – В кн.: Флора СССР. Т. XXVI. М., Л. С. 317–359.
[Yuzepchuk] Юзепчук С.В. 1941. Род Лапчатка – Potentilla L. – В кн.: Флора СССР. Т. X. М., Л. С. 78–223.
Дополнительные материалы отсутствуют.
Инструменты
Ботанический журнал