

Ботанический журнал, 2020, T. 105, № 3, стр. 280-292
ОСОБЕННОСТИ ПРОИЗРАСТАНИЯ DACTYLORHIZA FUCHSII (ORCHIDACEAE) НА ТЕРРИТОРИИ ЗАКАЗНИКА “БЫЛИНА” (КИРОВСКАЯ ОБЛАСТЬ)
Н. В. Косолапова 1, 3, *, Т. Л. Егошина 2, 3, **, Е. А. Лугинина 2, 3
1 Федеральный научный центр кормопроизводства и агроэкологии им. В.Р. Вильямса
141055 Московская обл., г. Лобня, Научный городок, корпус 1, Россия
2 Всероссийский научно-исследовательский институт охотничьего хозяйства
и звероводства им. проф. Б.М. Житкова
610000 г. Киров, ул. Преображенская, 79, Россия
3 Вятская государственная сельскохозяйственная академия
610017 г. Киров, Октябрьский пр., 133, Россия
* E-mail: natalika.vasil@yandex.ru
** E-mail: etl@inbox.ru
Поступила в редакцию 18.11.2019
После доработки 13.01.2020
Принята к публикации 14.01.2020
Аннотация
В условиях среднетаежной подзоны Кировской области установлена приуроченность Dactylorhiza fuchsii (Druce) Soo к классам Vaccinio-Piceetea, Querco-Fagetea, Molinio-Arrhenatheretea. Описаны растительные сообщества, в которых произрастал D. fuchsii. Представлены данные об освоении видом экологического пространства в условиях региона исследования по шкалам Д.Н. Цыганова и Г. Элленберга. Показано, что D. fuchsii характеризуется широким потенциальным диапазоном по большинству рассматриваемых факторов, а по нескольким почвенным шкалам диапазон изученных местообитаний выходит за потенциально возможные границы. Установлено, что по отношению к комплексу всех экологических шкал Д.Н. Цыганова D. fuchsii относится к мезобионтным видам (It = 0.51). По совокупности климатических факторов D. fuchsii принадлежит к гемиэврибионтным видам (It = 0.61). По фактору освещенности-затенения отностится к эврибионтным видам (It = 0.67). По отношению к комплексу почвенных факторов D. fuchsii является гемистенобионтным видом (It = 0.40). Выявлена антропотолерантность растительных сообществ с D. fuchsii по показателю гемеробии. При оценке уязвимости вида было установлено, что во всех ценопопуляциях с D. fuchsii преобладают o (олиго-) и m (мезо-) гемеробные виды. В онтогенезе D. fuchsii были выделены онтогенетические состояния от ювенильных до сенильных. Возрастные спектры ценопопуляций D. fuchsii за годы наблюдений в большинстве случаев – нормальные неполночленные (правосторонние), в которых преобладают имматурные и генеративные особи.
Семейство Орхидные (Orchidaceae Juss.) является одним из наиболее многочисленных среди цветковых растений. Представители семейства Orchidaceae встречаются практически во всех пригодных для обитания растений областях земли и отличаются своеобразием биологии на разных континентах. Большинство представителей семейства занесено в Красные книги государственного и регионального значения и охраняются на заповедных территориях, так как являются наиболее чувствительными к воздействию техногенных факторов и изменению окружающей среды. В связи с этим актуальна разработка критериев определения устойчивости популяций орхидных, которые могут быть основаны на выявлении эколого-ценотической приуроченности, структуры и состояния ценопопуляций (ЦП) этих растений.
Пальцекорник Фукса (Dactylorhiza fuchsii (Druce) Soo) – представитель семейства Orchidaceae, имеет евро-азиатский тип ареала (Aver’yanov, 2000), занимает значительную территорию Европы и Азии. В России распространен в европейской части в большинстве северных и центральных областей, в Заволжье, на Урале, Алтае, в Западной Сибири, доходит до Байкала в Иркутской области и Бурятии (Gubanov et al., 2002; Vakhrameeva et al., 2014; Il’ina, 2019).
D. fuchsii – многолетнее травянистое поликарпическое растение с пальчатораздельным стеблекорневым тубероидом (Tatarenko, 1996). Гигромезофит. Геофит. D. fuchsii лугово-лесной вид, встречающийся в лесах различного состава – смешанных, мелколиственных, широколиственных и темнохвойных, предпочитая участки с разреженным древесным и травяно-кустарничковым ярусом. Обычно ЦП D. fuchsii произрастают на сухих и незаболоченных участках, но также встречаются и по краям болот (Vakhrameeva et al., 1991; Ovesnov, 1997). Обладая широкой экологической амплитудой, ЦП этого вида встречаются как на полном свету, так и в полутени; произрастает как на сухих, так и влажных почвах. Представители вида обитают на щелочных почвах, богатых основаниями, иногда на слабокислой, но никогда не произрастают на очень кислой почве (Vakhrameeva et al., 1994).
D. fuchsii включен в список Приложения № 2 к Красной книге Кировской области (2014), как вид, нуждающийся в постоянном контроле и наблюдении.
Диагностика экологических параметров местообитаний по произрастающим на них видам растений является распространенной процедурой фитоиндикации местообитаний (Zubkova, 2011). Каждый вид характеризуется диапазоном значений экологического фактора, при котором возможно его существование. Экологическую позицию вида оценивают при помощи различных экологических шкал (Ilʻina, Dorogova, 2012). Наиболее часто используемыми при обработке геоботанических и экологических исследований в Европейской части России являются отечественные шкалы Д.Н. Цыганова (Tsyganov, 1983) и европейские шкалы Г. Элленберга (Ellenberg, 1974).
Целью исследования является выявление предпочтений Dactylorhiza fuchsii по ведущим экологическим и фитоценотическим факторам для оценки параметров местообитаний данного вида в условиях средне-таежной зоны (Кировская обл., Государственный природный заказник (ГПЗ) “Былина”).
МАТЕРИАЛ И МЕТОДИКА
Исследования проведены в 2012, 2014, 2016–2018 гг. на территории ГПЗ “Былина” расположенной в северо-западной части Кировской обл., в подзоне среднетаежных лесов.
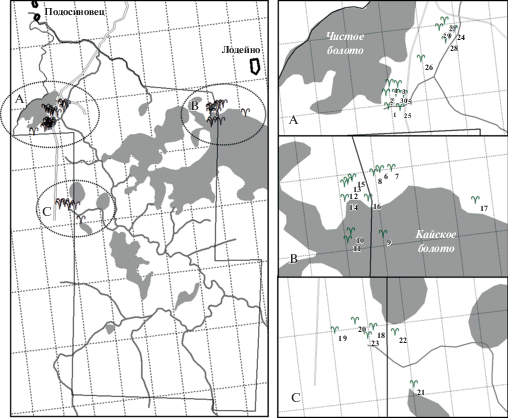
Исследовали 30 ценопопуляций (ЦП) D. fuchsii в разных частях заказника (рис. 1) с использованием общепринятых в популяционных исследованиях методов (Tsenopopuiyatsii…, 1988; Zlobin, 1989).
Рис. 1.
Месторасположение исследованных ценопопуляций Dactylorhiza fuchsii на территории ГПЗ “Былина”.
Соответствие обозначения на карте номеру ценопопуляции приведено в табл. 1.
Fig. 1. Location of studied Dactylorhiza fuchsii populations in “Bylina” State Nature Reserve.
Correspondence of marks on the map with population number is given in Table 1.
Растительные сообщества, в которых произрастал D. fuchsii, описывали согласно методикам эколого-фитоценотического подхода к классификации растительности (Polevaya…, 1964; Neshataev, 1987; Ipatov, 1998), с характеристикой видового состава и физико-географических условий. Таксационные характеристики приведены в соответствии с общепринятыми методами (Metody izucheniya…, 2002). Названия видов приведены в соответствии с базой данных The Plant List (http://www.theplantlist.org/).
За счетную единицу принимали побег. На каждой площадке подсчитывали число побегов и определяли их онтогенетическое состояние. В ходе исследований, были определены онтогенетические состояния для 2010 побегов. Поскольку D. fuchsii является редким видом, онтогенез определяли на основании надземной части растений с использованием методик, разработанных для редких видов орхидных (Programma…, 1986; Blinova, 1998b; Bychenko, 2008a).
Для выявления экологических предпочтений D. fuchsii проведена обработка геоботанических описаний по индикационным экологическим шкалам Д.Н Цыганова (Tsyganov, 1983) и Г. Элленберга (Ellenberg, 1974), которые содержат балловые значения различных свойств факторов среды.
Экологическую валентность и толерантность вида определяли в соответствии с методикой Л.А. Жуковой (Zhukova, 2010). Для определения приспособленности ценопопуляции вида к изменению одного экологического фактора, рассчитывали потенциальную экологическую валентность (PEV) по формуле: PEV = (Аmax – Аmin + + 1)/n, где Аmax и Аmin – максимальные и минимальные значения ступеней шкалы, занятых видом, n – общее число ступеней в шкале. По результатам PEV определяли 5 фракций валентности (стено-, гемистено-, мезо-, гемиэври-, эвривалентная). Популяции с низкой экологической валентностью (стеновалентные виды) могут выносить ограниченные изменения фактора среды. Эвривалентные виды занимают местообитания с чрезвычайно изменчивыми условиями среды (Bychenko, 2008а). Реализованную экологическую валентность определяли по формуле: REV = = (Аmax – Аmin + 0.01)/n, где Amax и Amin – максимальные и минимальные значения ступеней шкалы, занятые конкретными ЦП; n – общее число ступеней в шкале.
Для выявления степени использования экологических потенций вида, определяли коэффициент экологической эффективности (K. ec. eff.), как отношение реализованной экологической валентности к потенциальной. Индекс толерантности (It) вида или его меру стено-эврибионтности определяли по формуле: It = ∑PEV/∑ шкал рассматриваемых факторов (Zhukova, 2005; 2010). Чем выше It, тем больше лабильность вида по отношению к рассматриваемым экологическим факторам (Bychenko, 2008b).
Гемеробность D. fuchsii в растительных сообществах определяли по составу видов (Frank, 1990).
Полученные данные обработаны статистически с использованием традиционных методов (Zaitsev, 1984; Borovikov, 2001).
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
На территории ГПЗ “Былина” D. fuchsii встречается в зеленомошных еловых, смешанных и сосновых лесах (Tarasova, 2005). Отмечен по опушкам ельников, в разреженных липово-еловых лесах с березой, елово-березовых лесах с осиной, изредка в осиново-еловых лесах на зарастающих лесных дорогах, по окраинам болот, на сырых луговинах и опушках (Kapustina et al., 2015; Kapustina, 2018).
Ценопопуляции D. fuchsii изучены в следующих фитоценозах (табл. 1). В составе травяно-кустарничкового яруса (ТКЯ) исследованных сообществ зафиксировано 123 вида сосудистых растений, среди которых отмечены и другие редкие виды из сем. Orchidaceae, такие как Epipogium aphyllum, Listеra cordаta, Platanthera bifolia, Dactylorhiza russowii, D. maculata, D. traunsteineri, Cypripedium calceolus. Общее проективное покрытие ТКЯ в сообществах с D. fuchsii изменяется от 30 до 90%; средняя высота травостоя варьирует от 8 до 76 см; проективное покрытие D. fuchsii от 10 до 35%.
Таблица 1.
Характеристика исследованных местообитаний Dactylorhiza fuchsii в ГПЗ “Былина” Кировской области Table 1. Characteristics of studied Dactylorhiza fuchsii habitats in “Bylina” State Nature Reserve in Kirov Region
| № ЦП No. CP |
№ на карте No. on map |
Год
наблюдения Year of study |
Тип фитоценоза (Состав древостоя) Plant community type (Tree stand composition) |
Доминирующие виды травяно-кустарничкового яруса (ТКЯ) Dominants of herbaceous-shrub layer (HSL) |
Общее ПП ТКЯ, % Total cover of HSL, % |
Число видов ТКЯ Number of species in HSL |
Доминирующие виды подлеска Dominants of undergrowth layer |
|---|---|---|---|---|---|---|---|
| 1 | 1 | 2012 | Елово-липовый бруснично-черничный (Л6 Е4)/Spruce-linden cowberry-bilberry forest (L6 S4) | Lathyrus vernus, Vaccinium myrtillus, Vaccinium vitis-idaea, Maianthemum bifolium, Athyrium fílix-femina, Cirsium oleraceum | 45 | 17 | Sorbus aucuparia |
| 2014 | |||||||
| 2 | 2 | 2012 | Липняк бруснично-черничный (Л8 + Е + Б)/Linden cowberry-bilberry forest (L8 + S + B) | Vaccinium myrtillus, Vaccinium vitis-idaea, Angelica sylvestris, Geranium sylvaticum, Equisetum sylvaticum, Fragaria vesca, Maianthemum bifolium | 40 | 19 | Viburnum opulus, Alnus incana |
| 2014 | |||||||
| 3 | 15 | 2012 | Елово-березово-осиновый снытево-разнотравный (Ос6 Е1 Б2 Л1)/Spruce-birch-aspen Aegopodium-mixed herbs forest (As6 S1 B2 L1) | Aegopodium podagraria, Lathyrus vernus, Viola mirabilis, Melampyrum pretense, Vaccinium vitis-idaea, Poa trivialis | 90 | 13 | Sorbus aucuparia |
| 2014 | |||||||
| 4 | 16 | 2012 | Липово-еловый чернично-кисличный (Е6 Л4)/Linden-spruce bilberry-oxalis forest (S6 L4) | Vaccinium myrtillus, Solidago virgaurea, Oxalis acetosella, Cirsium oleraceum, Equisetum sylvaticum, Maianthemum bifolium, Adoxa moschatellina | 90 | 15 | Sorbus aucuparia |
| 2014 | |||||||
| 5 | 27 | 2012 | Осиново-березовый брусничник зеленомошный старовозрастной (Б7 Ос3)/Aspen-birch cowberry green moss old forest (B7 As3) | Vaccinium vitis-idaea, Vaccinium myrtillus, Trientalis europaea, Lathyrus vernus, Oxalis acetosella, Melampyrum silvaticum, Poa trivialis, Linnaea borealis | 35 | 20 | Sorbus aucuparia |
| 2014 | |||||||
| 6 | 28 | 2012 | Осиново-елово-сосновый бруснично-майниковый (С6 Е2 О2)/Aspen-spruce-pine cowberry-maianthemum forest (P6 S2 As2) | Vaccinium vitis-idaea, Alchemilla vulgaris, Linnaea borealis, Melampyrum silvaticum, Equisetum sylvaticum, Maianthemum bifolium, Angelica sylvestris, Oxalis acetosella | 70 | 19 | – |
| 2014 | |||||||
| 2017 | |||||||
| 7 | 30 | 2012 | Осиново-березово-еловый разнотравный (Е6 Б2 Ос2)/Aspen-birch herbaceous-maianthemum forest (S6 B2 As2) | Vaccinium myrtillus, Vaccinium vitis-idaea, Aegopodium podagraria, Aconitum septentrionаle, Lathyrus vernus | 70 | 22 | Sorbus aucuparia, Rosa acicularis |
| 2014 | |||||||
| 2016 | |||||||
| 2017 | |||||||
| 2018 | |||||||
| 8 | 29 | 2012 | Елово-березовый травяно-майниковый (Б5 Е5 + Л)/Spruce-birch herbaceous-maianthemum forest (B5 S5 + L) | Maianthemum bifolium, Oxalis acetosella, Asarum europaeum, Equisetum sylvaticum, Aconitum septentrionаle | 60 | 13 | – |
| 2014 | |||||||
| 9 | 3 | 2016 | Березово-липово-еловый разнотравный (Е5 Л3 Б2)/Birch-linden-spruce mixed herbs forest (S5 L3 B2) | Chamerion angustifolium, Lathyrus pratensis, Hieracium umbellatum, Athyrium fílix-femina, Maianthemum bifolium, Angelica sylvestris, Chamerion angustifolium | 60 | 13 | – |
| 2018 | |||||||
| 10 | 4 | 2016 | Елово-березовый марьянниково-разнотравный (Б7 Е3)/Spruce-birch cowwheat-mixed herbs forest (B7 S3) | Oxalis acetosella, Maianthemum bifolium, Melampyrum silvaticum, Aegopodium podagraria, Trientalis europaea, Angelica sylvestris | 80 | 16 | Sorbus aucuparia, Salix caprea |
| 2018 | |||||||
| 11 | 5 | 2016 | Березово-еловый сочевичниково-разнотравный (Е6 Б4)/Birch-spruce orobus-mixed herbs forest (S6 B4) | Ranunculus repens, Lathyrus vernus, Thelypteris palustris, Trientalis europaea, Geranium sylvaticum | 75 | 11 | Salix caprea, Frangula alnus |
| 2018 | |||||||
| 12 | 19 | 2016 | Елово-березовый разнотравно-долгомошный (Б6 Е4)/Spruce-birch mixed herbs mossy forest (B6 S4) | Vaccinium vitis-idaea, Rubus arcticus, Athyrium fílix-femina, Carex aquatilis, Thelypteris palustris, Equisetum sylvaticum | 95 | 16 | Rubus idaeus, Sorbus aucuparia, Juniperus communis |
| 2017 | |||||||
| 2018 | |||||||
| 13 | 20 | 2016 | Осиново-елово-березовый марьянниково-разнотравно- долгомошный (Б6 Е3 Ос1)/Aspen-spruce-birch cowwheat-mossy forest (B6 S3 As1) | Rubus saxatilis, Carex aquatilis, Vaccinium myrtillus, Oxalis acetosella, Melampyrum silvaticum | 80 | 13 | – |
| 2017 | |||||||
| 14 | 21 | 2016 | Березово-еловый снытево-разнотравный (Б7 Е3)/Birch-spruce Aegopodium-mixed herbs (B7 S3) | Cypripedium calceolus, Pyrola rotundifolia, Carex dioica, Aegopodium podagraria, Thelypteris palustris | 80 | 22 | Sorbus aucuparia, Salix caprea |
| 2018 | |||||||
| 15 | 22 | 2017 | Березово-еловый бруснично-черничный долгомошный (Е8 Б2)/Birch-spruce cowberry-mossy forest (S8 B2) | Vaccinium vitis-idaea, Vaccinium myrtillus, Asarum europaeum, Maianthemum bifolium, Carex dioica, Thelypteris palustris | 60 | 25 | Juniperus communis, Sorbus aucuparia, Rubus idaeus |
| 2018 | |||||||
| 16 | 25 | 2014 | Липово-березово-еловый сочевичниковый (Е7 Б2 Л1)/Linden-birch-spruce orobus forest (S7 B2 L1) | Geranium sylvaticum, Fragaria vesca, Melampyrum silvaticum, Lathyrus vernus, Angelica sylvestris, Alchemilla vulgaris | 70 | 10 | Sorbus aucuparia, Rosa acicularis |
| 2016 | |||||||
| 2017 | |||||||
| 17 | 26 | 2014 | Березово-еловый разнотравный (Е7 Б3)/Birch-spruce mixed herbs forest (S7 B3) | Vaccinium myrtillus, Adoxa moschatellina, Geranium sylvaticum, Vicia sepium | 50 | 19 | Sorbus aucuparia |
| 2017 | |||||||
| 18 | 24 | 2012 | Березово-липовый бруснично-черничный (Л5 Б5)/Birch-linden bilberry forest (L5 B5) | Vaccinium vitis-idaea, Vaccinium myrtillus, Oxalis acetosella, Milium effusum, Geranium sylvaticum | 75 | 17 | Alnus incana, Sorbus aucuparia, Lonicera xylosteum |
| 19 | 17 | Липняк травянистый (Л10 + Е + Б)/ Linden herbaceous forest (L10 + S + B) | Oxalis acetosella, Asarum europaeum, Equisetum sylvaticum, Maianthemum bifolium | 30 | 11 | Lonicera caerulea | |
| 20 | 6 | Суходольный разнотравно-злаковый луг/Dry mixed herbs-gramineous meadow | Bromopsis inermis, Poa pratensis, Lupinus polyphyllus, Achillea millefolium, Geranium pratense | 90 | 14 | – | |
| 21 | 7 | Разнотравный луг формирующийся (зарастающая полевая дорога)/ Forming mixed herbs meadow (overgrowing field track) | Lysimachia vulgaris, Bidens tripartite, Urtica dioica, Medicago lupulina, Geranium pratense | 95 | 18 | – | |
| 22 | 8 | Елово-осиновый бруснично-черничный (Ос6 Е5)/Spruce-aspen cowberry-bilberry forest (As6 S5) | Vaccinium vitis-idaea, Vaccinium myrtillus, Equisetum sylvaticum, Oxalis acetosella | 45 | 13 | – | |
| 23 | 9 | Елово-березовый разнотравный (Б8 Е2)/Spruce-birch mixed herbs forest (B8 S2) | Oxalis acetosella, Asarum europaeum, Dryopteris filix-mas, Milium effusum | 40 | 12 | – | |
| 24 | 10 | 2014 | Березово-елово-осиновый черничник зеленомошный (Ос4 Е3 Б3)/Birch-spruce-aspen bilberry green moss forest (As4 S3 B3) | Vaccinium myrtillus, Maianthemum bifolium, Vicia sylvatica, Viola mirabilis, Ajuga reptans | 35 | 13 | – |
| 25 | 11 | Осиново-елово-сосновый разнотравный (С6 О2 Е2)/Aspen-spruce-pine mixed herbs forest (P6 As2 S2) | Fragaria vesca, Aegopodium podagraria, Solidago virgaurea, Maianthemum bifolium, Equisetum sylvaticum, Trientalis europaea | 85 | 19 | Sorbus aucuparia, Salix phylicifolia | |
| 26 | 12 | Елово-липово-осиновый разнотравный (Ос4 Е3 Л3)/Spruce-linden-aspen mixed herbs forest (As4 S3 L3) | Vaccinium vitis-idaea, Crepis paludosa, Myosotis nemorosa | 35 | 14 | Lonicera xylosteum, Sorbus aucuparia | |
| 27 | 13 | Елово-березовый кислично-майниковый (Б5Е5+Ос)/Spruce-birch oxalis-maianthemum forest (B5 S5+As) | Maianthemum bifolium, Oxalis acetosella, Asarum europaeum, Aconitum septentrionаle | 40 | 10 | – | |
| 28 | 14 | Липово-еловый чернично-кисличный (Е7 Л3)/Linden-spruce bilberry-oxalis forest (S7 L3) | Vaccinium myrtillus, Oxalis acetosella, Asarum europaeum, Dryopteris filix-mas | 55 | 15 | Lonicera xylosteum | |
| 29 | 23 | 2017 | Березово-еловый разнотравно-долгомошный (Е6 Б4)/Birch-spruce mixed herbs-mossy forest (S6 B4) | Rubus arcticus, Stellaria nemorum, Chamerion angustifolium, Juncus bufonius | 90 | 10 | Rosa acicularis, Juniperus communis, Frangula alnus |
| 30 | 18 | 2018 | Елово-березовый долгомошно-майниковый (Б6 Е4)/Spruce-birch mossy-maianthemum forest (B6 S4) | Vaccinium vitis-idaea, Maianthemum bifolium, Rubus arcticus, Asarum europaeum, Trientalis europaea, Dryopteris filix-mas | 70 | 24 | Lonicera xylosteum |
Изученные ЦП D. fuchsii приурочены к бореальным хвойным, мезофильным и мезоксерофильным широколиственным лесам в зоне умеренного климата относящимся к классам: Vaccinio-Piceetea Br.-Bl. in Br.-Bl., Siss. et Vlieger 1939, Querco-Fagetea Br.-Bl. et Vlieger in Vlieger 1937, Molinio-Arrhenatheretea R.Tx. 1937 em. R.Tx. 1970.
Класс Vaccinio-Piceetea – это бореальные хвойные леса на бедных кислых почвах с развитым моховым покровом. Порядок Piceetalia excelsae – европейские хвойные леса на бедных, кислых почвах.
Союз Piceion excelsae – мезофильные и гигрофильные темнохвойные леса на бедных кислых почвах. Подсоюз – Eu-Piceenion (мезофильные темнохвойные таежные леса на бедных кислых почвах). Подсоюз Melico-Piceenion – мезофильные темнохвойных таежные леса на относительно более богатых кислых почвах, с участием неморальных видов класса Querco-Fagetea. Союз Dicrano-Pinion (Подсоюз Dicrano-Pinenion) – сосняки на кислых почвах с доминированием мхов в нижнем ярусе.
Класс Querco-Fagetea – мезофильные и мезоксерофильные широколиственные листопадные леса на богатых почвах в зоне умеренного климата. Порядок Fagetalia sylvaticae – мезофильные широколиственные леса. Союз Querco roboris-Tilion cordatae – мезофильные широколиственные и смешанные леса Восточной Европы, описанные на Европейской части России и Латвии. Порядок Quercetalia roboris – европейские мезофильные дубовые и буковые леса на кислых почвах с присутствием видов классов Querco-Fagetea и Vaccinio-Piceetea, а также хорошо развитым ярусом напочвенных мхов.
Класс Molinio-Arrhenatheretea – это вторичные послелесные луга умеренной зоны Евразии, формирующиеся на месте широколиственных лесов на достаточно богатых незасоленных почвах. Порядок Molinietalia – влажные луга на минеральных почвах.
В составе травяно-кустарничкового яруса обследованных фитоценозов зафиксировано 123 вида сосудистых растений. Среди них преобладают следующие: Vaccinium vitis-idaea, V. myrtillus, Equisetum sylvaticum, Rubus arcticus, Fragaria vesca, Aconitum septentrionаle, Angelica sylvestris, Oxalis acetosella, Asarum europaeum.
Для D. fuchsii определены ступени шкал для каждой ценопопуляции и построена диаграмма, демонстрирующая фрагмент экологической ниши вида, границы исследуемых фитоценозов и границы экологического ареала вида по всем факторам (рис. 2, табл. 2).
Рис. 2.
Фрагмент экологической ниши Dactylorhiza fuchsii на территории ГПЗ “Былина” Кировской области (по шкалам Д.Н. Цыганова (1983)).
А – климатические шкалы и шкала освещенности-затенения; В – почвенные шкалы.
⋅ ⋅ ⋅ экологическая позиция изученных видов в фитоценозе по шкалам;
— экологическая позиция вида по шкалам.
Fig. 2. Part of Dactylorhiza fuchsii ecological niche in “Bylina” State Nature Reserve in Kirov Region (by D.N. Tsyganov’s scales, 1983).
А – climatic scale and light-shading scale; В – soil scales.
⋅ ⋅ ⋅ ecological position of studied species in community on scales;
— ecological position of the species on scales.
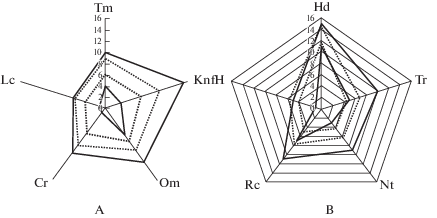
Таблица 2.
Характеристика экологических условий местообитания Dactylorhiza fuchsii на территории ГПЗ “Былина” Кировской области Table 2. Characteristics of ecological conditions of Dactylorhiza fuchsii habitats in “Bylina” State Nature Reserve in Kirov Region
| Экологическая шкала Ecological scale |
Экологическая позиция вида по шкале/Species ecological position on scale | PEV | REV | min–max изученных ЦП видов min–max of studied populations of different communities |
K. ec. eff. % | It | It общее It total |
|
|---|---|---|---|---|---|---|---|---|
| Климатичес-
кие шкалы Climatic scales |
Tm (17) | 4–10 | 0.41 | 0.16 | 6.17–8.95 | 39.95 | 0.61 ГЭБ HEB |
0.51 МБ MB |
| Kn (15) | 3–15 | 0.87 | 0.23 | 6.94–10.39 | 26.48 | |||
| Om (15) | 6–12 | 0.47 | 0.13 | 7.46–9.45 | 28.31 | |||
| Cr (15) | 1–10 | 0.67 | 0.18 | 5.70–8.45 | 27.44 | |||
| Почвенные
шкалы Soil scales |
Hd (23) | 11–15 | 0.22 | 0.12 | 11.92–14.55 | 52.24 | 0.40 ГCБ HSB | |
| Tr (19) | 5–10 | 0.32 | 0.13 | 4.38–6.87 | 41.14 | |||
| Nt (11) | 3–9 | 0.64 | 0.19 | 4.36–6.40 | 29.18 | |||
| Rc (13) | 7–11 | 0.38 | 0.20 | 5.12–7.66 | 51.65 | |||
| fH (11) | 1–5 | 0.45 | 0.14 | 4.01–5.54 | 31.07 | |||
| Шкала
освещенности-
затенения Light-shading scales |
Lc (9) | 1–6 | 0.67 | 0.22 | 3.70–5.70 | 33.33 | 0.67 ЭБ EB | |
Примечание. PEV – потенциальная экологическая валентность; REV – реализованная экологическая валентность; K. ec. eff. – коэффициент эффективности, %; It – индекс толерантности; ГЭБ – гемиэврибионт; ГСБ – гемистенобионт; МБ – мезобионт; ЭБ – эврибионт.
Note. PEV – potential ecological valence; REV – realized ecological valence; K. ec. eff. – efficacy coefficient, %; It – tolerance index; HEB – hemieurybiont; HSB – hemistenobiont; MB – mesobiont; EB – eurybiont.
В результате обработки описаний фитоценозов были получены следующие характеристики местообитания D. fuchsii во всех изученных ценопопуляциях по десяти шкалам Д.Н. Цыганова (табл. 2). D. fuchsii на территории заказника по отношению к комплексу всех экологических шкал относится к мезобионтным (МБ) видам (Itобщее = = 0.51). Вид имеет средний уровень лабильности по отношению к рассматриваемым экологическим факторам.
По совокупности климатических факторов и фактору освещенности-затенения D. fuchsii принадлежит к гемиэврибионтным (ГЭБ) и эврибионтным (ЭБ) видам соответственно. Полученные данные хорошо согласуются с результатами исследования других авторов (Ellenberg et al., 1991; Vakhrameeva, 2000), которые также отмечали широкую экологическую амплитуду D. fuchsii. Например, М.Г. Вахрамеева с соавторами (Vakhrameeva et al., 1994) отмечали, что ЦП этого вида встречаются как на полном свету, так и в полутени; произрастают как на сухих, так и влажных почвах, обитают на щелочных почвах, богатых основаниями, иногда на слабокислой, но никогда – на очень кислой почве.
Коэффициент экологической эффективности (K. ec. eff) колеблется не значительно. Максимально реализуют свои потенции ЦП D. fuchsii по термоклиматической шкале (K. ec. eff. = 39.95%). Минимальные значения K. ec. eff. отмечены для шкалы континентальности климата (26.48%).
Экологические условия изученных местообитаний большинства климатических шкал являются средними от потенциально возможных (рис. 2А). Диапазон по шкале освещенности (Lc) смещен к полуоткрытым пространствам и светлым лесам (3.70–5.70 балла). По шкале континентальности климата (Kn) территория исследования относится к зоне умерено-континентального климата (субматериковый, материковый вид) с диапазоном 6.94–10.39 балла. По термоклиматической шкале (Тm) D. fuchsii на территории ГПЗ “Былина” обитает в узких диапазонах от мезобореальной (6.17 балла) до неморальных (8.95 балла) климатических условий. ЦП вида на исследуемой территории обитают в условиях от субаридного (Om = 7.46 балла) до субгумидного (Om = 9.45 балла) климата с варьированием по криоклиматической шкале (Cr) от довольно суровых (5.70 балла) до умеренных зим (8.45 балла).
По отношению к комплексу почвенных факторов D. fuchsii является гемистенобионтным (ГСБ) видом (It = 0.40) (табл. 2). Коэффициент экологической эффективности (K. ec. eff) в рассматриваемых шкалах варьирует от 29.18 балла (по шкале богатства почв азотом) до 52.24 балла (по шкале увлажнения почв) и 51.65 балла (по шкале кислотности почв). Следует отметить, что по нескольким почвенным шкалам диапазон изученных местообитаний выходит за потенциально возможные границы (рис. 2Б). Так, например, по шкале кислотности (Rc) диапазон смещен в сторону кислых почв (5.12–7.66 балла); по шкале переменности почв (fH) в сторону умеренно переменного увлажнения (4.01–5.54 балла); по шкале солевого режима (Tr) в сторону бедных почв (гликосубме-зотрофная группа) (4.38–6.87 балла), что является подтверждением значительного адаптивного потенциала вида. ЦП данного вида могут произрастать в диапазоне от бедных (4.36 балла) до достаточно обеспеченных почв азотом (6.40 балла) при увлажнении (Hd) от сухолесолугового (11.92) до сыровато-лесолугового (14.55).
Для характеристики геометрического образа фундаментальной и реализованной экологических ниш, использованы диаграммы, построенные с использованием PEV и REV (Wittaker, 1980; Pianka, 1981). Экологическое пространство ЦП D. fuchsii (рис. 3) характеризуется широким потенциальным диапазоном по большинству рассматриваемых факторов.
Рис. 3.
Потенциальная и реализованная экологические валентности изученных ценопопуляций Dactylorhiza fuchsii на территории ГПЗ “Былина” Кировской области.
Fig. 3. Potential and realized ecological valences of studied Dactylorhiza fuchsii populations in “Bylina” State Nature Reserve in Kirov Region.
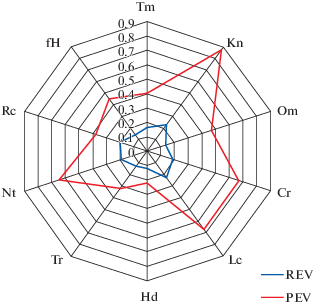
Почвенный фактор по шкале увлажнения (Hd) и шкале солевого режима (Tr) является лимитирующим (PEV = 0.22 и 0.32 соответственно), вид по данным шкалам является стеновалентным. D. fuchsii по шкалам Tm, Om, Rc, fH является гемистеновалентным, по шкале Nt – гемиэвривалентным, по шкалам Кn, Cr – эвривалентным видом.
Оценка экологических условий местообитаний по шкалам Г. Элленберга D. fuchsii по факторам увлажнения почв (Fе), богатству почв азотом (N), кислотности почв (R), освещенности (L) – континентальности климата (K) и фактору урбанизации (U) представлена на рис. 4.
Рис. 4.
Диаграмма экологического пространства Dactylorhiza fuchsii по шкалам Г. Элленберга (1974).
⋅ ⋅ ⋅ экологическая позиция вида в регионе исследования;
— экологическая позиция вида по шкалам Г. Элленберга.
Fig. 4. Dactylorhiza fuchsii ecological space chart by Ellenberg’s scales (1974).
⋅ ⋅ ⋅ species ecological position in the studied region;
— species ecological position by Ellenberg scales.
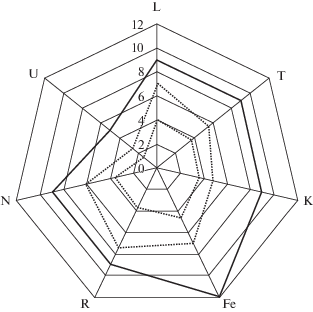
В результате оценки экологических условий местообитания D. fuchsii, выявлено, что популяции предпочитают условия от полутени (5-я ступень шкалы) до полусвета (7-я ступень). По отношению к шкале континентальности климата D. fuchsii выступает как слабо субконтинентальный вид (5-я ступень шкалы). D. fuchsii обитает на почвах умеренно богатых питательными веществами (5-я ступень шкалы), предпочитает почвы от кислых (3 ступень шкалы) до слабокислых (7-я ступень). По отношению к шкале влажности почв, предпочитает почвы от средне-влажных (5-я ступень шкалы) до влажных, хорошо пропитанных влагой, но не сырых (7-я ступень шкалы). По отношению к фактору урбанизации вид – умеренный урбанофоб (2-я ступень шкалы), обитает преимущественно вне поселений. Представленная оценка экологических условий местообитаний по шкалам Г. Элленберга согласуется с результатами исследований некоторых видов сем. Orchidaceae на территории Кировской (Chirkova et al., 2011; Egorova et al., 2014).
Для выявления антропотолерантности растительных сообществ с D. fuchsii использовали показатель гемеробии. При оценке уязвимости вида было установлено, что в условиях средней тайги во всех ЦП с D. fuchsii преобладают o (олиго-) и m (мезо-) гемеробные виды (42 и 37.1% соответственно), то есть виды, испытывающие незначительное и умеренное влияние человека. В-гемеробные виды составили 17.3%. Незначительную долю составили а-гемеробные виды (1%), не выносящие антропогенного воздействия и не устойчивые к незначительным спорадическим влияниям. С-p-гемеробы (сорные виды природных и антропогенных сообществ, переносящие регулярные сильные нарушения) составили 2.1 и 0.5%.
В целом, незначительная доля b-c-p-гемеробов говорит о достаточно низкой устойчивости D. fuchsii к антропогенному воздействию. Преобладание o-m-гемеробов указывает на способность вида испытывать незначительное и умеренное влияние человека. Относительная устойчивость вида к антропогенному воздействию и способность выдерживать слабые рекреационные нагрузки подтверждались в работах и других исследователей (Vakhrameeva, Varlygina, 1996).
В онтогенезе D. fuchsii были выделены все онтогенетические состояния от ювенильных (j) до сенильных (s). Возрастные спектры ЦП D. fuchsii за годы наблюдений в большинстве случаев нормальные неполночленные (правосторонние).
Возрастной спектр ценопопуляций в первый год наблюдений был бимодальным с двумя пиками, один из которых приходился на имматурные особи (37.7%), а другой пик – на генеративные особи (30.3%). В 2014 г. наблюдали изменение структуры ценопопуляций в генеративно-ориентированном направлении (доля генеративных особей составила 49%. К 2016 г. отмечен возрастной спектр так же в генеративно-ориентированном направлении, доля генеративных особей составляла (64.4%). В 2017 г. возрастной спектр был генеративно-ориентированным (57.8%). В 2018 г. возрастной спектр был бимодальным с двумя пиками на имматурном (17.5%) и генеративном (62.7%) онтогенетических состояниях.
ЗАКЛЮЧЕНИЕ
В условиях среднетаежной зоны Кировской области ценопопуляции Dactylorhiza fuchsii приурочены к бореальным хвойным лесам, относящимся к классу Vaccinio-Piceetea, мезофильным и мезоксерофильным широколиственным лесам, относящимся к классу Querco-Fagetea, и к вторичным послелесным лугам умеренной зоны Евразии, формирующимся на месте широколиственных лесов на достаточно богатых незасоленных почвах, относящимся к классу Molinio-Arrhenatheretea.
В травяно-кустарничковом ярусе исследованных фитоценозов с участием D. fuchsii отмечено 123 вида сосудистых растений, что составляет 8% от общего числа видов сосудистых растений в Кировской области.
В условиях региона исследований D. fuchsii осваивает достаточно широкий диапазон экологического пространства, как по экологическим шкалам Д.Н. Цыганова, так и по шкалам Г. Элленберга. По отношению к комплексу всех экологических шкал Д.Н. Цыганова D. fuchsii относится к мезобионтным видам (It–0.51).
Экологическое пространство D. fuchsii характеризуется широким потенциальным диапазоном по большинству рассматриваемых факторов, что позволяет виду занимать местообитания с чрезвычайно изменчивыми условиями среды. Диапазон изученных местообитаний выходит за потенциально возможные границы по нескольким почвенным шкалам: по шкале кислотности диапазон смещен в сторону кислых почв; по шкале переменности почв в сторону умеренно переменного увлажнения; по шкале солевого режима в сторону бедных почв.
D. fuchsii в регионе исследования по показателю гемеробности является видом с низкой устойчивостью к антропогенному воздействию, но переносящим незначительное и умеренное влияние человека.
В онтогенезе D. fuchsii были выделены все онтогенетические состояния от ювенильных (j) до сенильных (s). В возрастных спектрах ценопопуляций исследуемого вида преобладали имматурные и генеративные особи.
Список литературы
[Aver’yanov] Аверьянов Л.В. 2000. Орхидные (Orchidaceae) Средней России. – Turczaninowia. 1: 30–53.
[Blinova] Блинова И.В. 1998a. Особенности морфогенеза некоторых корнеклубневых орхидных (Orchidaceae). – Бот. журн. 83 (12): 83–91.
[Blinova] Блинова И.В. 1998b. Особенности онтогенеза некоторых корнеклубневых орхидных (Orchidaceae) Крайнего Севера. – Бот. журн. 83 (1): 85–89.
[Borovikov] Боровиков В.П. 2001. STATISTICA для студентов и инженеров. М. 301 с.
[Bychenko] Быченко Т.М. 2008а. Методы популяционного мониторинга редких и исчезающих видов растений Прибайкалья: учебное пособие. Иркутск. 164 с.
[Bychenko] Быченко Т.М. 2008b. Экологическое разнообразие ценопопуляций орхидных Прибайкалья. – В сб.: материалы III Всерос. научной конф. “Принципы и способы сохранения биоразнообразия”. Йошкар-Ола; Пущино. С. 313–316.
[Chirkova et al.] Чиркова Н.Ю., Сулейманова В.Н., Егошина Т.Л., Лугинина Е.А. 2011. Эколого-фитоценотическая и демографическая характеристика ценопопуляций Cypripedium calseolus L. (Сем. Orchidaceae) в условиях южно-таежных лесов Кировской области. – Вестник Тверского гос. ун-та. Серия “Биология и экология”. 24: 117–126.
[Egorova et al.] Егорова (Чиркова) Н.Ю., Сулейманова В.Н., Егошина Т.Л. 2014. Состояние ценопопуляций Platanthera bifolia (Orhidaceae) в Кировской области. – Раст. ресурсы. 50 (3): 398–414.
Ellenberg H. 1974. Zeigerwerte der Gefasspflanzen Mitteleuropas. Gottingen. 97 p.
Ellenberg H., Weber H.E., Dull R., Volkmar W., Willg W., Dirk P. 1991. Zeigerwerte von Pflanzen in Mitteleuropa. Scripta Geobotanica. 18 (1). 248 p.
Frank D., Klotz S. 1990. Biologisch-okologisch Daten zur Flora der DDR. Halle (Saale). 167 p.
[Gubanov et al.] Губанов И.А., Киселева К.В., Новиков В.С., Тихомиров В.Н. 2002. Иллюстрированный определитель растений Средней России. Том 1. Папоротники, хвощи, плауны, голосеменные, покрытосеменные (однодольные). М. 526 с.
[Il’ina, Dorogova] Ильина В.Н., Дорогова Ю.А. 2014. О положении ценопопуляций копеечника Гмелина (Hedysarum gmelinii Ledeb.) в экологическом пространстве (в условиях бассейна Средней Волги). – Известия Самарского научного центра Российской академии наук. Т. 14. 1 (7): 1745–1749.
[Il’ina] Ильина В.Н. 2019. Особенности структуры ценотических популяций Dactylorhiza fuchsia (Druce) Soo (Orchidaceae). – Международный научно-исследовательский журнал. 10 (88): 88–91.
[Ipatov] Ипатов В.С. 1998. Описание фитоценоза. Методические рекомендации. СПб. 93 с.
[Kapustina et al.] Капустина Н.В., Егорова Н.Ю., Егошина Т.Л. 2015. Состояние ценопопуляций Dactylorhiza fuchsii (Druce) Soo на территории ГПЗ “Былина”. – Вестник Оренбургского государственного университета. 6: 17–24.
[Kapustina] Капустина Н.В. 2018. Эколого-фитоценотическая характеристика Platanthera bifolia (L.) Rich на территории ГПЗ “Былина” Кировской области. – Самарский научный вестник. Т. 7. 1 (22): 45–50.
[Krasnaya kniga…] Красная книга Кировской области: животные, растения, грибы. 2014. Киров. 336 с.
[Metody izucheniya…] Методы изучения лесных сообществ. 2002. СПб. 240 с.
[Neshataev] Нешатаев Ю.Н. 1987. Методы анализа геоботанических материалов. СПб. 192 с.
[Ovesnov] Овеснов С.А. 1997. Конспект флоры Пермской области. Пермь. 252 с.
[Pianka] Пианка Э. 1981. Эволюционная экология. М. 400 с.
[Polevaya…] Полевая геоботаника. 1964. М. Т. 3. 530 с.
[Programma…] Программа и методика наблюдений за ценопопуляциями видов растений Красной книги СССР. 1986. М. 34 с.
[Tarasova] Тарасова Е.М. 2005. Флора государственного природного заказника “Былина”. Киров. 248 с.
[Tatarenko] Татаренко И.В. 1996. Орхидные России: жизненные формы, биология, вопросы охраны. М. 207 с.
The Plant List (TPL) http://www.theplantlist.org/
[Tsenopopuiyatsii…] Ценопопуляции растений (очерки популяционной биологии). 1988. М. 182 с.
[Tsyganov] Цыганов Д.Н. 1983. Фитоиндикация экологических режимов в подзоне хвойно-широколиственных лесов. М. 197 с.
[Vakhrameeva et al.] Вахрамеева М.Г., Варлыгина Т.И., Татаренко И.В. 2014. Орхидные России (биология, экология и охрана). М. 437 с.
[Vakhrameeva et al.] Вахрамеева М.Г., Денисова Л.В., Никитина С.В., Самсонов С.К. 1991. Орхидеи нашей страны. М. 244 с.
[Vakhrameeva et al.] Вахрамеева М.Г., Татаренко И.В., Быченко Т.М. 1994. Экологические характеристики некоторых видов евразиатских орхидных. – Бюл. МОИП. Отд. Биол. 99 (4): 75–82.
[Vakhrameeva, Varlygina] Вахрамеева М.Г., Варлыгина Т.И. 1996. Вопросы устойчивости и охраны и популяций орхидных. – Вестник МГУ. 16 (3): 30–35.
[Vakhrameeva] Вахрамеева М.Г. Род Пальчатокоренник // Биологическая флора Московской области. М., 2000. Вып. 14. С. 55–86.
[Wittaker] Уиттекер Р. 1980. Сообщества и экосистемы. М. 327 с.
[Zaitsev] Зайцев Г.Н. 1984. Математическая статистика в экспериментальной ботанике. М. 424 с.
Zeigerwerte von Pflanzen in Mitteleuropa. 1992. 2 Aufl. Göttingen. 258 p.
[Zhukova] Жукова Л.А. 2005. Экологическое разнообразие ценопопуляций модельных видов растений в Национальном парке “Марий Чодра” – В кн.: Биоразнообразие растений в экосистемах Национального парка “Марий Чодра”. Ч. 2. Йошкар-Ола. 196 с.
[Zhukova] Жукова Л.А., Дорогова Ю.А., Турмухаметова Н.В., Гаврилова М.Н., Полянская Т.А. 2010. Экологические шкалы и методы анализа экологического разнообразия растений: монография. Йошкар-Ола. 368 с.
[Zlobin] Злобин Ю.А. 1989. Принципы и методы изучения ценотических популяций растений. Казань. 146 с.
[Zubkova] Зубкова Е.В. 2011. О некоторых особенностях диапазонных экологических шкал растений Д.Н. Цыганова. – Известия Самарского научного центра Российской академии наук. 13 (5): 48–53.
Дополнительные материалы отсутствуют.
Инструменты
Ботанический журнал