

Ботанический журнал, 2020, T. 105, № 12, стр. 1169-1190
КРОВОХЛЕБКОВЫЕ ПОЙМЕННЫЕ ЛУГА НА ЕВРОСИБИРСКОМ ДОЛГОТНОМ ГРАДИЕНТЕ
И. Б. Кучеров 1, *, К. В. Щукина 1, **, И. В. Татаренко 2, Т. А. Паринова 3, ***, А. Г. Волков 3, Е. С. Нескрябина 4, О. В. Чередниченко 5, ****, Н. П. Савиных 6, О. Н. Пересторонина 6, С. В. Шабалкина 6, Е. М. Пыжикова 7, М. Г. Цыренова 7
1 Ботанический институт им. В.Л. Комарова РАН
197376 Санкт-Петербург, ул. Проф. Попова, 2, Россия
2 The Open University
MK7 6AA Milton, Keynes,
Walton Hall, Kents Hill, United Kingdom
3 Северный (Арктический) федеральный университет им. М.В. Ломоносова
163002 Архангельск, наб. Северной Двины, д. 17, Россия
4 Хоперский государственный природный заповедник
397418 Воронежская обл., п. Варварино, Новохоперский р-н, ул. Лесная, д. 61, Россия
5 Московский государственный университет им. М.В. Ломоносова
119991 Москва, Ленинские Горы, д. 1, Россия
6 Вятский государственный университет
610020 Кировская обл., Киров, ул. Московская, д. 36, Россия
7 Бурятский государственный университет им. Доржи Банзарова
670000 Республика Бурятия, Улан-Удэ, ул. Смолина, д. 24а, Россия
* E-mail: atragene@mail.ru
** E-mail: vyatka_ks_72@mail.ru
*** E-mail: nadeinata@mail.ru
**** E-mail: sciapoda@mail.ru
Поступила в редакцию 18.05.2020
После доработки 15.08.2020
Принята к публикации 25.08.2020
Аннотация
Приводятся результаты флористико-геоботанических исследований пойменных лугов с доминированием Sanguisorba officinalis L. на долготном градиенте в пределах Евразии, включая Англию, Европейскую Россию и Восточную Сибирь. Несмотря на азональность, состав и структура ценофлор пойменных кровохлебковых лугов демонстрируют зональные, а также секторальные черты, чему способствует протяженный ареал этих сообществ. “Ядро” флороценотического комплекса кровохлебковых лугов формируют евразиатские полизональные мезоэвтрофные мезофиты и гигромезофиты. Видовое богатство кровохлебковых лугов возрастает как в условиях автохтонного генезиса ценофлоры, так и в океаническом климате. Доминирование Sanguisorba officinalis на лугах в пойме р. Темзы способствует увеличению видовой насыщенности травостоя этих лугов. Сенокошение позитивно влияет на поддержание устойчивой продуктивности кровохлебковых лугов в долгосрочной перспективе.
ГЕОГРАФИЧЕСКОЕ РАСПРОСТРАНЕНИЕ И ИЗМЕНЧИВОСТЬ ЦЕНОТИЧЕСКОЙ ПРИУРОЧЕННОСТИ SANGUISORBA OFFICINALIS
Кровохлебка лекарственная (Sanguisorba officinalis L.) – полизональный вид с евразиатско-западноамериканским аборигенным ареалом, широко распространенный в северных и умеренных широтах Северного полушария (Hultén, Fries, 1986). Ареал S. officinalis имеет значительную протяженность на территории Европейской России. Этот вид произрастает во всех областях центрально-европейской части России, изреживаясь к северу (Maevskii, 2006) и лишь спорадически встречаясь в области бывшего Валдайского оледенения. Северная граница распространения S. officinalis на северном макросклоне Русской равнины идет от г. Архангельска по р. Сояне, далее южнее п-ова Канин вплоть до р. Большая Ою в Ненецком автономном округе, где вид выходит на побережье Баренцева моря около 68° с.ш.. На Урале, в Сибири южнее (а местами и севернее) Полярного круга, а также на севере азиатской части Евразии вид повсеместен на характерных для него экотопах. На западе Северной Америки S. officinalis распространена от Аляски до Калифорнии, в том числе по побережью Британской Колумбии. В горах она поднимается до высоты 3000 м над ур. м. (Yuzepchuk, 1941; Atlas…, 1980; Pojar et al, 1994; Yakubov, 1996; Li et al., 2003; Khela, 2013; Phipps, 2014).
S. officinalis встречается и во всех странах Западной и Центральной Европы, кроме Португалии и островов Средиземноморья (Nordborg, 1963; Kurtto, 2009; Khela, 2013). Это аборигенный вид флоры Британских островов. Чаще он произрастает в центральных и северных, реже в южных районах Англии, а также в Южном Уэльсе. В Ирландии вид редок (Preston et al., 2002).
По положению своих экологических оптимумов на градиентах богатства и увлажнения почвы S. officinalis – мезоэвтрофный гигромезофит. Однако она может произрастать на почвах от бедных до среднезасоленных при увлажнении от мезоксерофитного до мезогигрофитного (Ramensky et al., 1956). При оптимуме в теплоумеренном и умеренно континентальном климате (Ellenberg et al., 1992) она выдерживает условия от холодных субальпийских до умеренно жарких и от океанических до резко континентальных (Atlas …, 1980).
Широкая экологическая амплитуда S. officinalis позволяет ей заселять различные местообитания. Ее можно встретить по берегам озер и рек, на низинных болотах, в прибрежных и пойменных кустарниках. Вид отмечен также по опушкам мелколиственных (как в южной тайге юго-востока Ленинградской обл.) и даже темнохвойных (как в горной средней тайге хребта Басеги в Пермском крае) лесов. На Полярном (Soczava, 1927) и Северном (Degteva, Dubrovskii, 2014) Урале он нередко достигает значимого обилия под пологом подгольцовых еловых редколесий и березовых криволесий. В то же время S. officinalis может выступать пионерным видом, заселяющим отмели, песчаные и гравийные косы в прирусловой пойме Енисея (Nomokonov, 1959), Верхней и Средней Печоры и других рек Урало-Сибирского региона в силу приспособленности к условиям переменного увлажнения. В качестве сопутствующего вида кровохлебка обычна в остепненных дриадовых (из Dryas punctata) тундрах и реликтовых криофитных степях Западной Чукотки (Yurtsev, 1981). Она может расти и по обочинам дорог, а также на залежах. Но наиболее характерны для S. officinalis сообщества лугов – от сухих остепненных до заболоченных, в том числе солонцеватых (как, например, по Карельскому берегу Белого моря).
Изменчивость ценотической приуроченности S. officinalis обусловлена как жизненной формой этого вида (стержнекорневой поликарпик с многоглавым каудексом с большим запасом спящих почек (Nordborg, 1963; Ermakova, 1983; Zhukova, 1997)), позволяющей ему произрастать при нестабильном увлажнении, так и кариологическим полиморфизмом. Среди многих выявленных цитотипов (IPCN…, 2020) наиболее широко распространены тетра- (2n = 28) и октоплоиды (2n = 56). Первый из цитотипов характерен для регионов с экстремальными либо переменными экологическими режимами, включая Арктику и Сибирь, отмечен также в Японии. Октоплоиды свойственны более стабильным экологическим условиям, в том числе атлантическому или субатлантическому климату. Они преобладают на лугах, развитых на известняках о-ва Готланд, отмечены в Англии, Германии, Чехии, Беларуси, но также на юге Западной Сибири и на Дальнем Востоке (Nordborg, 1963; Chisla…, 1993; IPCN…, 2020).
Максимального обилия S. officinalis достигает на пойменных лугах, где переменность увлажнения сочетается с повышенным уровнем богатства почвы и выравненным микроклиматом (Walter, 1968). На протяжении многих веков развития европейской цивилизации пойменные луга благодаря их высокой естественной продуктивности служили важнейшими сенокосными угодьями (Klaap, 1941; Lepkovich, 2005; Rothero et al., 2016). При этом в силу значительной продолжительности жизни отдельных особей (на Залидовских лугах в пойме р. Угры (Калужская обл.) – до 75 л. (Ermakova, 1983)) даже на лугах, где не проводится сенокошение, кровохлебка удерживается намного дольше других видов двудольных. Тем не менее, в отсутствие сенокошения кровохлебковые луга зарастают кустарниковыми ивами (Tyurin, 2015).
В Англии S. officinalis выступает диагностическим видом среднепойменных лисохвостово-кровохлебковых лугов (MG4: Alopecurus pratensis–Sanguisorba officinalis floodplain meadow), когда-то распространенных по всей стране, но к настоящему времени сохранившихся лишь в пойме р. Темзы и ряда других рек (Rodwell, 1992; Jefferson, 1997; Rodwell et al., 2007; Floodplain…, 2014). С развитием интенсивного земледелия в поймах эти луга во второй половине XX в. исчезли более чем на 98% территории, исходно занятой ими в Великобритании (Jefferson, 1997). В Европе сообщества Alopecurus pratensis–Sanguisorba officinalis-типа признаны находящимися на грани исчезновения и взяты под особую охрану (Council Directive…, 1992; Zelnik, Carni, 2013). В то же время аналогичные луга до сих пор широко распространены на территории России, однако их изучение до недавнего времени почти не проводилось.
Целью данной публикации является сравнительный анализ видового состава, структуры ценофлор и продуктивности пойменных кровохлебковых лугов, описанных в разных частях Евразии – в Англии, Европейской России и Восточной Сибири.
ИСПОЛЬЗОВАННЫЕ ДАННЫЕ И МЕТОДЫ
В основу работы положена выборка из 102 геоботанических описаний, выполненных в 2016 г. на лугах в поймах рек Темза (Англия; данные И.В. Татаренко), Хопер (Россия, Воронежская обл.; данные Е.С. Нескрябиной), Северная Двина (в низовьях; Архангельская обл.; данные Т.А. Париновой), Вятка (Кировская обл.; данные К.В. Щукиной, Н.П. Савиных, О.Н. Пересторониной и С.В. Шабалкиной) и Байса (приток р. Большой Амалат; Республика Бурятия; данные Е.М. Пыжиковой и М.Г. Цыреновой). Совокупную последовательность всех пяти географических пунктов можно интерпретировать как евросибирский долготный профиль общей протяженностью 7000 км. С целью унификации методики с общепринятой в Великобритании все описания выполнены на площадках по 1 м2. На каждой из них выявлен видовой состав сосудистых растений с оценкой обилия каждого вида и измерена брутто-продуктивность травостоя.
Для оценки видового (α-) разнообразия (Whittaker, 1972) луговых сообществ использованы их общее видовое богатство и видовая насыщенность на 1 м2 (Markov, 1962). Оценки видовых обилий унифицированы как проективные покрытия (%) с помощью трансформационных алгоритмов информационной ботанической системы ИБИС 7.2 (Zverev, 2007; Kucherov, 2019). Сводные данные по всем обследованным участкам вместе с экологическими, хорологическими и биоморфными характеристиками произрастающих на них видов сведены в фитоценотическую таблицу (табл. 1).
Таблица 1.
Эколого-фитоценотическая характеристика пойменных кровохлебковых лугов на евросибирском долготном градиенте Table 1. Ecological and phytocoenotical record of great burnet floodplain meadows along the Eurosiberian longitudinal gradient
| Вид/Species | ЖФ | Гео- и экоэлементы/ Range and ecological types | Поймы рек / Floodplains of rivers | |||||||
|---|---|---|---|---|---|---|---|---|---|---|
| I | II | III | IV | 1 | 2 | 3 | 4 | 5 | ||
| Детерминанты географических пунктов/Determinants of geographical points | ||||||||||
| Lotus corniculatus s.l. | Cткр | ПЛ | Е-Др | Э | М2 | 763 | 10+ | 6+ | ||
| Festuca rubra | РД | ПЛ | Гол | МЭ | М1 | 10011 | 61 | 81 | ||
| Ranunculus acris | КК | БН | ЕС | МЭ | М2 | 1001 | 61 | |||
| Trifolium pratense | Сткр | ПЛ | ЕС-Др | МЭ | М2 | 412 | 102 | |||
| Agrostis stolonifera | Стол | ПЛ | ЕА | МЭ | МГ | 9316 | ||||
| Poa trivialis | РД | БН | ЕА | МЭ | ГМ | 9310 | ||||
| Holcus lanatus | РД | Н | Е | М | М1 | 932 | ||||
| Centaurea nigra | КК | Н | Е | М | М2 | 794 | ||||
| Carex flacca | ДК | Н | Е-Др | Э | Г | 614 | ||||
| Cynosurus cristatus | Д | Н | Е | МЭ | М1 | 722 | ||||
| Silaum silaus | Сткр | ЛС | ЕС-Др | Э | М1 | 722 | ||||
| Anthoxanthum odoratum | РД | БН | ЕС-Др | М | М2 | 721 | ||||
| Arrhenatherum elatius | РД | ПЛ | Е-Др | МЭ | М1 | 693 | ||||
| Plantago lanceolata | Сткр | ПЛ | ЕС-Др | МЭ | М1 | 691 | ||||
| Succisa pratensis | КК | БН | ЕС-Др | ОМ | М2 | 6621 | ||||
| Deschampsia cespitosa | ПД | ПЛ | ЕА | МЭ | ГМ | 553 | ||||
| Carex panicea | ДК | ПЛ | Е-Др | МЭ | ГМ | 483 | ||||
| Primula veris | КК | Н | Е | МЭ | М2 | 483 | ||||
| Hordeum secalinum | РД | Н | Е-Др | Э | ГМ | 482 | ||||
| Luzula campestris | РД | Н | Е-Др | ОМ | М2 | 38+ | ||||
| Bromus racemosus | МО | Н | Е-Др | Э | ГМ | 34+ | ||||
| Trisetum flavescens | РД | Н | Е-Др | МЭ | М2 | 31+ | ||||
| Phleum pratense | РД | БН | ЕС-Др | МЭ | М2 | 79+ | 445 | |||
| Taraxacum agg. officinale | Сткр | ПЛ | ЕС | Э | М2 | 551 | 283 | |||
| Dactylis glomerata | РД | БН | ЕС-Др | МЭ | М2 | 59+ | 112 | |||
| Briza media | РД | ПЛ | Е-Др | М | М2 | 411 | 11+ | |||
| Filipendula ulmaria s.l. | КК | БН | ЕС-Др | МЭ | ГМ | 4510 | 8611 | 8314 | 307 | |
| Rumex acetosa s.l. | КО, КК | БН | ЕА | МЭ | ГМ | 34+ | 173 | 405 | ||
| Equisetum palustre | ДК | ПЛ | Гол | МЭ | МГ | 38+ | 10+ | 508 | ||
| Galium verum s.l. | ДКС, ДК | ЛС | Гол | МЭ | М1 | 24+ | 583 | |||
| G. physocarpum | ДКС | БН | ЕС-Др | МЭ | М2 | 864 | 175 | |||
| Kadenia dubia | ММ | ПЛ | ЕС-Др | МЭ | ГМ | 862 | ||||
| Inula britannica | КО | ПЛ | ЕА | Э | М2 | 812 | ||||
| Poa palustris | РД | ПЛ | Гол | МЭ | ГМ | 712 | ||||
| Euphorbia palustris | ДК | ЛС | Е-Др | Э | МГ | 5212 | ||||
| Glechoma hederacea | НП | БН | ЕС-Др | МЭ | М2 | 482 | ||||
| Gratiola officinalis | КК | ЛС | Гол | МЭ | ГМ | 292 | ||||
| Symphytum tauricum | Сткр | ЛС | Е-Др | МЭ | М2 | 291 | ||||
| Alopecurus pratensis | ДК | БН | ЕА | М | М2 | 17+ | 718 | 505 | 704 | |
| Bromopsis inermis | ДК | ПЛ | ЕС-Др | Э | М2 | 195 | 409 | 42 | ||
| Carex praecox | ДК | ЛС | ЕА | Э | М1 | 7620 | 807 | |||
| Ranunculus agg. auricomus | КК | Б | ЕС | МЭ | М2 | 291 | 50+ | |||
| Rumex thyrsiflorus | Сткр | ПЛ | ЕС | МЭ | М2 | 29+ | 403 | |||
| Geranium pratense | КК | БН | ЕА | МЭ | М2 | 7210 | 203 | 131 | ||
| Elytrigia repens | ДК | ПЛ | Е-Др | Э | М2 | 14+ | 509 | |||
| Thalictrum simplex | ДК | ПЛ | ЕА | МЭ | М2 | 8918 | 81 | |||
| Equisetum arvense | ДК | ПЛ | Гол | МЭ | М2 | 508 | ||||
| Galium boreale | ДКС, ДК | ПЛ | Гол | МЭ | М2 | 10028 | 10013 | 332 | ||
| Veronica longifolia | ДК | ПЛ | ЕА | МЭ | МГ | 19+ | 707 | 211 | ||
| Allium angulosum | ЛК | ЛС | ЕС | Э | ГМ | 14+ | 40+ | |||
| Cenolophium denudatum | ММ | ЛС | ЕС | МЭ | ГМ | 60+ | 131 | |||
| Inula salicina | ДК | ПЛ | ЕА | МЭ | М1 | 8012 | ||||
| Viola canina | КК | Б | ЕС-Др | М | М2 | 501 | ||||
| Equisetum pratense | ДК | Б | Гол | М | М2 | 40+ | ||||
| Ptarmica vulgaris | КК | ПЛ | Е | МЭ | ГМ | 303 | ||||
| Iris sibirica | КК | ЛС | ЕС | МЭ | ГМ | 302 | ||||
| Geranium wlassovianum | КК | БН | А | МЭ | М2 | 796 | ||||
| Trollius vicarius | КК | ПЛ | А | МЭ | М2 | 637 | ||||
| Hedysarum alpinum | Сткр | Б | ЕС | М | М1 | 468 | ||||
| Artemisia integrifolia | КК | Б | С-Др | МЭ | М2 | 424 | ||||
| Seseli condensatum | ММ | ЛС | ЕА | МЭ | М2 | 294 | ||||
| Rumex gmelinii | Сткр | ПЛ | А | МЭ | ГМ | 291 | ||||
| Pedicularis resupinata | КК | БН | А | МЭ | М2 | 291 | ||||
| Helictotrichon hookeri | РД | Ст | С-Ам | МЭ | КМ | 251 | ||||
| Arctopoa subfastigiata | ДК | ПЛ | А | МЭ | М1 | 251 | ||||
| Константные виды/Constants | ||||||||||
| Sanguisorba officinalis | Сткр | ПЛ | ЕАзАм | МЭ | ГМ | 8629 | 6223 | 10038 | 10029 | 714 |
| Lathyrus pratensis | ДК | ПЛ | ЕА | МЭ | М2 | 79+ | 437 | 616 | 704 | 251 |
| Vicia cracca | ДК | ПЛ | ЕА | МЭ | М2 | 55+ | 573 | 8314 | 1004 | 6710 |
| Прочие виды/Companions | ||||||||||
| Lysimachia nummullaria | НП | Н | Е | Э | М2 | 141 | 292 | 203 | ||
| Carex acuta | ДК | ПЛ | ЕС-Др | МЭ | Г | 71 | 14+ | 10+ | ||
| Poa pratensis | ДК | ПЛ | Гол | МЭ | М2 | 24+ | 302 | 291 | ||
| Carex cespitosa | РД | ПЛ | ЕА | МЭ | МГ | 223 | 10+ | 82 | ||
| Thalictrum flavum | ДК | ПЛ | ЕС-Др | МЭ | ГМ | 171 | 14+ | |||
| Festuca pratensis | РД | БН | ЕС-Др | МЭ | М2 | 141 | 172 | |||
| Cirsium arvense s.l. | ДКС, КО | ПЛ | Е-Др | МЭ | М2 | 101 | 111 | |||
| Eleocharis palustris | ДК | ПЛ | Гол | Э | Г | 245 | 10+ | |||
| Trifolium medium | ДКС, Сткр | ПЛ | ЕС | МЭ | М1 | 14+ | 202 | |||
| Carex vulpina | РД | ПЛ | ЕС-Др | МЭ | Г | 14+ | 10+ | |||
| Lysimachia vulgaris | ДК | ПЛ | ЕС-Др | МЭ | МГ | 14+ | 10+ | |||
| Rubus caesius | К | ПЛ | ЕС-Др | МЭ | М2 | 31 | 202 | |||
| Veratrum lobelianum | КК | Б | ЕА | МЭ | ГМ | 282 | 293 | |||
| Helictotrichon pubescens | РД | ПЛ | ЕС-Др | МЭ | М2 | 24+ | ||||
| Juncus acutiflorus | РД | ПЛ | Е-Др | ОМ | ГМ | 2413 | ||||
| Leucanthemum vulgare | КК | ПЛ | ЕА | МЭ | М1 | 24+ | ||||
| Lolium perenne | РД | ПЛ | Е-Др | МЭ | М2 | 21+ | ||||
| Agrostis tenuis | РД | ПЛ | ЕС-Др | М | М2 | 171 | ||||
| Leontodon hispidus | КК | ПЛ | Е-Др | М | М1 | 17+ | ||||
| Cerastium holosteoides | Сткр | ПЛ | ЕС-Др | М | М2 | 14+ | ||||
| Ajuga reptans | Стол | Н | Е-Др | М | М2 | 14+ | ||||
| Ranunculus repens | Стол | ПЛ | ЕА | Э | ГМ | 331 | ||||
| Calystegia sepium | ДК | ПЛ | ЕА | МЭ | ГМ | 24+ | ||||
| Fritillaria meleagroides | ЛК | Ст | Е | Э | М2 | 14+ | ||||
| Viola nemoralis | КК | Б | ЕС-Др | М | М2 | 14+ | ||||
| Lythrum virgatum | КК | ЛС | ЕС-Др | МЭ | МГ | 101 | ||||
| Centaurea phrygia | КК | Б | ЕС | М | М2 | 173 | ||||
| Hieracium agg. umbellatum | КК | ПЛ | Гол | М | М1 | 112 | ||||
| Campanula glomerata | КО, КК | ПЛ | ЕС-Др | МЭ | М1 | 111 | ||||
| Fragaria viridis | Стол | ЛС | ЕС-Др | МЭ | КМ | 203 | ||||
| Hylotelephium triphyllum | Кл | ПЛ | ЕА | МЭ | М1 | 20+ | ||||
| Genista tinctoria | ПКЧ | ЛС | ЕС | М | М1 | 102 | ||||
| Calamagrostis langsdorffii | ДК | Б | Гол | М | ГМ | 212 | ||||
| Carex enervis | РД | ПЛ | А | Э | ГМ | 212 | ||||
| Pedicularis sceptrum-carolinum | КК | Б | ЕС | М | МГ | 211 | ||||
| Saussurea amurensis | ДК | БН | А | МЭ | М2 | 211 | ||||
| Parnassia palustris | КК | Б | ЕАзАм | МЭ | ГМ | 21+ | ||||
| Thalictrum contortum | КК | Н | А | М | М2 | 173 | ||||
| Filipendula palmata | КК | БН | А | МЭ | М2 | 172 | ||||
| Polemonium chinense | КК | БН | А | МЭ | М2 | 17+ | ||||
| Carex vesicata | ДК | Б | С-Ам | МЭ | ГМ | 132 | ||||
| C. coriophora | РД | БН | А | МЭ | М2 | 131 | ||||
| Calamagrostis neglecta | ДК | Б | Гол | М | МГ | 131 | ||||
| Elymus jacutensis | РД | Б | С-Ам | М | М2 | 131 | ||||
| Salix rosmarinifolia | К | ПЛ | ЕС-Др | МЭ | МГ | 131 | ||||
| Ranunculus japonicus | КК | Н | А-Ам | МЭ | М2 | 131 | ||||
| Artemisia tanacetifolia | ДК | Б | ЕАзАм | МЭ | М1 | 131 | ||||
| Ligularia sibirica | КК | Б | ЕС | МЭ | ГМ | 131 | ||||
| Bistorta alopecuroides | КК | Н | А | МЭ | М2 | 13+ | ||||
| Spiraea salicifolia | К | ЛС | ЕАзАм | МЭ | ГМ | 13+ | ||||
| Potentilla fragarioides | Сткр | ПЛ | А | МЭ | М2 | 13+ | ||||
| Galium trifidum | Сткр | Б | ЕАзАм | М | Г | 13+ | ||||
| Среднее проективное покрытие (%)/Average projective cover (%) of: | ||||||||||
| – травяного яруса/herb layer | 00 | 95 | 95 | 84 | 64 | |||||
| – мохового яруса/moss layer | 0 | 0 | 0 | 0 | 0 | |||||
| – ветоши/grassland litter | ? | ? | 7 | 64 | ? | |||||
| – незадернованного аллювия/exposed alluvium | ? | ? | 8 | 4 | ? | |||||
| Видовое богатство/Total species richness | 56 | 51 | 34 | 40 | 66 | |||||
| Видовая насыщенность на 1 м2/Number of species per 1 m2 | 24 ± 5 | 14 ± 3 | 11 ± 1 | 15 ± 1 | 12 ± 1 | |||||
| – в том числе с доминирующей Sanguisorba officinalis/with dominant Sanguisorba officinalis | 26 ± 4 | 14 ± 2 | 11 ± 1 | 15 ± 1 | 11 ± 1 | |||||
| – с малочисленной или без S. officinalis/with few or lacking S. officinalis | 18 ± 2 | 13 ± 3 | 9 | ? | 13 ± 1 | |||||
| t-критерий Стьюдента/Student’s t-test | 1.99(*) | 0.68 | ? | ? | 0.92 | |||||
| Широта/Latitude | 51.79 | 51.19 | 64.24 | 57.28 | 52.17 | |||||
| Долгота/Longitude | –00.70 | 41.73 | 41.74 | 50.05 | 107.50 | |||||
| Сумма градусо-дней выше +10°С/Sum of growing degree-days above +10°С | 835 | 1301 | 410 | 738 | 491 | |||||
| Коэффициент континентальности Конрада/Conrad’s continentality index | 2.3 | 8.0 | 1.5 | 7.5 | 6.0 | |||||
| Число описаний/Number of relevés | 29 | 21 | 18 | 10 | 24 | |||||
| – в том числе с доминирующей Sanguisorba officinalis/with dominant Sanguisorba officinalis | 1 | 9 | 7 | 0 | 5 | |||||
| – с малочисленной или без S. officinalis/with few or lacking S. officinalis | 8 | 12 | 1 | 0 | 19 | |||||
Примечания. ЖФ – жизненные формы: К – кустарники, ПКЧ – полукустарники. Травянистые поликарпики: Сткр – стержнекорневые, ДКС – длиннокорневищно-стержнекорневые, КО – корнеотпрысковые, ДК, КК – длинно- и короткокорневищные, РД, ПД – рыхло- и плотнодерновинные, Стол – столонообразующие, НП – надземноползучие, Кл – клубневые, ЛК – луковичные. Монокарпики: МО – однолетние, ММ – многолетние.
I, II – геоэлементы: I – широтные: Б – бореальный, БН – бореонеморальный, Н – неморальный, ЛС – лесостепной, Ст – степной, ПЛ – полизональный; II – хориономические: Гол – голарктический, ЕАзАм – евразиатско-западноамериканский, ЕА – евразиатский, ЕС – евросибирский, ЕС-Др, Е-Др, С-Др – евросибирско-, европейско- и сибирско-древнесредиземноморский, Е – европейский, А – азиатский, С – сибирский, С-Ам – сибирско-американский.
III, IV – экоэлементы по отношению к: III – влажности почвы: КМ – ксеромезофиты, М1, М2 – мезофиты сухо- и влажнолугового увлажнения, ГМ – гигромезофиты, МГ – мезогигрофиты, Г – гигрофиты; IV – богатству почвы: ОМ – олигомезотрофы, М – мезотрофы, МЭ – мезоэвтрофы, Э – эвтрофы и галофиты.
Поймы рек: 1 – Темза, 2 – Хопер, 3 – Северная Двина, 4 – Вятка, 5 – Байса.
Для видов приводятся постоянство (%) и (в надстрочном регистре) среднее проективное покрытие (%) в конкретных географических пунктах. Группы детерминантов последних выделены серым фоном и жирной рамкой; виды в них сортированы по убыванию встречаемости и далее покрытия во всем массиве описаний таблицы. Низкие кустарники включены в травяной ярус. Покрытие и постоянство доминантов и субдоминантов дано полужирным шрифтом. Плюс “+” соответствует среднему проективному покрытию менее 0.5%. Знак вопроса “?” – отсутствие данных. Исключены виды со встречаемостью не более 20% в каждой из групп.
Для видовой насыщенности данные приведены в формате “среднее арифметическое ± стандартная ошибка среднего”. Значения t-критерия Стьюдента, достоверные на уровне тенденций (α≤10%), выделены полужирным шрифтом и звездочкой в скобках (*).
Notes. ЖФ – life forms: К – shrubs, ПКЧ – semishrubs. Polycarp herbs: Сткр – tap-rooted, ДКС – tap-rooted with long rhizomes, КО – ratoon-forming, ДК, КК – long- and short-rhizomatous, РД, ПД – loosely and firm-bunched, Стол – stolon-forming, НП – creeping, Кл – tuberous, ЛК – bulbous. Monocarps: МО – annual, ММ – perennial.
I, II – plant range types: I – latitudinal: Б – boreal, БН – boreal-nemoral, Н – nemoral, ЛС – forest-steppe, Ст – steppe, ПЛ – multizonal; II – chorionomic: Гол – Holarctic, ЕАзАм – Eurasian-Western American, ЕА – Eurasian, ЕС – Eurosiberian, ЕС-Др, Е-Др, С-Др – Eurosiberian-, European- and Siberian-Ancient Mediterranean, Е – European, А – Asian, С – Siberian, С-Ам – Siberian-American.
III, IV – plant ecological types in regard to: III – soil moisture: КМ – xeromesophytes, М1, М2 – mesophytes of dry and moist meadow conditions, ГМ – hygromesophytes, МГ – mesohygrophytes, Г – hygrophytes; IV – soil fertility: ОМ – oligomesotrophic, М – mesotrophic, МЭ – mesoeutrophic, Э – eutrophic and halophytic.
Floodplains of rivers: 1 – Thames, 2 – Khopyor, 3 – Northern Dvina, 4 – Vyatka, 5 – Baisa.
Constancy (%) and average cover (%) (in superscripts) are given for species from different geographical points. Groups of point determinants are marked with margins and shaded; species within them are sorted first by descending occurrence, then by cover in the whole table relevé set. Low shrubs are included into the herb layer. The values of cover and constancy of dominants are shown in bold. Average species cover under 0.5% is given as “+”. Question mark “?” means lacking data. Species with occurrence under 10% in all of the points are excluded from the table.
Numbers of species per 1 m2 are given as arithmetical means ± standard errors. The Student’s t-test values trustworthy at the confidence level of tendencies (α≤10%) are shown in bold and marked by asterisks in brackets (*).
Для сводных ценофлор пойменно-луговых фитоценозов в каждом из географических пунктов проведен сравнительный таксономический и типологический анализ (табл. 2), результаты которого частично опубликованы ранее (Parinova et al., 2019). При таксономическом анализе рассчитаны коэффициенты флористической общности Жаккара на уровне видов, родов и семейств, а также семейственно-видовые спектры (Tolmachev, 1974; Schmidt, 1984). Типологический – биоморфный, географический и экологический – анализ проведен с учетом среднего проективного покрытия видов, слагающих сообщества (Zverev, 2007; Degteva, Dubrovskii, 2014; Kucherov, 2019). В основу биоморфного анализа положена система жизненных форм И.Г. Серебрякова (Serebryakov, 1962) с уточнениями по Т.Г. Полозовой (Polozova, 1983, etc.; Kucherov et al., 2000). У некоторых видов выявлена более чем одна биоморфа (данные Н.П. Савиных; табл. 1). При географическом анализе рассчитаны спектры широтных (Tolmachev, 1974; Yurtsev, Kamelin, 2002) и хориономических (Yurtsev, Kamelin, 2002; Kamelin, 2018) элементов. В первом случае расчеты велись по В.М. Шмидту (Schmidt, 2005) с региональными уточнениями (Kucherov, 2016 b, 2019), во втором – по результатам соотнесения видовых ареалов (Hultén, Fries, 1986) с единицами системы флористического районирования Земли (Takhtajan, 1978; Kamelin, 2018). При построении спектров экоэлементов по отношению к влажности и богатству почвы использованы фитоиндикационные шкалы Г. Элленберга (Ellenberg et al., 1992) для Англии, Л.Г. Раменского (Ramensky et al., 1956) c уточнениями (Kucherov, 2019) для Русской равнины, И.А. Цаценкина (Tsatsenkin et al., 1978) для Бурятии. Зональная специфика географических пунктов оценена с помощью сумм превышений среднесуточных температур (“градусо-дней”) над базовой температурой в 10°C по данным спутниковой метеосъемки за 1983–2005 гг. (NASA…, 2018). На основе этих же данных вычислены индексы континентальности Конрада (Tukhanen, 1980) (табл. 1).
Таблица 2.
Результаты анализа таксономических и типологических спектров ценофлор кровохлебковых лугов Table 2. Results of the analysis of taxonomical and typological spectra of great burnet meadows in different geographical points
| Семейства, группы жизненных форм, гео- и экоэлементы/ Families, groups of life forms, plant range and ecological types | Поймы рек/Floodplains of rivers | ||||
|---|---|---|---|---|---|
| 1 | 2 | 3 | 4 | 5 | |
| I. Ранги ведущих семейств в семейственно-видовых спектрах/Ranks of leading families in family-species spectra | |||||
| Poaceae | 1 | 1 | 1 | 4 | 1 |
| Cyperaceae | 2 | 2–6 | <10 | 1–3 | 5–6 |
| Ranunculaceae | 5–7 | 2–6 | 6–7 | <10 | 3–4 |
| Rosaceae | 5–7 | 2–6 | 3–5 | 1–3 | 2 |
| Fabaceae | 5–7 | 2–6 | 3–5 | 1–3 | 7–9 |
| Apiaceae | <10 | 9–10 | 3–5 | <10 | <10 |
| Scrophulariaceae | – | 9–10 | <10 | <10 | 3–4 |
| Asteraceae | 3 | 2–6 | 2 | 5–9 | 5–6 |
| II. Амплитуда значений коэффициентов флористической общности Жаккара (%) (Parinova
et al., 2019) для:/ Amplitude of Jacquard’s similarity index values (%) (Parinova et al., 2019) for: |
|||||
| – видов/species | 8–15 | 8–20 | 8–11 | 9–11 | 3–8 |
| – родов/genera | 17–27 | 17–38 | 17–27 | 17–25 | 17–18 |
| – семейств/families | 43–50 | 39–50 | 50–52 | 47–57 | 40–52 |
| III. Группы жизненных форм, %/Groups of life-forms, % | |||||
| Кустарники, кустарнички и полукустарники / Shrubs, dwarfshrubs and semishrubs | 0 | 1 | 0 | 2 | 4 |
| Травянистые поликарпики/Polycarp herbs: | |||||
| – стержнекорневые/tap-rooted | 21 | 19 | 22 | 23 | 18 |
| – длиннокорневищно-стержнекорневые/tap-rooted with long rhizomes | 1 | 5 | 18 | 11 | 4 |
| – длиннокорневищные/long-rhizomatous | 13 | 49 | 30 | 38 | 26 |
| – короткокорневищные (двудольные)/short-rhizomatous (dicots) | 24 | 15 | 18 | 12 | 32 |
| – рыхлодерновинные (однодольные)/loosely bunched (monocots) | 26 | 3 | 7 | 1 | 8 |
| – плотнодерновинные (однодольные)/firm-bunched (monocots) | 3 | 0 | 0 | 0 | 0 |
| – прочие поликарпики/other polycarps | 3 | 5 | 3 | 8 | 1 |
| Монокарпики/Monocarps | 1 | 2 | 2 | 2 | 6 |
| IV. Широтные географические элементы, %/Types of latitudinal ranges, % | |||||
| Арктоальпийские и гипоарктомонтанные/Arctic-alpine and hypoarctic-montane | 0 | 0 | 0 | 0 | 4 |
| Бореальные/Boreal | 1 | 1 | 3 | 2 | 6 |
| Бореонеморальные/Boreal-nemoral | 26 | 18 | 26 | 14 | 14 |
| Неморальные/Nemoral | 18 | 2 | 0 | 2 | 4 |
| Лесостепные и степные/Forest-steppe and steppe | 2 | 28 | 1 | 11 | 10 |
| Полизональные/Multizonal | 53 | 51 | 70 | 71 | 42 |
| V. Хориономические географические элементы, %/Types of chorionomic ranges, % | |||||
| Голарктические/Holarctic | 7 | 8 | 21 | 13 | 16 |
| Восточноамерикано-евразиатские и амфиатлантические/East American-Eurasian and Amphiatlantic | 1 | 0 | 1 | 0 | 0 |
| Евразиатские и евразиатско-западноамериканские/Eurasian and Eurasian-West American | 33 | 48 | 51 | 55 | 28 |
| Евросибирские/Eurosiberian | 3 | 3 | 5 | 8 | 11 |
| Евросибирско-, европейско-, сибирско-древнесредиземноморские/ Eurosiberian-, European- and Siberian-Ancient Mediterranean | 50 | 39 | 22 | 19 | 9 |
| Европейские/European | 6 | 2 | 0 | 5 | 0 |
| Азиатские и сибирские/Asian and Siberian | 0 | 0 | 0 | 0 | 30 |
| Сибирско- и азиатско-американские/Siberian- and Asian-American | 0 | 0 | 0 | 0 | 6 |
| VI. Экологические элементы по отношению к влажности почвы, %/Ecological types in regard to soil moisture, % | |||||
| Ксеромезофиты/Xeromesophytes | 1 | 1 | 1 | 3 | 2 |
| Мезофиты/Mesophytes | 41 | 47 | 67 | 54 | 67 |
| Гигромезофиты/Hygromesophytes | 40 | 32 | 30 | 34 | 18 |
| Мезогигрофиты и гигрофиты/Mesohygrophytes and hygrophytes | 18 | 19 | 2 | 9 | 13 |
| VII. Экологические элементы по отношению к богатству почвы, %/Ecological types in regard to soil fertility, % | |||||
| Олигомезотрофы/Oligomesotrophic | 20 | 0 | 0 | 1 | 1 |
| Мезотрофы/Mesotrophic | 7 | 6 | 6 | 6 | 18 |
| Мезоэвтрофы/Mesoeutrophic | 59 | 55 | 87 | 79 | 78 |
| Эвтрофы и галофиты/Eutrophic and halophytic | 14 | 39 | 7 | 14 | 3 |
Примечание. Типологические спектры рассчитаны с учетом среднего проективного покрытия видов в конкретных географических пунктах. Процентные доли доминирующих гео- и экоэлементов выделены полужирным шрифтом. Прочерк “–” – отсутствие видов данного семейства. Прочие обозначения как в табл. 1.
Note. Calculations of typological spectra are based upon average projective cover values for species in each geographical point. Percentages of dominant range or ecological types are shown in bold. Dash “–” means the absence of species of a given family. For other notes see Table 1.
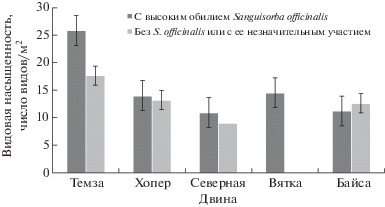
При сравнении видовой насыщенности на лугах с доминирующей и с малообильной или отсутствующей Sanguisorba officinalis (рис. 1) к первым отнесены площадки с покрытием вида более 10%. Использован t-критерий Стьюдента для неравночисленных выборок малого объема (Schmidt, 1984; Dörffel, 1990).
Рис. 1.
Видовая насыщенность на участках с доминирующей и с малочисленной (или отсутствующей) Sanguisorba officinalis в обследованных пойменно-луговых массивах
Fig. 1. Plant species numbers per 1 m2 in studied floodplain meadow plots with dominant and few (or lacking) Sanguisorba officinalis.
Темза / Thames;
Хопер / Khopyor;
Северная Двина / Northern Dvina;
Вятка / Vyatka;
Байса / Baisa.
Продуктивность травостоев оценена по сырому и сухому весу укоса в целом и по отдельным агроботаническим группам в период максимального развития травостоя (Klaap, 1961; Lepkovich, 2005). Отдельно взвешивался укос S. officinalis (табл. 3). Для каждого из географических пунктов по данным описаний рассчитаны параметрические коэффициенты корреляции между видовой насыщенностью и продуктивностью лугов (общей и по агроботаническим группам); результаты визуализированы с помощью программного пакета Vegan (Oksanen et al., 2017). Отдельно проанализирована многолетняя (1979–1986 гг.) динамика продуктивности кровохлебковых лугов в пойме р. Хопер порознь для участков с сенокошением и без него (280 площадок по 1 м2 за период наблюдений; данные Е.С. Нескрябиной).
Таблица 3.
Продуктивность травостоя на изученных лугах по отдельным агроботаническим группам (сухой вес, г/м2) Table 3. Herbage productivity of different agrobotanical groups (dry weight, g/m2) in studied meadows
| Агроботанические группы Agrobotanical groups |
Поймы рек/Floodplains of rivers | ||||
|---|---|---|---|---|---|
| Темза Thames |
Хопер Khopyor |
Северная Двина Northern Dvina |
Вятка Vyatka |
Байса Baisa |
|
| Злаки/Grasses | 198.4 | 114.2 | 72.1 | 151.4 | 36.3 |
| Осоки/Sedges | 35 | 73.2 | 11.1 | 65.7 | 71.2 |
| Ситники/Rushes | 107.6 | 0 | 0 | 0 | 0 |
| Хвощи/Horsetails | 0 | 0 | 9.9 | 6.4 | 7.2 |
| Бобовые/Legumes | 0 | 38.8 | 36.3 | 42.6 | 54.2 |
| Sanguisorba officinalis | 43.7 | 403.0 | 212.8 | 169.3 | 20.6 |
| Прочее разнотравье/Other forbs | 64.4 | 233.6 | 197.9 | 373.5 | 159.7 |
| Всего/Total | 449.1 | 862.8 | 540.1 | 808.9 | 349.2 |
Номенклатура сосудистых растений дана по С.К. Черепанову (Czerepanov, 1995).
ЦЕНОТИЧЕСКИЕ ОСОБЕННОСТИ КРОВОХЛЕБКОВЫХ ЛУГОВ В ПОЙМАХ РАЗНЫХ РЕК
Пойменные луга – классический пример азональной растительности. Они не выходят на плакоры и не формируют собственной зоны, но широко распространены в долинах рек таежной и широколиственнолесной зон, отчасти также в степной и тундровой зонах и в соответствующих высотных поясах горных систем (Walter, 1968; Tolmachev, 1974; Kamelin, 2018). Пойменные луга отличаются от водораздельных переменностью экологических режимов, пространственной неоднородностью растительного покрова и повышенным видовым разнообразием (Parinova et al., 2013, 2019; Rothero et al., 2016).
Все рассматриваемые массивы лугов с доминированием S. officinalis приурочены к центральной пойме, хотя, вероятно, и к различным ее уровням (либо сама центральная пойма различается по рельефу и относительной высоте в бассейнах разных рек). Для них характерно господство мезо- и гигромезофильного двудольного крупнотравья, что позволяет отнести эти луга к влажноразнотравной группе ассоциаций (Vasilevich, 2006). Ниже рассмотрим особенности таких лугов в конкретных географических пунктах, названных по соответствующим рекам (табл. 1).
Темза. Следуя английской Национальной классификации растительности (National vegetation classification) (Rodwell, 1992), кровохлебковые луга относятся к ассоциации Alopecurus pratensis–Sanguisorba officinalis (см. выше). Исходя из экологических условий произрастания сообществ, последняя должна входить скорее в лисохвостовую группу ассоциаций (Vasilevich, Bibikova, 2007). Однако в анализируемой выборке Alopecurus pratensis практически отсутствует, хотя представлены иные виды злаков, диагностических для ассоциации (Cynosurus cristatus, Festuca rubra, Holcus lanatus, Lolium perenne), и в целом сравнительно с другими географическими пунктами доля злаков в сложении травостоя здесь максимальна. В число доминантов, помимо Sanguisorba officinalis и Succisa pratensis, входят Agrostis stolonifera и кальцефильный мезогигрофит Carex flacca. К субдоминантам относятся Festuca rubra, Poa trivialis, Filipendula ulmaria s.l., изредка также Juncus acutiflorus. Исходя из набора и обилия видов, описанные луга тяготеют к субассоциации Agrostis stolonifera, сообщества которой наиболее толерантны к переувлажнению почв (Floodplain…, 2014). Но если судить по отдельным описаниям, эти луга могут быть отнесены, как минимум, к трем субассоциациям, что затрудняет более детальную характеристику всего лугового массива. В целом же список доминантов соответствует мезоэвтрофным лугам среднего и средневысокого уровня центральной поймы. Вхождение в число доминантов и субдоминантов соответственно Agrostis stolonifera и Festuca rubra отражает секторальную специфику ценотической приуроченности этих видов. Для Agrostis stolonifera характерна сезонная экологическая ниша: ее покрытие может варьировать от 1 до 70% в зависимости от сезона и погодных условий, вплоть до формирования сплавин во время продолжительных осенних паводков. Сезонная дифференциация экологических ниш в целом характерна для английских лугов, учитывая круглогодичный характер их вегетации (Grime, 1979). Вместе с тем A. stolonifera произрастает и на слабозадернованных отмелях, а также по берегам морей, что наблюдается и восточнее, тогда как первая из ниш перестает быть типичной. Произрастание (и даже доминирование) Festuca rubra на гигромезофильных лугах нередко не только в Западной Европе, но и в западной части Европейской России, в частности, на севере Смоленской и в центральной части Ленинградской областей.
Хопер. Здесь также представлены луга центральной поймы, однако в сложении их травостоя бóльшую роль играет разнотравье (Beguchev et al., 1930), в том числе мезогигрофильное (Euphorbia palustris). В числе сопутствующих мезогигрофитов и гигрофитов следует отметить виды Cyperaceae – Eleocharis palustris и Carex vulpina. В то же время в списке географических детерминантов представлены и мезофиты (Galium physocarpum, Inula britannica и др.). Среди них особое значение имеет Carex praecox, формирующая густой нижний подъярус под пологом более высоких трав. Alopecurus pratensis и Filipendula ulmaria s.l. играют роль субдоминантов, сопровождающих S. officinalis в верхнем подъярусе травостоя. Сочетание мезофитов с мезогигрофитами свидетельствует о пестроте микроусловий, что, впрочем, характерно для большинства типов пойменных лугов.
Северная Двина. Это самый северный из изученных луговых массивов, расположенный всего в 255 км южнее Полярного круга. Обследованные луга располагаются в центральной части островной и материковой поймы низовий Северной Двины. Они кратковременно и однократно заливаются весной при вскрытии льда (Parinova et al., 2013). Это типичные влажноразнотравные луга, где совместно доминируют гигромезофильные (Sanguisorba officinalis, Filipendula ulmaria s.l.) и мезофильные (Thalictrum simplex, Vicia cracca, Geranium pratense, Galium boreale) виды двудольных. Субдоминанты представлены Elytrigia repens и Equisetum arvense, что указывает на регулярное появление участков свежих аллювиальных наносов. Среди сопутствующих видов наиболее значимы Alopecurus pratensis и Lathyrus pratensis. Как обилие крупнотравья, так и высокий процент покрытия L. pratensis и (особенно) Vicia cracca придает северодвинским лугам урало-сибирские флороценогенетические черты (Kamelin, 2018). В целом сообщества флористически близки к вятским, однако, в отличие от них, формируются по более выровненной либо местами пониженной (судя по обилию Carex cespitosa) поверхности.
Вятка. Луга достаточно влажные, крупнотравные, с большой биомассой, занимают ровные плоские гривы в центральной пойме среднего уровня, реже неглубокие плоские понижения. Совместно с Sanguisorba officinalis доминируют мезофиты Galium boreale и Inula salicina. Сочетание гигромезо- и мезогигрофитов (Filipendula ulmaria s.l., Veronica longifolia) с мезофитами (Bromopsis inermis, Carex praecox) характерно и для субдоминантов. Alopecurus pratensis в списке сводного описания выглядит сопутствующим видом, однако в природе местами достигает значимого обилия. Carex praecox растет в смеси с Alopecurus pratensis и Bromopsis inermis, как правило, в понижениях между облесенными гривами. При доминантно-детерминантной классификации вятские луга в основном отнесены к ассоциации Galio-Alopecuretum Hundt 1958 var. Sanguisorba officinalis (Shchukina, 2012). Однако с позиций эколого-фитоценотической классификации они неоднородны: лишь 6 площадок из 10 можно отнести к лисохвостовым лугам, еще две – к костровым (с господством Bromopsis inermis), по одной – к девясиловым (с господством Inula salicina) и таволговым.
Байса. Влажноразнотравные луга с сочетанием мезофитов (Geranium wlassovianum, Vicia cracca, Hedysarum alpinum, Trollius vicarius) и мезогигрофитов (Equisetum palustre) в числе доминантов, по-видимому, наиболее сырые из всех. Sanguisorba officinalis переходит на роль субдоминанта. Местами присутствуют низкие кустарники (Salix rosmarinifolia, Spiraea salicifolia и др.). Характерно выраженное варьирование как по набору доминантов, так и по флористическому составу в целом, в первую очередь в зависимости от локальных особенностей режима увлажнения (Beliko-vich, 2015). Существенное влияние на набор видов оказывает и наличие многолетней мерзлоты, одновременно охлаждающей почву и обеспечивающей ее высокое влагосодержание. К концу лета на пойменных лугах мерзлота протаивает лишь на глубину 125–175 см (Berg, 1947). Невысокие (сравнительно с другими участками) значения видовых покрытий обусловлены экстремальными весенне-летними засухами на протяжении 15 лет, включая и год описания. На момент последнего многие виды еще не успели полностью отрасти.
СРАВНИТЕЛЬНЫЙ АНАЛИЗ ПОЙМЕННО-ЛУГОВЫХ ЦЕНОФЛОР
Различные методы анализа ценофлор обследованных лугов выявляют сходные черты их состава и структуры на разных широтах и в разных долготных секторах, что обусловлено азональным характером формирования лугового покрова пойм. Также прослеживаются специфические особенности ценофлор, зависящие от географического положения речных бассейнов и от степени их взаимной удаленности.
Видовое богатство и видовая насыщенность. Самая богатая видами ценофлора свойственна лугам в пойме Байсы. Это единственная автохтонная ценофлора из числа изученных, поскольку территория юга Сибири не подвергалась покровному оледенению в плейстоцене (Khotinskii, 1977); все остальные ценофлоры – миграционные, хотя и различающиеся по времени генезиса (Tolmachev, 1974; Kamelin, 2018). Однако видовая насыщенность лугов по Байсе довольно низка и лишь ненамного превосходит таковую на лугах по Северной Двине, для которых характерен и самый низкий уровень видового богатства. Луга в пойме Северной Двины – самые северные и наименее теплообеспеченные из числа изученных; вдобавок их ценофлора самая молодая (Yelina et al., 2000; Schmidt, 2005). Низкий уровень видовой насыщенности на лугах по Байсе, вероятно, является следствием континентальности восточносибирского климата с его морозными зимами и выраженным суточным ходом температур.
Вторыми по богатству ценофлоры и первыми по видовой насыщенности оказались луга в пойме Темзы, развитые в океаническом теплоумеренном климате без выраженных колебаний температуры и влажности. Максимальная видовая насыщенность кровохлебковых пойменных лугов в Великобритании составляет 43 вида, средняя – 16–25 (Rothero et al., 2016), тогда как для европейских лугов класса Molinio-Arrhenatheretea R.Tx. 1937, к которому относятся и исследуемые луга, последний показатель варьирует от 4 до 28 видов на 1 м2 (Dengler et al., 2016).
На лугах в поймах Хопра и Вятки наблюдаются промежуточные значения показателей α-разнообразия. По богатству ценофлоры луга по Хопру ближе к таковым на Темзе, по Вятке – ближе к северодвинским. Это соответствует зональному положению луговых массивов: первая пара расположена в пределах широколиственнолесной (в широком смысле, включая и лесостепь), вторая – таежной зоны. Известно, что азональным сообществам до известной степени тоже свойственны зональные отличия (Lipatova, 1980; Rebristaya, 2013; Kucherov, 2016 a). При этом видовая насыщенность травостоя на лугах по Хопру и Вятке примерно одинакова (табл. 1).
На некоторых из европейских лугов с господством Sanguisorba officinalis (Темза, Хопер; рис. 1) видовая насыщенность оказалась несколько выше сравнительно с лугами с малым обилием этого вида. Статистически, однако, это подтверждено лишь на лугах по Темзе, притом только в качестве тенденции (на уровне значимости α ≤ 10%; табл. 1) (Dörffel, 1990). Тем не менее, полученные цифры свидетельствуют о S. officinalis как о сравнительно мощном (по меркам луговых сообществ) эдификаторе, создающем благоприятные условия для произрастания сопутствующих ему видов. Противоположная тенденция отмечена на лугах по Байсе (рис. 1), где кровохлебка никогда не входит в число основных доминантов. Однако там различия недостоверны из-за недостаточного числа описаний.
Вероятная причина того, что доминирование S. officinalis благоприятствует видовой насыщенности лугов по Темзе, кроется в глубокой стержневой корневой системе этого вида. Корневые окончания кровохлебки и сопровождающих ее корневищных и дерновинных трав хотя бы отчасти приурочены к разным горизонтам почвенного профиля, т.е. наблюдаются комплементарные стратегии водного и минерального питания, а это способствует сосуществованию различных видов. Аналогичные явления подземной ярусности всасывающих корней отмечены в криофитных степях (Polozova, 1983) и дриадовых горных тундрах (Kucherov, Zagidullina, 2001) Чукотки, а также в прериях и луговых степях (Walter, 1968). В то же время на таволговых лугах (Lysimachio vulgaris–Filipenduletum Bal.-Tul. 1978), соседствующих с кровохлебковыми в поймах рек Европейской России, корневые окончания всех видов, включая и короткокорневищную Filipendula ulmaria s.l., приурочены к одному и тому же верхнему горизонту профиля, что ведет к усилению напряженности конкуренции. Низкая видовая насыщенность травостоя на таволговых лугах очевидна при их описании (Cherednichenko, Borodulina, 2017; наблюдения И.Б. Кучерова 1996–2017 гг.).
Таксономический анализ ценофлор. По числу видов и родов в большинстве ценофлор преобладает семейство (сем.) Poaceae, что закономерно для Голарктики, как и высокий ранг сем. Asteraceae в первых трех ценофлорах из пяти. Первая триада семейств ценофлоры лугов на Темзе тоже наиболее типична для спектров флор по всей Голарктике (Tolmachev, 1974). Второе место сем. Cyperaceae в семейственно-видовых спектрах ценофлор Темзы, Хопра и Вятки характерно для арктобореальной области Голарктики. Высокие ранги семейств Rosaceae и Fabaceae отражают соответственно европейские (неморально-субатлантические) и древнесредиземноморские черты формирования ценофлор Хопра, Северной Двины и Вятки, Ranunculaceae и Scrophulariaceae – высокогорные черты генезиса ценофлоры Байсы (Khokhryakov, 2000). Сказанное, однако, вряд ли относится ко 2-му месту сем. Rosaceae в спектре ценофлоры Байсы (табл. 2), которое следует объяснять иными причинами.
В целом следует согласиться с тем, что анализ таксономических спектров флор и ценофлор не слишком и не всегда информативен (Kamelin, 2018).
При сравнении коэффициентов флористической общности Жаккара на уровне видов закономерно выделяются низкие значения последних для лугов по Байсе, наиболее флористически специфичных сравнительно с европейскими луговыми массивами. Самые низкие значения коэффициентов (3%) отмечены в парах Темза–Байса и Хопер–Байса. Однако на уровне родов и семейств значения коэффициентов существенно выше: от 39 до 57% семейств одинаковы в каждой паре лугов (табл. 2).
Биоморфный анализ ценофлор. В спектрах биоморф всех исследованных луговых массивов (табл. 2) лидируют длиннокорневищные поликарпические травы с их развитой способностью к вегетативному расселению. Преобладание этой биоморфы в разных географических пунктах обусловлено доминированием различных видов. На лугах по Темзе это Carex flacca, по Хопру – Euphorbia palustris и Alopecurus pratensis, по Северной Двине – Thalictrum simplex, Elytrigia repens, Equisetum arvense, по Вятке – Inula salicina и Veronica longifolia, по Байсе – Equisetum palustre. На лугах по Хопру и Вятке к этому списку добавляются Bromopsis inermis и Carex praecox, по Хопру и Двине – Lathyrus pratensis, по Двине и Байсе – Vicia cracca.
Вторую и третью позиции в спектрах разделяют стержнекорневые и короткокорневищные двудольные травы. Позиции стержнекорневых видов во всех европейских ценофлорах обусловлены доминированием Sanguisorba officinalis, в ценофлоре Байсы – согосподством S. officinalis и Hedysarum alpinum вместе с совокупным эффектом произрастания многих сибирских видов, характерных для континентальных климатических условий и порознь не обильных. Во всех случаях значимая роль стержнекорневых видов обусловлена переменностью режима увлажнения. На лугах по Северной Двине и Вятке к списку стержнекорневых доминантов присоединяется длиннокорневищно-стержнекорневой Galium boreale.
К доминантам из числа короткокорневищных двудольных во всех европейских ценофлорах относится Filipendula ulmaria s.l., на Темзе – также Succisa pratensis, на Двине – Geranium pratense, на Байсе – G. wlassovianum и Trollius vicarius.
Только на лугах по Темзе существенную роль в сложении спектра играют рыхлодерновинные (Festuca rubra, Poa trivialis, Juncus acutiflorus) и столонообразующие (Agrostis stolonifera) травы; отмечены и плотнодерновинные злаки (Cynosurus cristatus, Deschampsia cespitosa), в целом нетипичные для средней поймы.
Соотношение широтных геоэлементов. Во всех ценофлорах наибольшая доля в спектрах широтных геоэлементов приходится на полизональные виды. Это обусловлено как азональным характером пойменно-луговой растительности (Walter, 1968; Tolmachev, 1974; Kamelin, 2018), так и широким зональным простиранием ареалов многих видов, формирующих луговые сообщества (Hultén, Fries, 1986), и столь же характерно для суходольных лугов (Kucherov, 2016 a). Полизональными являются Festuca rubra s. l. (аркто-неморальная), Bromopsis inermis, Thalictrum simplex, Lathyrus pratensis, Vicia cracca, Inula salicina (бореально-лесостепные), Equisetum palustre (бореонеморально-степной). Galium boreale и сама Sanguisorba officinalis относятся к еще более широко распространенным видам гипоаркто-степного субэлемента полизонального элемента (Kucherov, 2016 b; Parinova et al., 2019). К полизональным принадлежат все константные виды кровохлебковых лугов (S. officinalis, Lathyrus pratensis, Vicia cracca). На лугах по Темзе к полизональным доминантам относятся также Agrostis stolonifera, Festuca rubra, Juncus acutiflorus, к сопутствующим видам – Arrhenatherum elatius, Deschampsia cespitosa, Carex panicea, Lotus corniculatus s.l., Trifolium pratense, Plantago lanceolata. На Северной Двине региональные полизональные доминанты представлены Elytrigia repens, Thalictrum simplex, Equisetum arvense, на Вятке – Bromopsis inermis, Inula salicina, Veronica longifolia, в обоих луговых массивах – Galium boreale. Для этих лугов, находящихся в пределах бореально-лесной зоны Европы, характерна самая высокая доля участия полизональных видов в формировании спектра. На лугах по Байсе из видов данного элемента наиболее значимы Equisetum palustre и Trollius vicarius (табл. 1, 2).
В трех ценофлорах из пяти (Темза, Двина, Вятка) на 2-е место в спектре выходят бореонеморальные виды, в спектрах лугов по Хопру и Байсе занимающие лишь третью позицию. В числе бореонеморальных доминантов следует упомянуть Filipendula ulmaria s.l. для всех европейских ценофлор, Succisa pratensis и Poa trivialis для лугов по Темзе, Alopecurus pratensis – по Хопру, Двине и Вятке, Galium physocarpum – по Хопру, Geranium pratense – по Двине, Rumex acetosa – по Вятке. На лугах по Байсе к доминантам данного элемента относится Geranium wlassovianum (Parinova et al., 2019).
В спектрах ценофлор Хопра (2-е место), Вятки и Байсы (3-е место) важную роль играют также лесостепные виды, использующие поймы как миграционные коридоры для продвижения на север благодаря наличию грив и иных дренированных и прогреваемых экотопов на фоне общей выравненности пойменно-лугового микроклимата. Как на Хопре, так и на Вятке список лесостепных доминантов возглавляет Carex praecox. На расположенных собственно в пределах лесостепной полосы лугах по Хопру к ней добавляется Euphorbia palustris, а из числа спутников – Symphytum tauricum и Gratiola officinalis. На Вятке к значимым сопутствующим видам данного элемента относятся Cenolophium denudatum, Allium angulosum, Iris sibirica, Fragaria viridis. На Байсе лесостепными субдоминантами являются Seseli condensatum и Galium verum s.l. На Темзе и Северной Двине лесостепные виды почти не представлены в силу соответственно океаничности либо недостаточной теплообеспеченности климата; исключением в первом случае служит Silaum silaus.
Бореальные виды выходят на 2-е место лишь в спектре ценофлоры Байсы вследствие как континентализации климата, так и наличия многолетней мерзлоты (см. выше). Высокий ранг обусловлен обилием доминирующего Hedysarum alpinum, отчасти также Artemisia integrifolia, Veratrum lobelianum и других сопутствующих видов. Неморальные виды достигают 3-го места только в спектре ценофлоры Темзы (что опять-таки соответствует зональному положению последней) благодаря обилию Carex flacca и большому числу сопутствующих видов: Centaurea nigra, Holcus lanatus, Cynosurus cristatus, Primula veris и др. (табл. 1, 2).
Соотношение хориономических геоэлементов. Во всех трех ценофлорах Русской равнины (Хопер, Двина, Вятка) первое место в спектрах занимают евразиатские (вкупе с евразиатско-западноамериканскими) виды, что указывает на общность генезиса этих ценофлор в голоцене (Tolmachev, 1974; Schmidt, 2005; Kamelin, 2018). Помимо Sanguisorba officinalis это Alopecurus pratensis, Lathyrus pratensis, Vicia cracca, по Двине – также Thalictrum simplex, Geranium pratense, Carex cespitosa, по Хопру и Вятке – Carex praecox, только по Вятке – Inula salicina, Veronica longifolia, Rumex acetosa. В ценофлорах Темзы и Байсы доля евразиатских видов снижается до 2-й позиции в спектрах. На лугах по Темзе из них наиболее значимы Agrostis stolonifera, Poa trivialis, Deschampsia cespitosa, по Байсе – Seseli condensatum и Veratrum lobelianum.
Второе место в спектрах ценофлор Русской равнины принадлежит сравнительно теплолюбивым евросибирско- и европейско-древнесредиземноморским видам, хотя бы отчасти вошедшим в состав ценофлор в период атлантического оптимума голоцена (Khotinskii, 1977; Yelina et al., 2000). Список этих видов возглавляет Filipendula ulmaria s.l. На лугах по Хопру обильны также Euphorbia palustris, Kadenia dubia, Glechoma hederacea, Galium physocarpum, по Хопру и Вятке – Bromopsis inermis, по Двине – Elytrigia repens и Phleum pratense. В пойме Темзы обсуждаемые виды выходят на первое место в спектре за счет обилия Succisa pratensis, Carex flacca, а также большого числа сопутствующих видов – Arrhenatherum elatius, Carex panicea, Lotus corniculatus s.l., Silaum silaus, Hordeum secalinum, Briza media, Plantago lanceolata. В сложении континентальной ценофлоры Байсы роль подобных видов невелика; упоминания заслуживает лишь сибирско-древнесредиземноморская Artemisia integrifolia. Вместо этого на первое место в спектре этой автохтонной флоры выходят азиатские виды: Geranium wlassovianum, Trollius vicarius, из сопутствующих – Rumex gmelinii, Thalictrum contortum, Carex enervis, Filipendula palmata и др. Благодаря обилию Hedysarum alpinum на 4-е место в спектре поднимаются евросибирские виды, малозначимые при формировании спектров европейских ценофлор.
Третью строчку в спектрах всех ценофлор таежной зоны (Двина, Вятка, Байса) занимают голарктические (циркумбореальные) виды. Это Galium boreale, а также виды Equisetum – E. arvense на Двине и E. palustre на Байсе. В формировании спектра последней из ценофлор также важна совокупная роль сопутствующих голарктических видов: Calamagrostis langsdorffii, Poa pratensis, Galium verum s.l.
В более южных ценофлорах роль голарктических видов невелика несмотря на то, что Festuca rubra входит в число доминантов лугов в пойме Темзы.
Роль европейских видов заметна только на лугах по Темзе (Centaurea nigra, Cynosurus cristatus, Holcus lanatus, Primula veris) и Вятке (Ptarmica vulgaris и Lysimachia nummullaria). То же касается и сибирско-американских видов в спектре Байсы (Helictotrichon hookeri, Carex vesicata, Elymus jacu-tensis и др.) (табл. 1, 2).
Экологический анализ ценофлор. В спектрах трофоэлементов всех пяти ценофлор выраженно преобладают мезоэвтрофные виды, что ожидаемо для пойменных лугов с их ежегодным отложением наилка. К этому трофоэлементу относятся как S. officinalis (см. выше), так и большинство региональных доминантов и субдоминантов кровохлебковых лугов. В ценофлоре Северной Двины на мезоэвтрофные виды приходится более 4/5 всего спектра. В большинстве остальных спектров возможно выделить второй по значимости трофоэлемент, варьирующий от региона к региону. В ценофлорах Хопра и Вятки это эвтрофные виды в широком их понимании: Carex praecox, Bromopsis inermis, из видов-спутников – Allium angulosum. На хоперских лугах в числе эвтрофов-доминантов характерны также Euphorbia palustris и Inula britannica, среди сопутствующих видов – олигогалинный Sium sisaroideum. При сравнении всех пяти ценофлор роль эвтрофных видов наиболее весома именно на лугах по Хопру. Это тоже закономерно, учитывая как карбонатность почв, так и их засоление в поймах рек по мере континентализации климата в южных широтах (Kamelin, 2018). Напротив, на лугах по Байсе за счет евросибирского бореального Hedysarum alpinum возрастает роль мезотрофов, что обусловлено преобладанием силикатных почвообразующих пород на фоне резко континентального восточносибирского климата.
В пойме Темзы в качестве сопровождающих трофоэлементов выступают как эвтрофы (Carex flacca, из видов-спутников – Lotus corniculatus, Bromus racemosus, олигогалинный Hordeum secalinum, мезогалинный Silaum silaus), так и характерные лишь для этих лугов олигомезотрофы (Succisa pratensis, из видов-спутников – Luzula campestris). Столь сложная структура спектра свидетельствует о пестроте микроусловий на этих лугах, включая сочетание задернелых кислых почв с богатыми аллювиями.
По отношению к влажности почвы в спектрах всех ценофлор преобладают мезофиты, на втором месте гигромезофиты. Для ценофлоры лугов по Байсе характерна наибольшая мезофитизация, что свойственно и растительности таежной зоны Сибири в целом (Tsatsenkin et al., 1978). Напротив, в пойме Темзы доли мезофитов и гигромезофитов почти выравниваются. Состав спектров лугов таежной зоны Европейской России (Двина, Вятка) практически исчерпывается этими двумя элементами, тогда как в спектрах остальных ценофлор к ним добавляются мезогигрофиты: Carex flacca по Темзе, Euphorbia palustris по Хопру, Equisetum palustre по Байсе (табл. 1, 2). Описания лугов в пойме Хопра в наибольшей степени неоднородны по условиям увлажнения. О принадлежности пойменно-луговых доминантов к тем или иным экоэлементам по отношению к влажности почв было сказано выше при характеристике отдельных луговых массивов.
ПРОДУКТИВНОСТЬ КРОВОХЛЕБКОВЫХ ЛУГОВ
Продуктивность надземной фитомассы кровохлебковых лугов различна в разных частях географического градиента (табл. 3). Наименьшая продуктивность (менее 350 г/м2) отмечена на лугах по Байсе, что лишь отчасти объяснимо исходя из континентальности и малой теплообеспеченности региона и наличия мерзлотных почв. Данные для этих лугов скорее выглядят потенциально заниженными вследствие экстремальных летних засух последних 15 лет. Для сравнения, продуктивность высоковейниковых (Calamagrostis langsdorffii) пойменных лугов Верхнечарской котловины (Северное Забайкалье) составляет 470–530 г/м2 (Garashchenko, 1993).
Кровохлебковые луга по Темзе и Северной Двине более продуктивны; при этом основной вклад в суммарную продукцию травостоя лугов в пойме Темзы вносят не виды разнотравья, а злаки и ситники. Отмеченные показатели продуктивности (табл. 3) соответствуют таковым пойменных лугов таежной и широколиственнолесной зон европейского субконтинента. В поймах рек таежной зоны Европейской России продуктивность лисохвостовых, костровых и пырейных лугов может превышать 300–400, а влажноразнотравных – достигает 500 г/м2 (Shennikov, 1941). В частности, в Холмогорском районе Архангельской обл. продуктивность чемеричных (Veratrum lobelianum) лугов равняется 388, пырейно-щучковых средней поймы – 432, красноовсяницевых прируслового вала Северной Двины – 454, а у осоково (Carex acuta)-лисохвостовых лугов превышает таковую кровохлебковых, достигая 754 г/м2 (Shennikov, Kondratieva, 1933). В пойме р. Оки (средняя полоса Европейской России) продуктивность костровых лугов составляет 554, двукисточниковых – 517, а влажноразнотравных (гераневых) – 475 г/м2 (Rabotnov, 1984).
Наибольшей продуктивностью (свыше 800 г/м2) отличаются кровохлебковые луга в поймах Хопра (за счет собственно Sanguisorba officinalis) и Вятки (за счет других видов разнотравья). Известно, что в годы с благоприятными условиями увлажнения продуктивность крупноразнотравно-лисохвостовых пойменных лугов лесостепи существенно превышает таковую лугов, произрастающих севернее (Shennikov, 1941). Тем не менее, возможно, что в данных условиях господство S. officinalis действительно способствует повышению количественных показателей продуктивности травостоя. Следует, однако, учесть и разброс значений продуктивности по годам (рис. 2), а также по микроэкотопам. Даже на высоковейниковых лугах бассейна Нижней Оби (подзона южных тундр) на наиболее благоприятных микроэкотопах продуктивность достигала 775 г/м2 при средней продуктивности всего лишь 214 г/м2 (Peshkova, 1987).
Рис. 2.
Многолетние (1979–1986 гг.) изменения биомассы травостоя кровохлебковых лугов (вверху) и Sanguisorba officinalis (внизу) в пойме р. Хопер
Fig. 2. Long-term (1979–1986) changes of total burnet meadow (top) and Sanguisorba officinalis (bottom) standing crop in the Khopyor River floodplain
Dashed and solid lines correspond to plots with and without mowing, respectively.
Общая биомасса / total weight.
Биомасса сухой вес, г/м2 / total dry weight, g/m2.
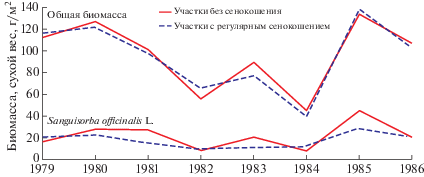
Регулярное сенокошение не способствует увеличению продуктивности Sanguisorba officinalis в абсолютном выражении в каждое из отдельно взятых лет, скорее повышая продуктивность видов, которые ей сопутствуют. Однако оно в какой-то мере выравнивает погодичные колебания биомассы S. officinalis, что оказывает нивелирующее влияние и на колебания общей биомассы травостоя, делая его продуктивность более устойчивой (рис. 2).
В отсутствие сенокошения либо пожаров на кровохлебковых лугах Западной Сибири покрытие Sanguisorba officinalis резко уменьшается в течение 15 лет (Tyurin, 2015). Уничтожая всходы деревьев и кустарников и запасы неразложенной подстилки, регулируемые весенние палы являются одним из факторов, поддерживающих существование пойменно-луговых сообществ как в Западной Сибири, так и на севере Европейской России. При этом, однако, очевидно негативное влияние этих палов на видовое разнообразие растений и животных.
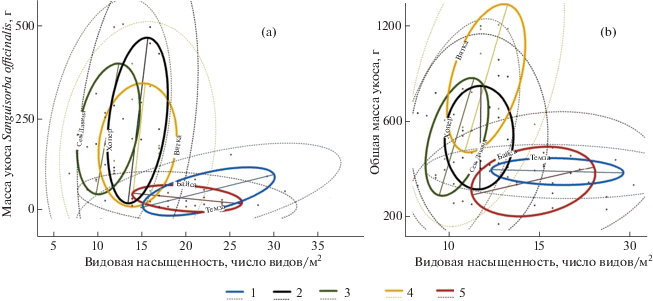
Примечательно, что на лугах по Хопру и Северной Двине не наблюдается положительных корреляций между биомассой Sanguisorba officinalis и видовой насыщенностью сообществ. Существенно меньшая биомасса S. officinalis на лугах по Темзе и Байсе также соответствует большей видовой насыщенности травостоев. Наиболее продуктивными оказались сообщества с промежуточными, а не крайними значениями видовой насыщенности (рис. 3). Эти факты можно было бы счесть проявлениями эффекта доминирования S. officinalis, если бы они не противоречили данным о росте видовой насыщенности лугов на Темзе с господством этого вида сравнительно с лугами с его малым обилием или без него (рис. 1; см. выше). Аналогичные закономерности характерны и для связи общей продуктивности травостоев с их видовой насыщенностью (рис. 3). Полученные данные требуют дальнейшего анализа и осмысления.
Рис. 3.
Видовая насыщенность сообществ не коррелирует положительно ни с биомассой Sanguisorba officinalis (а), ни с продуктивностью кровохлебковых лугов (b)
Поймы рек: 1 – Темза, 2 – Хопер, 3 – Северная Двина, 4 – Вятка, 5 – Байса.
Сплошная линия – стандартное отклонение, пунктирная – эллипс вероятностей (95%).
Fig. 3. Plant species number per unit area positively correlates neither with Sanguisorba officinalis biomass (a), nor with total herbage productivity in burnet meadows (b)
Floodplains of rivers: 1 – Thames, 2 – Khopyor, 3 – Northern Dvina, 4 – Vyatka, 5 – Baisa. Solid and pointed lines correspond to standard deviation and 95% probability ellipse, respectively.
Масса укоса Sanguisorba officinalis, г / biomass of Sanguisorba officinalis, g.
Общая масса укоса, г / total herbage productivity, g.
Видовая насыщенность, число видов/м2 / number of species/m2.
ВЫВОДЫ
1. Несмотря на азональную природу сообществ, состав и структура ценофлор пойменных кровохлебковых лугов демонстрируют зональные, а также секторальные черты, чему способствует протяженный ареал этих сообществ.
2. “Ядро” флороценотического комплекса кровохлебковых лугов формируют евразиатские полизональные мезоэвтрофные мезофиты и гигромезофиты.
3. Видовое богатство кровохлебковых лугов возрастает как в условиях автохтонного генезиса ценофлоры, так и в океаническом климате.
4. Доминирование Sanguisorba officinalis на лугах в пойме р. Темзы способствует увеличению видовой насыщенности травостоя этих лугов.
5. В пойме р. Хопер господство S. officinalis предположительно приводит к приросту продуктивности травостоя, что может оказаться характерным и для пойменных лугов лесостепи в целом.
Список литературы
[Atlas] Атлас ареалов и ресурсов лекарственных растений СССР. 1980. М. 340 с.
[Beguchev et al.] Бегучев П.П., Бычкова Е.Х., Николюкин А.И. 1930. Ботанико-агрономические исследования лугов р. Хопра в пределах Хоперского округа б. Сталинградской губернии. – Тр. Нижне-Волжск. о-ва краеведения. Саратов. Вып. 37. 48 с.
[Belikovich] Беликович А.В. 2017. Растительность Даурии [Электронный ресурс]. http://ukhtoma.ru/geobotany/dahuria_11.htm (доступ от 20.04.2020).
[Berg] Берг Л.С. 1947. Климат и жизнь. 2-е изд. М. 356 с.
[Cherednichenko, Borodulina] Чередниченко О.В., Бородулина В.П. 2017. Таволговые луга Полистовского заповедника. – Бюл. Брянск. отд. РБО. 3 (11): 63–68.
[Czerepanov] Черепанов С.К. 1995. Сосудистые растения России и сопредельных государств (в пределах бывшего СССР). СПб. 991 с.
[Chisla] Числа хромосом цветковых растений Флоры СССР: Семейства Moraceae–Zygophyllaceae. 1993. СПб. 430 с.
Council Directive 92/43/EEC of 21 May 1992 on the conservation of natural habitats and of wild fauna and flora. 1992. – Official Journal of European Communities. L 206: 7–50.
[Degteva, Dubrovskii] Дегтева С.В., Дубровский Ю.А. 2014. Лесная растительность бассейна р. Илыч в границах Печоро-Илычского заповедника. СПб. 291 с.
Dengler J., Biurrun I., Apostolova I., Baumann E., and 27 more participants. 2016. Scale-dependent plant diversity in Palaearctic grasslands: a comparative overview. – Bull. Eurasian Dry Grassland Group. 31: 12–36. https://doi.org/10.1127/phyto/2018/0267
Dörffel K. 1990. Statistik in der analytischen Chemie. Leipzig. 256 S.
Ellenberg H., Weber H.E., Düll R., Wirth W., Werner W., Paulißen D. 1992. Zeigerwerte von Pflanzen in Mitteleuropa. 2 Aufl. – Scripta Geobot. 18: 1–258.
[Ermakova] Ермакова И.М. 1983. Кровохлебка лекарственная (Sanguisorba officinalis). – В кн.: Диагнозы и ключи возрастных состояний луговых растений. Ч. 2. М. С. 47–51.
Floodplain Meadows Partnership. 2014. A revision of the Alopecurus pratensis–Sanguisorba officinalis (MG4) grassland community of the NVC 2014. http://www.floodplainmeadows.org.uk/sites/www.floodplainmeadows.org.uk/files/files/events/MG4subcomms%20brochure%20final.pdf (accessed on 21.04.2020).
[Garashchenko] Гаращенко А.В. 1993. Флора и растительность Верхнечарской котловины (Северное Забайкалье). Новосибирск. 280 с.
Grime J.P. 1979. Plant strategies and vegetation processes. London; New York. 222 p.
Hultén E., Fries M. 1986. Atlas of North European vascular plants, north of the Tropic of Cancer: In 3 t. Königstein. 1172 p.
IPCN Chromosome Reports. 2020. http://www.tropicos.org/Name/27800186?projectid=9 (accessed on 21.04.2020).
Jefferson R.J. 1997. Distribution, status and conservation of Alopecurus pratensis–Sanguisorba officinalis floodplain meadows in England: English Nature Research Report No. 249. www.jncc.gov.uk/page-3711 (accessed on 14.02.2018).
[Kamelin] Камелин Р.В. 2018. География растений. СПб. 306 с.
Khela S. 2013. Sanguisorba officinalis. – In: The IUCN Red List of threatened species 2013: e.T203458A2765918. http://www.iucnredlist.org/details/203458/1 (accessed on 13.02.2018).
[Khokhryakov] Хохряков А.П. 2000. Таксономические спектры и их роль в сравнительной флористике. – Бот. журн. 85 (5): 1–11.
[Klaap] Клаап Э. 1961. Сенокосы и пастбища. М. 613 с.
[Kucherov] Кучеров И.Б. 2016 a. Линейная зависимость состава и обилия видов растений от климатических факторов на суходольных лугах Севера и Северо-Запада Европейской России. – Фиторазнообразие Восточной Европы. 10 (2): 4–32.
[Kucherov] Кучеров И.Б. 2016 b. О подразделении типов ареалов полизональных и плюрирегиональных видов для целей сопряженного анализа флор сосудистых растений, мохообразных и лишайников. – Комаровские чтения (Владивосток). 64: 138–197.
[Kucherov] Кучеров И.Б. 2019. Ценотическое и экологическое разнообразие светлохвойных лесов средней и северной тайги Европейской России. СПб. 568 с.
[Kucherov et al.] Кучеров И.Б., Милевская С.Н., Полозова Т.Г. 2000. Структура флоры заповедника “Кивач” в контексте сравнения локальных флор на широтном профиле Восточной Фенноскандии. – В кн.: Сравнительная флористика на рубеже III тысячелетия: достижения, проблемы, перспективы. СПб. С. 63–83.
[Kucherov, Zagidullina] Кучеров И.Б., Загидуллина А.Т. 2001. Самовосстановление растительных сообществ: примеры, механизмы, подходы к описанию. – Журн. общей биологии. 62 (5): 410–424.
Kurtto A. 2009. Rosaceae. – In: The Euro+Med Plantbase – the information resource for Euro-Mediterranean plant diversity. http://ww2.bgbm.org/EuroPlusMed/PTaxonDetail.asp?NameCache=Rosaceae&PTRefFk=7300000 (accessed on 13.02.2018).
[Lepkovich] Лепкович И.П. 2005. Современное луговодство. СПб. 424 с.
Li Chaoluan, Ikeda H., Ohba H. 2003. Sanguisorba Linnaeus. – In: Flora of China. Vol. 9: Pittosporaceae through Connaraceae. Beijing; St.Louis. P. 384–387.
[Lipatova] Липатова В.В. 1980. Растительность пойм. – В кн.: Растительность европейской части СССР. Л. С. 346–372.
[Maevskii] Маевский П.Ф. 2006. Флора средней полосы европейской части России. 10-е изд. М. 600 с.
[Markov] Марков М.В. 1962. Общая геоботаника. М. 450 с.
NASA prediction of worldwide energy resources. 2018. https://power.larc.nasa.gov/data-access-viewer/ (accessed on 23.04.2020).
[Nomokonov] Номоконов Л.И. 1959. Пойменные луга Енисея. М. 478 с.
Nordborg G. 1963. Studies in Sanguisorba officinalis L. – Bot. Notis. 116 (2): 267–288.
Oksanen J., Blanchet F.G., Kindt R., Legendre P., O’Hara R.B., Simpson G.L., Solymos P., Stevens M.H.H., Wagner H. 2017. Vegan: Community ecology package. R package version 2.4-4. https://CRAN.R-project.org/package=vegan (downloaded on 13.02.2018).
[Parinova et al.] Паринова Т.А., Наквасина, Е.Н., Сидорова О.В. 2013. Луга островной поймы низовий Северной Двины. Архангельск. 146 с.
[Parinova et al.] Паринова Т.А., Татаренко И.В., Волков А.Г., Чередниченко О.В., Кучеров И.Б., Щукина К.В., Нескрябина Е.С., Пыжикова Е.М., Цыренова М.Г., Савиных Н.П., Пересторони-на О.Н, Шабалкина С.В. 2019. Флористические особенности пойменных лугов с Sanguisorba officinalis на географическом градиенте. – В сб.: Пойменные и дельтовые биоценозы Голарктики: биологическое многообразие, экология и эволюция. Астрахань. С. 127–131.
[Peshkova] Пешкова Н.В. 1987. Реальная и потенциальная продуктивность злаковых сообществ. Свердловск. 136 с.
Phipps J.B. 2014. Rosaceae. – In: Flora of North America north of Mexico. Vol. 9: Picramniaceae to Rosaceae. http://www.efloras.org/florataxon.aspx?flora_id=1&taxon_id=10776 (accessed on 14.02.2018).
Pojar J., MacKinnon A., Alaback P.B., and British Columbia Forest Service. 1994. Plants of coastal British Columbia, including Washington, Oregon and Alaska. Edmonton. 527 p.
[Polozova] Полозова Т.Г. 1983. Состав биоморф и некоторые особенности структуры реликтовых степных сообществ Западной Чукотки. – Бот. журн. 68 (11): 1503–1512.
Preston C.D., Pearman D.A., Dines T.D. 2002. New atlas of the British and Irish flora: An atlas of the vascular plants of Britain, Ireland, the Isle of Man and the Channel Islands. Oxford. 11+910 p.
[Rabotnov] Работнов Т.А. 1984. Луговедение. 2-е изд. М. 320 с.
[Ramensky et al.] Раменский Л.Г., Цаценкин И.А., Чижиков О.Н., Антипин Н.А. 1956. Экологическая оценка кормовых угодий по растительному покрову. М. 472 с.
[Rebristaya] Ребристая О.В. 2013. Флора полуострова Ямал: современное состояние и история формирования. СПб. 311 с.
Rodwell J.S. (ed.) 1992. British plant communities. Vol. 3: Grasslands and montane communities. Cambridge. X+540 p.
Rodwell J.S., Morgan V., Jefferson R.G., Moss D. 2007. The European context of British lowland grasslands: JNCC Report, No. 394. Peterborough. 148 p.
Rothero E., Lake S., Gowing D. 2016. Floodplain meadows – beauty and utility: A technical handbook. Milton Keynes. 104 p.
[Shchukina] Щукина. К.В. 2012. Классификация разнотравных лугов поймы реки Вятки. – Бот. журн. 97 (12): 1568–1578.
[Shennikov] Шенников А.П. 1941. Луговедение. Л. 509 с.
[Shennikov, Kondratieva] Шенников А.П., Кондратьева Е.А. 1933. Пастбища Холмогорского района. – Тр. БИН АН СССР. 3 (1): 99–178.
[Schmidt] Шмидт В.М. 1984. Математические методы в ботанике. Л. 288 с.
[Schmidt] Шмидт В.М. 2005. Флора Архангельской области. СПб. 346 с.
[Serebryakov] Серебряков И.Г. 1962. Экологическая морфология растений: жизненные формы покрытосеменных и хвойных. М. 378 с.
[Soczava] Сочава В.Б. 1927. Ботанический очерк лесов Полярного Урала от р. Нельки до р. Хулги. – Тр. Бот. музея АН СССР. 21: 1–78.
[Takhtajan] Тахтаджян А.Л. 1978. Флористические области Земли. Л. 248 с.
[Tolmachev] Толмачев А.И. 1974. Введение в географию растений. Л. 244 с.
[Tsatsenkin et al.] Цаценкин И.А., Савченко И.В., Дмитриева С.И. 1978. Методические указания по экологической оценке кормовых угодий тундровой и лесной зон Сибири и Дальнего Востока по растительному покрову. М. 302 с.
Tukhanen S. 1980. Climatic parameters and indices in plant geography. – Acta Phytogeogr. Suec. 67: 1–105.
[Tyurin] Тюрин В.Н. 2015. Результаты 16-летних наблюдений за кровохлебковым лугом (пойма Оби в окрестностях Сургута). – В сб.: Современные проблемы ботаники, микробиологии и природопользования в Западной Сибири и на сопредельных территориях. Сургут. С. 181–183.
[Khotinskii] Хотинский Н.А. 1977. Голоцен Северной Евразии. М. 200 с.
[Vasilevich] Василевич В.И. 2006. Влажные разнотравные луга северо-запада Европейской России. – Бот. журн. 91 (9): 1313–1328.
[Vasilevich, Bibikova] Василевич В.И., Бибикова Т.В. 2007. Щучковые и лисохвостные луга Северо-Запада Европейской России. – Бот. журн. 92 (1): 29–41.
Walter H. 1968. Die Vegetation der Erde in öko-physiologischer Betrachtung. Bd II: Die gemäßigten und arktischen Zonen. Jena. 1001 S.
Whittaker R.H. 1972. Evolution and measurement of species diversity. – Taxon. 21: 213–251.
[Yakubov] Якубов В.В. 1996. Род Кровохлебка – Sanguisorba L. – В кн.: Сосудистые растения советского Дальнего Востока. Т. 8. СПб. С. 227–230.
[Yelina et al.] Елина Г.А., Лукашев А.Д., Юрковская Т.К. 2000. Позднеледниковье и голоцен Восточной Фенноскандии (палеорастительность и палеогеография). Петрозаводск. 242 с.
[Yurtsev] Юрцев Б.А. 1981. Реликтовые степные комплексы Северо-Восточной Азии: Проблемы реконструкции криоксеротических ландшафтов Берингии. Новосибирск. 168 с.
[Yurtsev, Kamelin] Юрцев Б.А., Камелин Р.В. 1991. Основные понятия и термины флористики. Пермь. 80 с.
[Yuzepchuk] Юзепчук С.В. 1941. Sanguisorba L. – В кн.: Флора СССР. Т. 10: Rosaceae. М.; Л. С. 422–428.
Zelnik I., Carni A. 2013. Plant species diversity and composition of wet grasslands in relation to environmental factors. – Biodivers. Conserv. 22: 2179–2192.
[Zhukova] Жукова Л.А. 1997. Онтогенетический атлас лекарственных растений. Йошкар-Ола. 240 с.
[Zverev] Зверев А.А. 2007. Информационные технологии в исследованиях растительного покрова. Томск. 304 с.
Дополнительные материалы отсутствуют.
Инструменты
Ботанический журнал