

Ботанический журнал, 2020, T. 105, № 3, стр. 263-279
МЕЗОТРОФНЫЕ ЕЛЬНИКИ МОРОШКОВО-ХВОЩОВЫЕ СФАГНОВЫЕ СЕВЕРНОЙ И СРЕДНЕЙ ТАЙГИ ЕВРОПЕЙСКОЙ РОССИИ И УРАЛА
И. Б. Кучеров 1, *, С. А. Кутенков 2, **
1 Ботанический институт им. В.Л. Комарова РАН
197376 Санкт-Петербург, ул. Проф. Попова, 2, Россия
2 Институт биологии Карельского научного центра РАН
185910 Республика Карелия, Петрозаводск, ул. Пушкинская, 11, Россия
* E-mail: atragene@mail.ruIKucherov@binran.ru
** E-mail: effort@krc.karelia.ru
Поступила в редакцию 26.11.2019
После доработки 10.01.2020
Принята к публикации 14.01.2020
Аннотация
На основе 125 геоботанических описаний, сделанных авторами в 1995–2019 гг. или взятых из литературы, проведена доминантно-детерминантная классификация мезотрофных ельников морошково-хвощовых сфагновых с доминированием Picea abies s.l., Equisetum sylvaticum, Rubus chamaemorus и Sphagnum girgensohnii в Европейской России и на Урале. Выделены 2 ассоциации с 7 субассоциациями и 4 вариантами. Разнообразие синтаксонов обусловлено как широтной зональностью, так и изменениями континентальности климата, для субассоциаций и вариантов – также различиями во внутриландшафтной приуроченности. Ельники деренно-хвощовые сфагновые, где Equisetum sylvaticum и Rubus chamaemorus сопровождаются Chamaepericlymenum suecicum, свойственны преимущественно северной, морошково-хвощовые сфагновые без C. suecicum – также средней тайге. В Предуралье и на Урале на фоне роста континентальности климата в обеих ассоциациях выделяются субассоциации или варианты с участием Abies sibirica в древостое. Леса всех синтаксонов развиваются на начальных этапах мезотрофного заболачивания, тяготея к переувлажненным почвам умеренного минерального богатства.
Ельники хвощовые сфагновые – физиономически специфичный тип заболоченных еловых (из Picea abies s.l.) лесов, развивающихся на начальных этапах мезотрофного заболачивания, преимущественно при усилении проточности. Первые упоминания о нем в литературе, посвященной таежным лесам Европы, восходят к работам А. Каяндера (Cajander, 1913), Ф.В. Самбука (Sambuk, 1932) и Ю.Д. Цинзерлинга (Zinserling, 1932). Отличительной чертой данных ельников является господство Equisetum sylvaticum в кустарничково-травяном и Sphagnum girgensohnii в моховом ярусах. В таежной зоне Европейской России и Урала ельники хвощовые сфагновые произрастают в широком диапазоне зонально- и секторально-климатических и топоэдафических условий, что является предпосылкой их флористического и синтаксономического разнообразия.
По данным табличной обработки, совокупность описаний ельников хвощовых сфагновых, сделанных на упомянутой территории, распадается минимум на четыре ассоциации. Две из них, с совместным доминированием Equisetum sylvaticum и Rubus chamaemorus в кустарничково-травяном ярусе, свойственны преимущественно северной и средней тайге. Их анализу и посвящена данная статья.
ИСПОЛЬЗОВАННЫЕ ДАННЫЕ И МЕТОДЫ
В работе использована выборка из 125 геоботанических описаний мезотрофных ельников морошково-хвощовых сфагновых Европейской России и Урала, из них 84 выполнены авторами в экспедициях 1996–2019 гг. При описании на площади не менее 400 м2 оценивались проективные покрытия (ПП) видов по ярусам, делалась почвенная прикопка или измерялась мощность торфяной залежи. Подробное описание методики сбора полевых данных опубликовано ранее (Kucherov, 2019; Kucherov, Kutenkov, 2019 a). Часть описаний из Республики Коми (14) взята из рукописи докторской диссертации Ю.П. Юдина (Yudin, 1948), хранящейся в библиотеке БИН РАН. Содержание диссертации было издано без исходных описаний (Yudin, 1954). Еще 27 описаний заимствовано из опубликованных источников (Korchagin, 1929, 1940, 1956; Naumova, 1929; Sambuk, 1932; Soczava, 1930; Andreyev, 1935; Nekrasova, 1935; Smirnova A., 1954; Nepomilueva, 1970; Kolesnikov, 1985; Morozova, Korotkov, 1999).
Классификация растительности выполнена доминантно-детерминантным методом с выделением синтаксонов по доминантам и последующим уточнением их объема по детерминантным группам экологически близких видов (Vasilevich, Bibikova, 2004; Kucherov et al., 2010; Kucherov, Kutenkov, 2019a, 2019b). К одной ассоциации отнесены описания с одинаковым набором доминантов и детерминантов на всем протяжении ее ареала. У субассоциации могут быть собственные доминанты и детерминанты, отличные от таковых для ассоциации в целом (Kucherov, 2019), но, как правило, близкие к последним экологически, хотя и специфичные географически. Варианты могут выделяться только по детерминантам, модификации – по доминантам (Kucherov, Kutenkov, 2019a, 2019b).
Выделенные при классификации 9 синтаксонов разного ранга представлены в виде фитоценотической таблицы (табл. 1), где содержатся также сведения об общем покрытии и высоте ярусов. Названия синтаксонов даны согласно традиции школы В.Н. Сукачева (Sukachev, 1931). Названия в форме, принятой в работах по доминантно-детерминантной классификации (Vasilevich, Bibikova, 2004; Kucherov et al., 2010), приводятся как синонимы. При наименовании синтаксонов использован принятый в европейских работах принцип nomina conservanda (Kucherov, Kutenkov, 2019а, 2019b).
Таблица 1.
Фитоценотическая характеристика мезотрофных ельников деренно-морошково-хвощовых и морошково-хвощовых сфагновых Европейской России и Урала Table 1. Phytocoenotic record of mesotrophic dwarf cornel-cloudberry-horsetail and cloudberry-horsetail peatmoss spruce forests of European Russia and the Urals
| Вид/Параметр Species/Parameter | Ярус/Layer | Синтаксоны/Syntaxa | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | ||
| Детерминанты субассоциаций и вариантов/Subassociation and variant determinants | ||||||||||
| Empetrum nigrum s.l. | c | 602 | 603 | 33+ | 13+ | 12+ | ||||
| Vaccinium uliginosum | c | 455 | 8014 | 27+ | 8+ | |||||
| Comarum palustre | c | 20+ | 20+ | 27+ | 8+ | 20+ | 7+ | 4+ | ||
| Sphagnum warnstorfii | d | 205 | 4025 | 275 | 71 | 9+ | 81 | |||
| Betula pubescens s.l. | a2 | 403 | 672 | 313 | 331 | 331 | 271 | 333 | 12+ | |
| Salix myrsinifolia s.l. | a2 + b | 20+ | 33+ | |||||||
| Ledum palustre | c | 353 | 403 | 20+ | 13+ | 12+ | ||||
| Betula pubescens s.l. | b | 802 | 80+ | 671 | 693 | 671 | 671 | 181 | 681 | |
| Listera cordata | c | 25+ | 40+ | 33+ | 54+ | 47+ | 27+ | 40+ | ||
| Abies sibirica | a1+2 | 202 | 605 | |||||||
| A. sibirica | b | 807 | 806 | |||||||
| Melampyrum sylvaticum s.l. | c | 15+ | 40+ | 13+ | 8+ | 27+ | 13+ | 16+ | ||
| Oxycoccus palustris | c | 15+ | 40+ | 33+ | 4+ | |||||
| Sphagnum angustifolium | d | 51 | 803 | 6720 | 153 | 274 | 406 | 9+ | 17+ | 245 |
| Juniperus communis s.l. | b | 25+ | 733 | 151 | 7+ | 20+ | 9+ | 17+ | 4+ | |
| Salix phylicifolia | b | 151 | 332 | 7+ | 4+ | |||||
| Betula nana | b | 201 | ||||||||
| Equisetum palustre | c | 5+ | 9312 | 15+ | 20+ | |||||
| Aulacomnium palustre | d | 25+ | 67+ | 23+ | 13+ | 27+ | 9+ | 28+ | ||
| Sphagnum centrale | d | 101 | 534 | 31+ | 201 | 33+ | 361 | 4+ | ||
| Carex canescens | c | 25+ | 33+ | 46+ | 33+ | 7+ | 18+ | 8+ | ||
| Sphagnum riparium | d | 151 | 273 | 382 | 131 | 9+ | 163 | |||
| Pinus sylvestris | a1+2 | 252 | 674 | 8+ | 131 | 474 | 453 | 33+ | 8+ | |
| Picea abies s.l. | a2 | 304 | 202 | 7313 | 468 | 6011 | 679 | 646 | 839 | 243 |
| Calamagrostis purpurea s.l. | c | 25+ | 53+ | 692 | 47+ | 361 | 671 | 28+ | ||
| Luzula pilosa | c | 10+ | 60+ | 38+ | 60+ | 27+ | 18+ | 83+ | 24+ | |
| Solidago virgaurea s.l. | c | 20+ | 20+ | 60+ | 23+ | 47+ | 7+ | 83+ | ||
| Phegopteris connectilis | c | 311 | 202 | 9+ | 171 | |||||
| Maianthemum bifolium | c | 15+ | 33+ | 621 | 732 | 471 | 916 | 1009 | 522 | |
| Gymnocarpium dryopteris | c | 33+ | 1009 | 1009 | 27+ | 835 | 561 | |||
| Avenella flexuosa s.l. | c | 352 | 40+ | 20+ | 23+ | 531 | 7+ | 18+ | 33+ | 281 |
| Chamaenerion angustifolium | c | 15+ | 33+ | 31+ | 67+ | 18+ | 17+ | 12+ | ||
| Rubus arcticus | c | 10+ | 201 | 13+ | 15+ | 47+ | 7+ | 9+ | 17+ | 28+ |
| Pohlia nutans s.l. | z | 5+ | 7+ | 15+ | 40+ | 7+ | 18+ | 17+ | 24+ | |
| Sanionia uncinata | z | 7+ | 8+ | 33+ | 7+ | 27+ | 17+ | 20+ | ||
| Melampyrum pratense s.l. | c | 20+ | 20+ | 401 | 23+ | 53+ | 47+ | 17+ | 16+ | |
| Dicranum scoparium | d | 201 | 331 | 15+ | 331 | 531 | 181 | 33+ | 164 | |
| Sphagnum russowii | d | 309 | 131 | 8+ | 131 | 406 | 2712 | 282 | ||
| Oxalis acetosella | c | 15+ | 20+ | 531 | 551 | 8312 | 561 | |||
| Dicranum polysetum | d | 20+ | 13+ | 8+ | 401 | 271 | 675 | 12+ | ||
| Cladonia coniocraea | z | 5+ | 13+ | 8+ | 13+ | 40+ | 45+ | 67+ | 28+ | |
| Rubus idaeus | b | 9+ | 50+ | |||||||
| Dryopteris carthusiana | c | 13+ | 31+ | 824 | 671 | 81 | ||||
| Plagiothecium laetum | z | 10+ | 7+ | 15+ | 33+ | 7+ | 55+ | 33+ | 12+ | |
| Dicranum scoparium | z | 5+ | 7+ | 7+ | 13+ | 36+ | 50+ | 20+ | ||
| Carex brunnescens | c | 8+ | 91 | 50+ | ||||||
| Plagiothecium laetum | d | 5+ | 20+ | 8+ | 7+ | 50+ | ||||
| Ptilidium pulcherrimum | z | 10+ | 27+ | 15+ | 27+ | 27+ | 18+ | 67+ | 24+ | |
| Tetraphis pellucida | z | 10+ | 20+ | 7+ | 18+ | 50+ | 20+ | |||
| Dicranum majus | z | 5+ | 7+ | 8+ | 13+ | 33+ | 27+ | 50+ | ||
| Pleurozium schreberi | z | 5+ | 7+ | 20+ | 13+ | 9+ | 501 | 16+ | ||
| Dryopteris expansa s.l. | c | 151 | 9+ | 10024 | 44+ | |||||
| Pinus sibirica | a1+2 | 764 | ||||||||
| P. sibirica | b | 601 | ||||||||
| Ptilium crista-castrensis | d | 20+ | 23+ | 20+ | 33+ | 271 | 441 | |||
| Детерминанты ассоциаций табл. 1/Determinants of associations in Table 1 | ||||||||||
| Chamaepericlymenum suecicum | c | 704 | 1009 | 602 | 1004 | 17+ | ||||
| Rubus chamaemorus | c | 9512 | 10016 | 8711 | 927 | 876 | 1008 | 1003 | 501 | 10011 |
| Lycopodium annotinum s.l. | c | 551 | 604 | 731 | 691 | 53+ | 40+ | 451 | 17+ | 52+ |
| Orthilia secunda | c | 501 | 80+ | 621 | 671 | 731 | 361 | 50+ | 24+ | |
| Детерминанты всех ассоциаций мезотрофных ельников хвощовых сфагновых Determinants of all the associations of mesotrophic horsetail-peat moss spruce forests |
||||||||||
| Equisetum sylvaticum | c | 10022 | 10016 | 9320 | 10029 | 9323 | 10034 | 10022 | 10035 | 10020 |
| Sphagnum girgensohnii | d | 7024 | 6034 | 7327 | 10051 | 10052 | 8747 | 8242 | 10068 | 8028 |
| Константные виды/Constants | ||||||||||
| Picea abies s.l. | a1 | 10032 | 10040 | 10029 | 10035 | 9336 | 10044 | 10043 | 10064 | 10044 |
| Betula pubescens s.l. | a1 | 9011 | 1006 | 9312 | 10010 | 8715 | 8013 | 8213 | 674 | 1008 |
| Sorbus aucuparia s.l. | a2 + b | 401 | 20+ | 53+ | 623 | 732 | 531 | 822 | 1006 | 802 |
| Picea abies s.l. | b | 956 | 805 | 1007 | 928 | 939 | 1007 | 9111 | 1005 | 10012 |
| Vaccinium myrtillus | c | 9012 | 1007 | 804 | 1009 | 879 | 10016 | 10012 | 10016 | 10010 |
| V. vitis-idaea | c | 1007 | 408 | 9311 | 854 | 1009 | 1005 | 1008 | 1002 | 803 |
| Carex globularis | c | 807 | 10019 | 1009 | 853 | 876 | 10014 | 643 | 831 | 10011 |
| Linnaea borealis | c | 701 | 20+ | 801 | 922 | 931 | 601 | 732 | 83+ | 882 |
| Trientalis europaea | c | 601 | 40+ | 60+ | 921 | 1001 | 731 | 911 | 1002 | 722 |
| Polytrichum commune | d | 8014 | 10015 | 874 | 697 | 806 | 1006 | 647 | 675 | 10014 |
| Pleurozium schreberi | d | 858 | 802 | 804 | 623 | 804 | 936 | 913 | 502 | 1009 |
| Hylocomium splendens | d | 904 | 402 | 8713 | 929 | 735 | 875 | 645 | 501 | 765 |
| Прочие виды/Companions | ||||||||||
| Populus tremula | a1+2 | 7+ | 232 | 201 | 7+ | 175 | ||||
| Alnus incana s.l. | a2 + b | 20+ | 27+ | 23+ | 7+ | 7+ | 33+ | |||
| Rosa acicularis | b | 10+ | 401 | 33+ | 23+ | 40+ | 13+ | 9+ | 50+ | 20+ |
| Pinus sylvestris | b | 20+ | 8+ | 4+ | ||||||
| Deschampsia cespitosa | c | 5+ | 33+ | 8+ | 13+ | 7+ | 33+ | |||
| Dactylorhiza maculata s.l. | c | 7+ | 8+ | 13+ | 13+ | 9+ | 33+ | |||
| Geranium sylvaticum s.l. | c | 13+ | 231 | 7+ | 17+ | |||||
| Rubus saxatilis | c | 31+ | 7+ | 33+ | ||||||
| Carex aquatilis | c | 20+ | 8+ | 4+ | ||||||
| Viola epipsila | c | 20+ | 9+ | 17+ | ||||||
| Carex loliacea | c | 13+ | 23+ | |||||||
| Equisetum fluviatile | c | 20+ | 7+ | |||||||
| Carex nigra | c | 13+ | 17+ | |||||||
| C. rhynchophysa | c | 8+ | 17+ | |||||||
| Rhizomnium pseudopunctatum | d | 10+ | 27+ | 15+ | 20+ | 27+ | 16+ | |||
| Dicranum majus | d | 15+ | 27+ | 31+ | 13+ | 18+ | 331 | 4+ | ||
| Sphagnum wulfianum | d | 10+ | 8+ | 13+ | 20+ | 366 | 12+ | |||
| S. fallax | d | 158 | 131 | 152 | 201 | 9+ | 121 | |||
| Rhytidiadelphus triquetrus | d | 5+ | 13+ | 231 | 7+ | 13+ | 18+ | 331 | 8+ | |
| Sphagnum flexuosum | d | 132 | 206 | 94 | 206 | |||||
| Straminergon stramineum | d | 10+ | 33+ | 8+ | 17+ | 4+ | ||||
| Sphagnum capillifolium | d | 102 | 13+ | 7+ | 201 | 4+ | ||||
| Calliergon cordifolium | d | 5+ | 13+ | 13+ | 13+ | 33+ | ||||
| Plagiomnium ellipticum | d | 15+ | 13+ | 8+ | 7+ | 9+ | ||||
| Pseudobryum cinclidioides | d | 5+ | 7+ | 8+ | 13+ | 33+ | ||||
| Sphagnum squarrosum | d | 202 | 8+ | 171 | ||||||
| Sciuro-hypnum curtum | d | 7+ | 331 | |||||||
| Dicranum fuscescens | z | 13+ | 15+ | 40+ | 33+ | 27+ | 44+ | |||
| Cladonia deformis | z | 10+ | 13+ | 8+ | 13+ | 33+ | 18+ | 12+ | ||
| Hylocomium splendens | z | 131 | 20+ | 18+ | 331 | 24+ | ||||
| Средние сомкнутость или покрытие ярусов, %/Average density or cover of layers, %: | ||||||||||
| – 1-го яруса древостоя/1st tree stand layer | 0.5 | 0.5 | 0.5 | 0.5 | 0.6 | 0.6 | 0.6 | 0.8 | 0.6 | |
| – 2-го яруса древостоя/2nd tree stand layer | 0.2 | 0.1 | 0.3 | 0.1 | 0.2 | 0.2 | 0.2 | 0.2 | 0.1 | |
| – подроста, подлеска/regrowth, undergrowth | 10 | 15 | 15 | 15 | 10 | 10 | 15 | 5 | 20 | |
| – кустарничково-травяного/dwarf shrub and herb layer | 70 | 75 | 70 | 70 | 65 | 75 | 65 | 90 | 60 | |
| – мохового/moss layer | 90 | 100 | 90 | 85 | 75 | 90 | 90 | 80 | 85 | |
| Средняя высота ярусов, м/Average height of layers, m: | ||||||||||
| – 1-го яруса древостоя/1st tree stand layer | 14 | 13 | 17 | 18 | 18 | 18 | 20 | 20 | 16 | |
| – 2-го яруса древостоя/2nd tree stand layer | 9 | 7 | 9 | 9 | 11 | 9 | 13 | 9 | 13 | |
| – подроста, подлеска/regrowth, undergrowth | 2.4 | 1.8 | 3.5 | 2.4 | 2.3 | 2.1 | 1.4 | 1.3 | 3.2 | |
| Бонитет древостоя/Tree stand quality class | V | V | IV | IV | IV | IV | III | III | IV | |
| Мощность торфа, см/Peat layer thickness, cm | 30 | 95 | 35 | 35 | 25 | 30 | 30 | 20 | 25 | |
| Число описаний/Number of relevés | 20 | 5 | 15 | 13 | 15 | 15 | 11 | 6 | 25 | |
Примечания. Синтаксоны: 1–4 – Piceetum (P.) sphagno girgensohnii-cornoso-equisetosum sylvaticae: 1–2 – empetrosum: 1 – var. typica, 2 – var. Abies sibirica; 3 – equisetosum palustris; 4 – gymnocarpiosum dryopteris; 5–9 – P. sphagno girgensohnii-chamaemoroso-equisetosum sylvaticae: 5 – gymnocarpiosum dryopteris; 6 – typicum; 7–8 – dryopteridosum carthusianae: 7 – var. typica, 8 – var. Dryopteris expansa; 9 – abietosum sibiricae.
Ярусы: a1 и a2 – 1-й и 2-й ярусы древостоя, b – подрост и подлесок, c – кустарничково-травяной, d – моховой, z – эпифитные и эпиксильные мхи и лишайники. Для видов приводятся постоянство (%) и (в верхнем регистре) среднее проективное покрытие (%). Детерминантные группы выделены серым фоном и жирной рамкой; виды в их пределах сортированы по ярусам, далее по убыванию встречаемости во всем массиве описаний таблицы. Для доминирующих видов значения покрытия и постоянства даны полужирным шрифтом. Среднее проективное покрытие видов менее 0.5% отмечено плюсом “+”. Исключены сопутствующие виды со встречаемостью менее 20% хотя бы в одном из синтаксонов, всходы деревьев и кустарников.
Notes. Syntaxa: 1–4 – Piceetum (P.) sphagno girgensohnii-cornoso-equisetosum sylvaticae: 1–2 – empetrosum: 1 – var. typica, 2 – var. Abies sibirica; 3 – equisetosum palustris; 4 – gymnocarpiosum dryopteris; 5–9 – P. sphagno girgensohnii-chamaemoroso-equisetosum sylvaticae: 5 – gymnocarpiosum dryopteris; 6 – typicum; 7–8 – dryopteridosum carthusianae: 7 – var. typica, 8 – var. Dryopteris expansa; 9 – abietosum sibiricae.
Layers: a1 and a2 – 1st and 2nd layers of treestand , b – regrowth and undergrowth, c – dwarf shrub and herb layer, d – moss layer, z – epiphytic and epixylic mosses and lichens. Constancy (%) and average cover (%, superior figures) are given for species. Determinant groups are marked by grey backgrounds and bold margins; species within them are sorted first by layer, then by descending occurrence in the whole table relevé set. Cover and constancy values for dominants are shown in bold. Average species cover less than 0.5% is given as “+”. Species with occurrence less than 20% at least in one of the syntaxa are excluded from the table together with tree and shrub seedlings.
Номенклатура сосудистых растений дана по С.К. Черепанову (Czerepanov, 1995), мхов – по М.С. Игнатову с соавторами (Ignatov et al., 2006), лишайников – по “Списку лихенофлоры России” (A checklist…, 2010). Европейские таежные популяции ели отнесены к единому комплексу Picea abies s.l. (Popov, 2005). Границы зон и подзон растительности приняты по Т.И. Исаченко и Е.М. Лавренко (Isachenko, Lavrenko, 1980) с последующими уточнениями (Safronova, Yurkovskaya, 2015). В подзоне северной тайги выделяются северная и южная полосы – крайнесеверная и “типичная” северная тайга (Yudin, 1948, 1954). Точечные ареалы синтаксонов представлены на рисунках (рис. 1, 2). Как и ранее (Kucherov, Kutenkov, 2019а, 2019b), для оценки климатических условий распространения выделенных единиц использованы суммы превышений среднесуточных температур над базовой температурой в 10°C (“суммы градусо-дней выше 10°C”) по данным глобальной сети спутниковой метеосъемки (NASA…, 2006). На основе данных этой же базы рассчитаны индексы континентальности Конрада (Tukhanen, 1980) (табл. 2).
Рис. 1.
Распространение мезотрофных ельников (1) и пихто-ельников (2) воронично-деренно-хвощовых, ельников болотнохвощовых (3) и папоротничково-деренно-хвощовых сфагновых (4) в Европейской России и на Урале.
Зоны и подзоны: I – тундра, II – лесотундра и подгольцовые редколесья, III – северная, IV – средняя, V – южная тайга. Картооснова (Isachenko, Lavrenko, 1980; Safronova, Yurkovskaya, 2015) объединяет зональные выделы и их высотно-поясные аналоги. Полоса крайнесеверной тайги в северной части подзоны III и (на Северо-Востоке Европейской России) в южной части зоны II не отражена на карте в силу недостатка данных о ее границах (Kucherov, Kutenkov, 2019b).
Fig. 1. Distribution of Piceetum (P.) sphagno girgensohnii-cornoso-equisetosum subass. empetrosum var. typica (1) and var. Abies sibirica (2), subass. equisetosum palustris (3), and subass. gymnocarpiosum dryopteris (4) in European Russia and the Urals.
Zones and subzones: I – tundra, II – forest-tundra and subalpine open woodlands, III – northern-boreal, IV – middle-boreal, V – southern-boreal. The base map (Isachenko, Lavrenko, 1980; Safronova, Yurkovskaya, 2015) is generalized by the aggregation of zonal / subzonal units together with their altitudinal analogues. The northernmost-boreal belt approximately corresponds to the northern part of subzone III and (in the NE European Russia) the southern part of zone II; its ranges are known insufficiently (Kucherov, Kutenkov, 2019b).
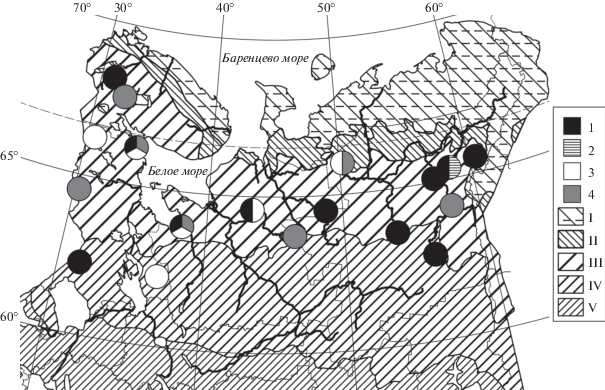
Рис. 2.
Распространение мезотрофных ельников папоротничково-морошково-хвощовых (5), типичных морошково-хвощовых (6) и крупнопапоротниково-морошково-хвощовых сфагновых типичного (7) и щитовникового (8) вариантов, а также пихто-ельников морошково-хвощовых сфагновых (9) в Европейской России и на Урале. Прочие обозначения как на рис. 1.
Fig. 2. Distribution of Piceetum (P.) sphagno girgensohnii-chamaemoroso-equisetosum subass. gymnocarpiosum dryopteris (5), typicum (6), dryopteridosum carthusianae var. typica (7) and var. Dryopteris expansa (8), and abietosum sibiricae (9) in European Russia and the Urals. For other notes and symbols see Fig. 1.
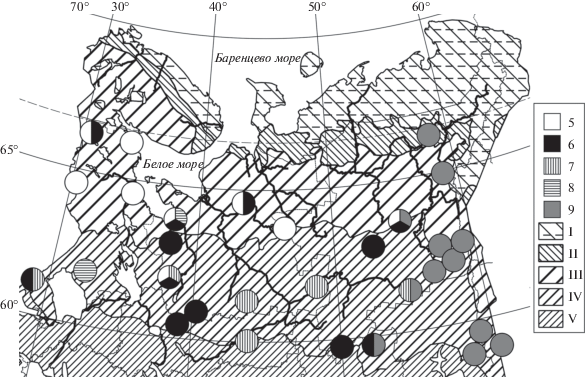
Таблица 2.
Амплитуды среднемноголетних (1983–2004) значений метеопараметров в пределах ареалов синтаксонов мезотрофных ельников хвощовых сфагновых Table 2. Amplitudes of averaged (1983–2004) climatic parameter values within the ranges of mesotrophic horsetail-peatmoss spruce forest syntaxa
| Синтаксоны/Syntaxa | Метеопараметры/Climatic parameters | |
|---|---|---|
| GDD > 10, °C | K | |
| 1. Piceetum (P.) sphagno girgensohnii-cornoso-equisetosum subass. empetrosum var. typica | 208–489 | 25.9–44.7 |
| 2. Var. Abies sibirica | 310–316 | 44.0–44.7 |
| 3. Subass. equisetosum palustris | 215–471 | 25.9–39.7 |
| 4. Subass. gymnocarpiosum dryopteris | 215–477 | 25.9–45.4 |
| 5. P. sphagno girgensohnii-chamaemoroso-equisetosum subass. gymnocarpiosum dryopteris | 281–544 | 31.2–44.1 |
| 6. Subass. typicum | 285–659 | 32.6–44.1 |
| 7. Subass. dryopteridosum carthusianae var. typica | 505–639 | 32.6–43.5 |
| 8. Var. Dryopteris expansa | 471–519 | 34.4–37.3 |
| 9. Subass. abietosum sibiricae | 283–635 | 41.8–45.7 |
Примечания. GDD > 10 – сумма градусо-дней выше 10°С (NASA, 2006), K – коэффициент континентальности Конрада (Tukhanen, 1980). Амплитуды рассчитаны с учетом как имеющейся совокупности описаний, так и данных литературы.
Notes. GDD > 10 – sum of growing degree-days above 10°С (NASA, 2006); K – Conrad continentality index (Tukhanen, 1980). Amplitude calculations are based upon both the examined relevé set and the published sources.
ОБЩАЯ ХАРАКТЕРИСТИКА ЕЛЬНИКОВ МОРОШКОВО-ХВОЩОВЫХ СФАГНОВЫХ
Ельники морошково-хвощовые сфагновые чаще всего растут по долинам лесных ручьев, в Карелии также в скрытопроточных межсельговых понижениях. Реже их можно встретить по окраинам мезотрофных болот при слабопроточном увлажнении; в этом случае наблюдаются начальные стадии заболачивания при наступлении болота на близлежащий суходол (Kutenkov, Kuznetsov, 2013). В северной тайге участки данных лесов встречаются и на заболоченных водоразделах, как правило, вблизи от склонов речных долин, а также на заболоченных участках пойменных и надпойменных террас.
Площадь, занятая ельниками морошково-хвощовыми, не слишком велика. Вместе с ельниками травяно-сфагнового типа они занимают лишь 1% от всей покрытой лесом площади в средней и северной тайге Карелии, тогда как все еловые леса – соответственно 47 и 24% в зависимости от подзоны (Gromtsev, 2008).
Нанорельеф обычно двухкомпонентный из невысоких кочек и приствольных повышений и волнистых сфагновых ковров, без западин (Kutenkov, Kuznetsov, 2013). На ветровальных участках высота возвышенных элементов нанорельефа возрастает до 1–2 м за счет вывалов, под которыми образуются западины. Почвы от торфянисто- и торфяно-подзолисто-глеевых до торфяно-глеевых (Andreyev, 1935; Dmitrieva, 1973), в Карелии также суглинистые элювиально-поверхностно-глеевые (Morozova, 1991). Мощность торфяной залежи от 20–50 см по берегам ручьев до 60–80 (Dmitrieva, 1973) и даже 200 см (Kutenkov, Kuznetsov, 2013) по окраинам мезотрофных болот.
Кислотность (pHH20) торфа в ельниках папоротничково-хвощовых сфагновых в горах Норвегии равна лишь 3.8 (Kielland-Lund, 1981). Однако в Карелии значения pHKCl в ельниках морошково-хвощовых сфагновых составляют 3.3–4.8 (Medvedeva et al., 1980), что сопоставимо с мезотрофными ельниками черничными сфагновыми на силикатах (Kucherov, Kutenkov, 2019b). В силу утяжеления гранулометрического состава плодородие почв выше, чем в упомянутых ельниках. Это позволяет мезотрофным видам расти как на ровных участках и в западинах, так и на микроповышениях.
Сомкнутость 1-го яруса древостоя обычно не превышает 0.5–0.6, а бонитет – IV. Постоянна примесь Betula pubescens к Picea abies s.l. от 1 до 3 единиц по составу. В западном и центральном секторах Европейской России в древостое наблюдается также примесь Pinus sylvestris. На востоке Русской равнины и на Урале рост континентальности климата приводит к формированию лесов с примесью Abies sibirica к Picea abies s.l. (в основном P. obovata) в древостое и “спутниками” пихты в кустарничково-травяном ярусе на плодородных почвах, свободных от мерзлоты. В результате на дифференциацию субассоциаций и вариантов ельников хвощовых сфагновых в отличие от черничных сфагновых (Kucherov, Kutenkov, 2019b) влияют не только широтная зональность, но и изменения континентальности климата. Как правило, выражен разреженный 2-й ярус древостоя, сложенный теми же (реже иными) видами, что и 1-й. Подрост еловый, иногда с примесью Betula pubescens и/или Abies sibirica в зависимости от синтаксона, групповой, с ПП до 10% (Kutenkov, Kuznetsov, 2013). В нижнем подъярусе подлеска обычны Juniperus communis и Rosa acicularis.
Сообщества отличает характерный аспект ажурной зелени обильного Equisetum sylvaticum, растущего на сфагновых коврах. Одновременно с хвощом в кустарничково-травяном ярусе могут доминировать Dryopteris expansa s.l. и/или Gymnocarpium dryopteris, обычно выступающие детерминантами субассоциаций или вариантов. К числу доминантов и субдоминантов яруса относятся также Rubus chamaemorus и Chamaepericlymenum suecicum; по сочетанию этих видов выделяются отдельные ассоциации. На микроповышениях обильны Vaccinium myrtillus и V. vitis-idaea, постоянны виды таежного мелкотравья: Trientalis europaea, Linnaea borealis, Orthilia secunda, Lycopodium annotinum, в средней тайге также Oxalis acetosella. Доминант мохового яруса Sphagnum girgensohnii иногда замещается S. russowii, чаще растущим в примеси на кочках, как и в ельниках черничных сфагновых (Kucherov, Kutenkov, 2019b). На микроповышениях преобладают таежные зеленые мхи: Hylocomium splendens, Pleurozium schreberi, Dicranum polysetum. Polytrichum commune может расти как на кочках, так и в примеси в сфагновом ковре, а также формировать плотные подушки (Kutenkov, Kuznetsov, 2013).
Константами всех синтаксонов ельников морошково-хвощовых сфагновых выступают Picea abies s.l. и Betula pubescens s.l. в древостое, Picea abies также в составе подроста, Sorbus aucuparia s.l., Vaccinium myrtillus, V. vitis-idaea, Trientalis europaea, Linnaea borealis, Carex globularis, Pleurozium schreberi, Hylocomium splendens, Polytrichum commune. Доминирующие в приземных ярусах бореальные (Equisetum sylvaticum) и арктобореальные (Sphagnum girgensohnii) гигромезофиты одновременно входят в число детерминантов обеих выделенных ассоциаций (табл. 1).
От мезотрофных хвощовых сфагновых следует отличать мезоэвтрофные ельники таволгово-хвощовые, сходные физиономически, но намного более богатые флористически, с иным набором индикаторных видов в приземных ярусах. Это, прежде всего, Filipendula ulmaria s.l. и Sphagnum warnstorfii, также Geum rivale, Viola epipsila, Crepis paludosa, Athyrium filix-femina, Ranunculus repens, Calliergon cordifolium, Rhytidiadelphus triquetrus. Последние шесть видов в мезотрофных ельниках хвощовых (преимущественно в южнотаежных крупнопапоротниково-хвощовых, не обсуждаемых в данной статье) могут быть лишь детерминантами отдельных вариантов, тогда как в таволгово-хвощовых они выступают детерминантами ассоциации в целом.
ХАРАКТЕРИСТИКА ВЫДЕЛЕННЫХ СИНТАКСОНОВ
Piceetum (P.) sphagno girgensohnii-cornoso-equisetosum (Sphagno girgensohnii-Corno suecici-Equiseto-P.) – ельник (е.) деренно-хвощовый сфагновый (табл. 1, рис. 1: № 1–4). Северотаежные сообщества с единичными нахождениями в средней тайге. Могут расти как на песках и щебнистых отложениях, так и на тяжелых почвах. Древостой несколько разрежен (сомкнутость 1-го яруса не выше 0.5). Подрост и подлесок без отчетливой стратификации по высоте. Диагностический вид ассоциации – Chamaepericlymenum suecicum, гипоарктический пустошно-лесной мезотрофный гигромезофит. В то же время Rubus chamaemorus (гипоарктический тундрово-болотный гигромезофит) наряду с лесными мезофитами (Orthilia secunda, Lycopodium annotinum s.l.) выступает детерминантом как этой ассоциации, так и ельников морошково-хвощовых сфагновых без Chamaepericlymenum suecicum (табл. 1).
От мезотрофных ельников деренно-ерниковых хвощово-морошкового варианта (P. sphagno girgensohnii-cornoso-nanobetulosum var. Rubus chamaemorus (Kucherov, Kutenkov, 2019b)) деренно-хвощовые отличают отсутствие Betula nana, образующей ярус низкого подлеска, а также других детерминантов первого синтаксона (Rubus arcticus, Eriophorum vaginatum, Polytrichum strictum, Cladonia rangiferina s.l.), более высокий (IV–V против Va) бонитет 1-го яруса древостоя, наличие его 2-го яруса.
В составе ассоциации выделяются три субассоциации.
Subass. empetrosum – е. воронично-деренно-хвощовый сфагновый (табл. 1, рис. 1: № 1–2). В отличие от других субассоциаций, в древостое нет примеси Pinus sylvestris; во 2-м ярусе отсутствует Picea abies s.l. В половине описаний 2-й ярус не выражен. В кустарничково-травяном ярусе согосподствуют Equisetum sylvaticum, Rubus chamaemorus, Vaccinium myrtillus и Carex globularis. Спорадически обильны Vaccinium uliginosum и Empetrum nigrum s.l. – гипоарктические и гипоаркто-бореальные олиготрофные кустарнички микроповышений, детерминанты субассоциации. В то же время Comarum palustre (мезотрофный гигрофит) и Sphagnum warnstorfii (мезоэвтрофный мезогигрофит, растущий в примеси в сфагновом ковре) объединяют синтаксон с subass. equisetosum palustris (см. ниже). Выделены два варианта.
Ельники var. typica (табл. 1, рис. 1: № 1) встречаются по берегам ручьев, окраинам болот, реже по коренным берегам рек и заболоченным участкам водоразделов, по берегам Белого моря в приморской полосе, почти всегда на силикатных породах, на Тиманском кряже иногда на известняках. Мощность торфа 10–60 см. Нанорельеф кочковатый с равным соотношением невысоких (15–30 см) кочек и ковров либо преобладанием последних. Значительна примесь Betula pubescens к Picea abies s.l. (P. × fennica, к востоку от р. Северной Двины также P. obovata) в 1-м ярусе древостоя. Березой сложен и 2-й ярус. Подрост березово-еловый, в подлеске Sorbus aucuparia s.l. Chamaepericlymenum suecicum иногда входит в число доминантов, но чаще выступает сопутствующим видом, как и все виды лесного мелкотравья. В моховом ярусе доминирующий Sphagnum girgensohnii изредка замещается на S. russowii, что приводит к росту среднего ПП последнего вида. В число доминантов, обильных наравне со S. girgensohnii, могут входить и Pleurozium schreberi или Polytrichum commune; Hylocomium splendens – всегда сопутствующий вид.
Betula pubescens s.l. во 2-м ярусе древостоя, Salix myrsinifolia s.l. (в основном S. borealis) и Ledum palustre отличают вариант от предуральского var. Abies sibirica, одновременно сближая его с преимущественно карельской subass. equisetosum palustris.
В крайнесеверной тайге описания варианта сделаны в Лапландском заповеднике, в нижнем течении р. Усы (P. cornosum, P. myrtilloso-polytrichosum (Sambuk, 1932)), по рекам Кожве (P. chamaemoroso-equisetosum), Кожиму, руч. Седьель (P. uligini-vaccinioso-sphagnosum (Yudin, 1948, 1954)). В южной полосе северной тайги вариант отмечен в Керетском Беломорье, в низовьях р. Онеги и в Пинежском заповеднике, в последнем случае – на силикатной морене, заполняющей палеокарстовое понижение. Восточнее он найден на водоразделе Мезенской Пижмы и Четласа (P. equisetoso-sphagnosum (Korchagin, 1956)), на Среднем Тимане по р. Ижме (P. sphagnoso-equisetosum (Andreyev, 1935)) и на Средней Печоре (P. sphagno-equisetosum (Sambuk, 1932)). Единственная изолированная находка в средней тайге сделана в Карелии (Толвоярви).
Var. Abies sibirica – пихто-ельник воронично-деренно-хвощовый сфагновый (табл. 1, рис. 1: № 2). Сообщества окраин болот и понижений на водоразделах. Торфяная залежь мощнее: от 35–50 до 300 см; кочки высотой 30–35 см. Picea abies s.l. представлена P. obovata. В 1-м ярусе древостоя незначительная примесь не только Betula pubescens, но и Abies sibirica, при этом именно пихтой образован 2-й ярус. Подрост пихтово-еловый; подлесок почти не развит. Chamaepericlymenum suecicum всегда входит в число доминантов своего яруса. Покрытие Vaccinium myrtillus нередко снижается, что компенсируется ростом ПП замещающей ее V. vitis-idaea. Иногда доминантом 2-го порядка становится и Lycopodium annotinum. Случаев замещения Sphagnum girgensohnii на S. russowii не отмечено, но бывает обилен S. centrale. Pleurozium schreberi переходит на роль сопутствующего вида; Hylocomium splendens встречается лишь изредка. В то же время Polytrichum commune всегда выступает одним из доминантов.
Детерминанты варианта, отличающие его от прочих синтаксонов ельников деренно-хвощовых и сближающие с пихто-ельниками морошково-хвощовыми, – Abies sibirica в древостое и в подлеске и гипоаркто-бореальный Melampyrum carpathicum (M. sylvaticum s.l.). В то же время с subass. equisetosum palustris вариант объединяют Sphagnum angustifolium (в примеси в моховом ковре) и сопровождающий его болотный оксилофит Oxycoccus palustris. О негативной дифференциации варианта сказано выше. Ареал ограничен крайнесеверной (Седьель, Джимтуй; P. uligini-vaccinioso-sphagnosum, P. globulari-caricoso-chamaemorosum (Yudin, 1948, 1954)) и “типичной” северной (вдоль р. Большая Сыня (Naumova, 1929)) тайгой Приполярного Предуралья.
Subass. equisetosum palustris – е. болотнохвощовый сфагновый (табл. 1, рис. 1: № 3). Леса этой субассоциации также произрастают по окраинам болот, особенно аапа-типа, и вдоль ручьев, текущих через болота, реже в заболоченных понижениях речных террас и водоразделов, всегда на силикатах; намного сильнее обводнены. Мощность торфа 15–60 см. Соотношение кочек (высотой 20–50 см) и ковров изменчиво, часто нанорельеф сглажен. В 1-м ярусе древостоя к Picea abies s.l. значимо примешивается Betula pubescens, в меньшей степени Pinus sylvestris. Густота 2-го яруса возрастает; он еловый, как и умеренно высокий подрост. В подлеске обычны характерные для окраин аапа-болот Juniperus communis s.l. и Salix phylicifolia, изредка встречается Betula nana. Постоянна, но не обильна Sorbus aucuparia s.l.
В кустарничково-травяном ярусе на коврах согосподствуют Equisetum sylvaticum и E. palustre (мезоэвтрофный мезогигрофит). На кочках доминируют Rubus chamaemorus, Carex globularis и Vaccinium vitis-idaea; ПП брусники может достигать 25–50%. V. myrtillus становится доминантом 2-го порядка. Изредка к числу доминантов добавляется Ledum palustre, но Vaccinium uliginosum и Empetrum nigrum s.l. более не характерны. Chamaepericlymenum suecicum и виды лесного мелкотравья малообильны.
В моховом ярусе к Sphagnum girgensohnii парадоксальным образом примешиваются одновременно мезоэвтрофные (S. warnstorfii), топяные мезотрофные (S. riparium) и олиготрофные (S. angustifolium) виды сфагнов, в чем также ощущается влияние соседства аапа-болот. Очень редко к S. angustifolium переходит доминирование. На микроповышениях Pleurozium schreberi, Polytrichum commune и (редко) Sphagnum centrale играют роль доминантов 2-го порядка; обильнее их лишь Hylocomium splendens.
Субассоциации свойственна многосторонняя дифференциация. С предыдущим вариантом ее объединяют группы Comarum palustre–Sphagnum warnstorfii и Oxycoccus palustris–Sphagnum angusti-folium, с subass. empetrosum var. typica – группа Betula pubescens–Ledum palustre. С subass. gymnocarpiosum синтаксон сближают Carex canescens и Sphagnum riparium, а также растущий на коврах Calamagrostis purpurea s.l. и лесные мезофиты приствольных повышений Luzula pilosa и Solidago virgaurea s.l. Последние три вида – также детерминанты ельников папоротничково-морошково-хвощовых. От subass. empetrosum синтаксон отличает примесь Pinus sylvestris в древостое и подъем Picea abies во 2-й его ярус, что типично также для subass. gymnocarpiosum и для большинства синтаксонов ельников морошково-хвощовых. К собственным детерминантам субассоциации, помимо Equisetum palustre, относятся кустарники подлеска (Juniperus communis s.l., Salix phylicifolia, Betula nana) и олигомезотрофные мхи микроповышений (Aulacomnium palustre, Sphagnum centrale).
Ельники болотнохвощовые наиболее обычны в “типичной” северной тайге Керетского Беломорья, восточнее отмечены в болотистых левобережных низовьях Онеги и на заболоченной четвертичной террасе Средней Пинеги. В крайнесеверной тайге синтаксон найден в национальном парке (НП) “Паанаярви” (отнесение его территории к крайнесеверной тайге обосновано ранее с позиций набора отмеченных там синтаксонов (Kucherov, 2019)) и в Республике Коми на междуречье Космы и Тобыша (P. equisetoso-sphagnosum (Yudin, 1948, 1954)). Самое южное местонахождение выявлено в средней тайге карельской части НП “Водлозерский” (Шуелахта).
Subass. gymnocarpiosum dryopteris – е. папоротничково-деренно-хвощовый сфагновый (табл. 1, рис. 1: № 4). Сообщества долин ручьев и проточных межсельговых понижений, лишь на востоке ареала отмечены по окраинам болот и на заболоченных водоразделах, всегда на силикатных породах. Торф мощностью от 20–30 до более 100 см. Нанорельеф ровный или кочковатый с преобладанием ковров. Второй ярус древостоя березово-еловый, в остальном его состав и высота, а также ПП подроста Picea abies s.l. как у предыдущей субассоциации. Несколько возрастает ПП подроста Betula pubescens. В подлеске Sorbus aucuparia s.l., редко также Alnus incana, Rosa acicularis. В кустарничково-травяном ярусе на коврах доминирует лишь Equisetum sylvaticum, на микроповышениях – Vaccinium myrtillus и мезотрофный лесной мезофит Gymnocarpium dryopteris. Chamaepericlymenum suecicum и Vaccinium vitis-idaea служат доминантами 2-го порядка. Становится постоянным Maianthemum bifolium. Изредка встречается Phegopteris connectilis, объединяющий этот и следующий синтаксоны. ПП Rubus chamaemorus и Carex globularis снижается. В моховом ярусе безраздельно господствует Sphagnum girgensohnii; Polytrichum commune и Hylocomium splendens – доминанты 2-го порядка. Редко при стволах появляется Rhytidiadelphus triquetrus.
С папоротничково-морошково-хвощовыми ельниками помимо Phegopteris connectilis синтаксон объединяют и другие лесные мезофиты (Gymnocarpium dryopteris, Maianthemum bifolium). В то же время мезотрофные гигромезофиты и мезофиты из группы Calamagrostis purpurea–Luzula pilosa и гигрофиты из группы Carex canescens–Sphagnum riparium сближают субассоциацию также с ельниками болотнохвощовыми.
В крайнесеверной тайге леса данной субассоциации отмечены по склонам долин ручьев в Лапландском заповеднике (P. chamaemorosum (Nekrasova, 1935)) и на водоразделе близ устья р. Космы (P. linnaeani-dryopteridoso-sphagnosum (Yudin, 1948, 1954)). В южной полосе северной тайги они описаны в материковой Керетской Карелии и на островах Керетского архипелага, в низовьях Онеги, на Пинежско-Мезенском водоразделе по р. Нюхча и в Предуралье при слиянии рек Щугор и Вача-ёль (P. equisetoso-sphagnosum (Yudin, 1948, 1954)). В среднюю тайгу синтаксон не заходит.
P. sphagno girgensohnii-chamaemoroso-equisetosum (Sphagno girgensohnii-Rubo chamaemori-Equiseto-P.) – е. морошково-хвощовый сфагновый (табл. 1, рис. 2: № 5–9). Северо- и среднетаежная ассоциация, леса которой изредка встречаются также в крайнесеверной или в южной тайге. Торфяная залежь неглубокая, 20–40, лишь иногда до 60–100 см. Как и в предыдущей ассоциации, леса могут развиваться равно на легких и тяжелых почвах; стратификация подроста и подлеска по высоте не выражена. К константам ассоциации (см. выше) примыкает Maianthemum bifolium; о детерминантах также сказано выше. В древостое большинства субассоциаций есть примесь Pinus sylvestris в 1-м ярусе и 2-й ярус Picea abies s.l. Выделяются четыре субассоциации.
Subass. gymnocarpiosum dryopteris – е. папоротничково-морошково-хвощовый сфагновый (табл. 1, рис. 2: № 5). Более “южный” аналог ельников папоротничково-деренно-хвощовых. Леса развиваются вдоль ручьев и по ложбинам стока, по берегам рек, но не столь редки и по окраинам болот. По Карельскому берегу Белого моря представлены также в приморской полосе. Растут в основном на силикатных породах, очень редко на гипсах. В последнем случае обычно небольшая (до 20 см) мощность торфа достигает 60 см. Соотношение кочек и ковров в нанорельефе изменчиво, высота кочек 15–70 см; часто нанорельеф не выражен. В 1-м ярусе древостоя примесь Betula pubescens к Picea abies s.l. существенна, Pinus sylvestris – незначительна. Второй ярус еловый, как и ярус подроста и подлеска; в составе последнего лишь малообильные Sorbus aucuparia s.l. и Rosa acicularis. В несколько разреженном кустарничково-травяном ярусе лишь Equisetum sylvaticum играет роль доминанта 1-го порядка, тогда как Gymnocarpium dryopteris, Vaccinium myrtillus, V. vitis-idaea, Rubus chamaemorus и Carex globularis – 2-го. К постоянным видам лесного мелкотравья (Maianthemum bifolium, Trientalis europaea, Linnaea borealis, Luzula pilosa и др.) добавляется Melampyrum pratense. Моховой ярус тоже слегка разрежен, его доминанты те же, что и в предыдущем синтаксоне; Pleurozium schreberi не столь обилен.
Собственные детерминанты субассоциации – олигомезотрофы (бореальный психромезофит Avenella flexuosa) и мезотрофы: послепожарно-боровой (а на Урале горно-луговой) бореальный мезофит Chamaenerion angustifolium, гипоарктический гигромезофит Rubus arcticus, растущие при стволах и на валеже мхи Pohlia nutans и Sanionia uncinata. Gymnocarpium dryopteris, встречающийся реже Phegopteris connectilis и группа Calamagrostis purpurea–Luzula pilosa сближают субассоциацию с экологически близкими ельниками папоротничково-деренно-хвощовыми, а Melampyrum pratense s.l. и Dicranum scoparium – с типичной субассоциацией морошково-хвощовой ассоциации.
Ельники папоротничково-морошково-хвощовые обычны по всей южной (“типичной”) полосе северной тайги Европейской России. Из Костомукшского заповедника они приводятся как Rubo chamaemori-Piceetum typicum K.-Lund 1962 (Morozova, Korotkov, 1999), хотя, возможно, было бы корректнее описать их как вариант субассоциации dryopteridosum phegopteris K.-Lund 1962 этой же ассоциации. В приземных ярусах этих лесов в горах Юго-Восточной Норвегии постоянны Phegopteris connectilis и Gymnocarpium dryopteris вместе с Rubus chamaemorus и Equisetum sylvaticum по ковру из Sphagnum girgensohnii и S. angustifolium (Kielland-Lund, 1981).
Восточнее Костомукши леса неоднократно отмечены от Керетской и Кемской (P. equisetosum c Gymnocarpium dryopteris и Ledum palustre на торфе мощностью до 80 см (Rutkovskii, 1933)) Карелии и низовий Онеги до среднего течения Пинеги, Пинежско-Мезенского (Нюхча) и Печоро-Ижемского (Нижний Одес) водоразделов. При этом ни один из типов мезотрофных ельников хвощовых сфагновых не упомянут в монографии Д.Н. Сабурова (Saburov, 1972), т. е. в карстовых ландшафтах Пинежья они крайне редки. В горной крайнесеверной тайге Карелии ельники данной субассоциации обнаружены лишь один раз в НП “Паанаярви”, а в средней тайге Архангельской обл. – также лишь однажды в южной части НП “Кенозерский”. Считается, однако, что в долинах рек и на заболоченных водоразделах они изредка встречаются по всей северной и средней тайге Коми, включая Урал (P. filicoso-sphagnosum (Yudin, 1948, 1954; Rysin, Savelieva, 2002)), где, видимо, представлен иной вариант.
Ельники subass. typicum (табл. 1, рис. 2: № 6) с равной частотой встречаются в поймах лесных ручьев и по краю болот, иногда также в заболоченных понижениях в скальных ландшафтах, обычно на силикатных породах, намного реже на известняках и гипсах. Мощность торфа 15–50 см. Нанорельеф кочковатый с преобладанием ковров; высота кочек до 30 см. Примесь Betula pubescens к Picea abies s.l. не столь велика, как в древостоях предыдущего синтаксона. Второй ярус еловый, низкий. По сравнению с предыдущим синтаксоном снижается высота подроста и подлеска, одновременно возрастает ПП как Equisetum sylvaticum, так и Vaccinium myrtillus и Carex globularis. Rubus chamaemorus остается доминантом 2-го порядка, но Vaccinium vitis-idaea становится сопутствующим видом. Gymnocarpium dryopteris отсутствует; Luzula pilosa и Solidago virgaurea нетипичны. Взамен при стволах, а иногда и на коврах становится постоянной Oxalis acetosella, вместе c Dicranum polysetum и эпифитом Cladonia coniocraea отличая субассоциацию от предыдущей и сближая с ельниками крупнопапоротниковой субассоциации. В то же время группа Melampyrum pratense–Dicranum scoparium сближает субассоциацию с папоротничково-морошково-хвощовой. В собственную детерминантную группу входит один лишь Sphagnum russowii, который может примешиваться к господствующему S. girgensohnii, но замещает его очень редко. В то же время в северо-восточной части ареала субассоциации (Пинега, Одес) замещающим видом может выступать S. flexuosum, реже к S. girgensohnii примешивается S. angustifolium. Моховой ярус вновь смыкается. На кочках Pleurozium schreberi, Hylocomium splendens и Polytrichum commune представлены в равном обилии.
В южной полосе северной тайги типичные ельники морошково-хвощовые характерны для Двино-Печорского региона, где описаны в низовьях Онеги и в окрестностях оз. Кожозеро, на Средней Пинеге и Печоро-Ижемском водоразделе. При этом в крайнесеверной тайге единственное их описание сделано в Карелии в НП “Паанаярви”. В средней тайге сообщества найдены на северо-западе Карельского перешейка (P. equisetoso-caricoso-sphagnosum с Carex echinata в межсельговых понижениях (Dmitrieva, 1973)), далее на юге и юго-западе Архангельской обл. у ст. Коноша (P. sphagnosum (Sambuk, 1927)) и в северной части НП “Кенозерский”. Восточнее они отмечены на западе Коми по р. Выми (P. globulari-caricoso-chamaemorosum (Yudin, 1948, 1954)) и в Кайском р-не на севере Кировской обл. (P. equiseto-polytrichoso-sphagnosum (Smirnova A., 1954)). В южной тайге рассматриваемые леса известны из Вологодской обл. (“ельник сфагновый” на карбонатной морене водораздела Вожега–Сить (Gavrilov, Karpov, 1962)) и из Нагорского р-на Кировской обл. (P. equisetoso-sphagnosum (Smirnova A., 1954)).
Subass. dryopteridosum carthusianae – е. крупнопапоротниково-морошково-хвощовый сфагновый (табл. 1, рис. 2: № 7–8). Сообщества речных долин (включая древние террасы), нехарактерные для окраин болот. Встречаются как на силикатных, так и на карбонатных породах. Мощность торфа 20–40 см. Ковры и кочки высотой 15–50 см обычно сочетаются в равной пропорции. Сравнительно с типичной субассоциацией, сомкнутость 1-го яруса древостоя возрастает, одновременно высота подроста и подлеска еще более снижается. К Equisetum sylvaticum примешивается Dryopteris carthusiana. Maianthemum bifolium становится доминантом 2-го порядка, в то же время Rubus chamaemorus и Carex globularis переходят в разряд видов-спутников. Диагностические виды – мезотрофные мезофиты: бореонеморальные Dryopteris carthusiana и растущая на вывалах Rubus idaeus, бореальные и бореально-полизональные мхи при стволах (Plagiothecium laetum, Dicranum scoparium). Возможно, вайи Dryopteris spp. создают дополнительное затенение, из-за чего нехарактерны Listera cordata и подрост Betula pubescens, постоянные во всех прочих синтаксонах ельников деренно- и морошково-хвощовых (табл. 1). Выделены два варианта.
Var. typica (табл. 1, рис. 2: № 7). Примесь Betula pubescens к Picea abies s.l. в 1-м ярусе древостоя невелика, Pinus sylvestris – напротив, возрастает. Подрост Picea abies умеренно густой; ПП Sorbus × gorodkovii (S. aucuparia s.l.) в подлеске незначимо. Покрытие Equisetum sylvaticum и кустарничково-травяного яруса в целом несколько снижено. Как Vaccinium myrtillus, так и V. vitis-idaea – доминанты 2-го порядка. Oxalis acetosella малообильна. В моховом ковре к Sphagnum girgensohnii примешиваются S. russowii и/или S. wulfianum. Доминантами 2-го порядка при стволах выступают Polytrichum commune и Hylocomium splendens; Pleurozium schreberi – сопутствующий вид. Дифференциация как у субассоциации в целом.
Ельники варианта описаны в средней тайге на восток от северо-запада Карельского перешейка (P. equisetoso-filicoso-sphagnosum с господством Equisetum sylvaticum и Dryopteris carthusiana, но со Sphagnum fallax и Viola epipsila (Dmitrieva, 1973)). На юге Архангельской обл. они известны по всему НП “Кенозерский” (Equiseto-Piceetum (Kucherov et al., 2010)), в бассейне р. Устьи и в низовьях р. Вычегды. В средней тайге Коми подобные леса отмечены по притоку р. Нем (P. equisetoso-sphagnosum (Kolesnikov, 1985)) и в долине р. Северной Кельтмы (P. chamaemoroso-sphagnosum (Yudin, 1948, 1954)). В Вологодской обл. на пермских глинах по р. Илезе сообщества заходят и в южную тайгу (P. humidum permense (Korchagin, 1929)).
Var. Dryopteris expansa (табл. 1, рис. 2: № 8). Примесь Betula pubescens к Picea abies s.l. возрастает, Pinus sylvestris – напротив, становится единичной. Высота 2-го яруса и ПП яруса подроста и подлеска снижены; при этом существенно увеличиваются ПП Sorbus × gorodkovii и (в своем ярусе) Equisetum sylvaticum. Наряду с хвощом доминантами яруса выступают также Dryopteris expansa s.l. (в основном D. assimilis), Vaccinium myrtillus и Oxalis acetosella. Первый из названных трех видов растет в основном на коврах, последние два – “спускаются” на них с приствольных повышений. При стволах вновь обилен также Gymnocarpium dryopteris, в то же время снижаются ПП Vaccinium vitis-idaea и (вдвое) постоянство Rubus chamaemorus. В моховом ярусе доминантами 2-го порядка, сопровождающими Sphagnum girgensohnii, служат Polytrichum commune и Dicranum polysetum; прочие виды таежных зеленых мхов им сопутствуют. Дифференциальные виды – Dryopteris expansa s.l. и Gymnocarpium dryopteris, таежные мезотрофные мезофиты, объединяющие вариант с уральскими пихто-ельниками морошково-хвощовыми. В то же время группа Calamagrostis purpurea–Luzula pilosa и опять-таки Gymnocarpium dryopteris сближают его с ельниками папоротничково-морошково- и папоротничково-деренно-хвощовыми. В число собственных детерминантов входят мезофит Carex brunnescens на вывалах, Plagiothecium laetum в составе эпигейной синузии, эпифиты Ptilidium pulcherrimum, Tetraphis pellucida и растущие на валеже Dicranum majus и Pleurozium schreberi.
Ареал варианта выявлен недостаточно полно. На сегодня сообщества отмечены лишь в северной тайге низовий Онеги и в бывшем природном парке “Хийсъярви” на западе среднетаежной Карелии. В последнем случае леса растут на карбонатных породах и обогащены Molinia caerulea (Equisetum sylvaticum-korpi (Brandt, 1933)).
Subass. abietosum sibiricae – пихто-ельник морошково-хвощовый сфагновый (табл. 1, рис. 2: № 9). Замещает типичную и крупнопапоротниковую субассоциации в Предуралье и на Урале. Леса растут как по долинам ручьев и в поймах рек, так и на заболоченных междуречьях, склонах водоразделов, шлейфах склонов гор, с равной частотой на силикатах и на карбонатах. Мощность торфа 15–25, реже до 100 см. Нанорельеф мелкокочковатый с господством ковров. В древостое Picea abies s.l. представлена P. obovata. Примесь Betula pubescens и Pinus sibirica в составе 1-го яруса постоянна, но незначительна. Второй ярус смешанный из Picea obovata и Abies sibirica, отдельные пихты выходят и в 1-й ярус. Подрост пихтово-еловый. В подлеске малообильные Sorbus × gorodkovii и S. sibirica. Демутация пихто-ельников данного синтаксона, видимо, проходит через стадии Betula pubescens и (возможно) Pinus sibirica без участия P. sylvestris, вследствие чего в бассейне Печоры не отмечены сосняки хвощовые сфагновые (Kucherov, 2019), хотя ельники данного типа обычны.
В кустарничково-травяном ярусе сочетаются “северные” и “южные” черты. В нем согосподствуют Equisetum sylvaticum, Rubus chamaemorus, Carex globularis и Vaccinium myrtillus. Как V. viti-sidaea, так и Gymnocarpium dryopteris, Maianthemum bifolium и Oxalis acetosella являются постоянными спутниками наряду с Trientalis europaea, Linnaea borealis и другими видами мелкотравья. Dryopteris carthusiana, Rubus idaeus и Solidago virgaurea s.l. отсутствуют. Dryopteris expansa s.l. (как D. assimilis, так и D. expansa s.str.) постоянен, но не обилен. В моховом ярусе совместно доминируют Sphagnum girgensohnii (иногда замещаемый S. flexuosum) Polytrichum commune и Pleurozium schreberi; Hylo-comium splendens – доминант 2-го порядка.
Детерминанты субассоциации – лесообразователи сибирской тайги (Abies sibirica, Pinus sibirica) в древостое и в подросте и таежный мезофильный зеленый мох Ptilium crista-castrensis. Кроме того, Dryopteris expansa s.l. и Gymnocarpium dryopteris сближают субассоциацию с предыдущим (крупнопапоротниковым) синтаксоном.
Основная часть описаний выполнена в Республике Коми. В южной полосе северной тайги леса описаны на Печоро-Ижемском водоразделе, в Илычском участке Печоро-Илычского заповедника (P. globulari-caricoso-chamaemorosum (Korchagin, 1940)), на Северном Урале в верховьях Щугора, а также (в Ханты-Мансийском автономном округе) у подножия хребта Паснер (Sphagnopiceetum equisetosum (Soczava, 1930)). Единственное описание в крайнесеверной тайге сделано в долине р. Понью (левый приток Усы; Betuleto-Piceetum equisetoso-sphagnosum (Nepomilueva, 1970)).
В средней тайге Коми пихто-ельники морошково-хвощовые выявлены в верховьях Вычегды (P. equiseti-sphagnosum, P. polytrichi-sphagnosum (Kolesnikov, 1985)), бассейне Северной Кельтмы (P. chamaemoroso-sphagnosum (Yudin, 1948, 1954)) и в Якшинском и Верхнепечорском участках Печоро-Илычского заповедника, где отмечались и ранее (P. equisetosum (Sambuk, 1932)). Описаны они и из средней тайги Кировской обл. (P. equisetoso-sphagnosum, P. equisetoso-polytrichosum (Smirnova A., 1954)). Также эти леса нередки по всей горной северной и средней тайге Северного (P. equisetoso-sphagnosum (Rysin, Savelieva, 2002)) и Среднего Урала, в том числе в Свердловской обл. (Kolesnikov et al., 1973) и в заповеднике “Басеги” в Пермском крае.
Замещающий вариант с Picea obovata и Pinus sibirica без участия Abies sibirica в древостое и с обильным Ledum palustre в кустарничково-травяном ярусе описан на вечномерзлых почвах в северной тайге верхнего течения р. Таз в Западной Сибири (P. obovatae ledoso-sphagnosum var. equisetosum sylvaticae (Neshatayev et al., 2002)).
Без деления на субассоциации и варианты ельники морошково-хвощовые сфагновые Финляндии описаны как Equisetum sylvaticum-Bruchmoore (Cajander, 1913) и Equisetum sylvaticum spruce mires (Eurola et al., 1994), а зарубежной Фенноскандии в целом – как Picea abies-Equisetum sylvaticum-variant в рамках Picea abies-Vaccinium spp.-Sphagnum spp.-typ'а с согосподством Sphagnum girgensohnii и S. angustifolium в моховом ярусе (Påhlsson, 1994). Они приводятся также для всей Карелии как развитые в условиях значительной проточности (Zinserling, 1932).
От олиготрофных ельников хвощово-багульниковых (P. sphagno angustifolii-ledosum subass. equisetosum) и хвощово-морошково-черничных (P. sphagno angustifolii-myrtillosum var. Equisetum sylvaticum (Kucherov, Kutenkov, 2019a)) все синтаксоны ельников деренно- и морошково-хвощовых отличает преобладание Sphagnum girgensohnii, а не S. angustifolium в моховом ковре, намного меньшее покрытие Carex globularis, большее разнообразие и постоянство видов лесного мелкотравья (Orthilia secunda, Lycopodium annotinum s.l., в ряде синтаксонов также Maianthemum bifolium, Gymnocarpium dryopteris, Oxalis aceto-sella), не говоря уже о видах Dryopteris.
ЗАКЛЮЧЕНИЕ
Всего в Европейской России и на Урале нами описаны 2 ассоциации мезотрофных ельников морошково-хвощовых сфагновых с 7 субассоциациями и 4 вариантами.
С точки зрения широтной зональности, всем четырем синтаксонам ельников деренно-хвощовых свойственны сугубо либо преимущественно северотаежные ареалы, охватывающие и крайнесеверную тайгу, и южную полосу подзоны. Ареалы четырех синтаксонов ельников морошково-хвощовых из пяти приурочены к южной полосе северной тайги вкупе со средней тайгой; лишь ельники крупнопапоротниково-морошково-хвощовые var. typica свойственны средней и южной тайге. Как и в ельниках черничных сфагновых (Kucherov, Kutenkov, 2019b), амплитуды толерантности сообществ всех синтаксонов по отношению к фактору теплообеспеченности соответствуют их зональным ареалам (табл. 2).
С точки зрения долготного распространения, лишь ельники деренно-хвощовые subass. gymnocarpiosum и subass. empetrosum var. typica, а также морошково-хвощовые subass. gymnocarpiosum со сравнительно “северными” ареалами распространены по всей Европейской России до Урала включительно. Всем остальным синтаксонам свойствен тот или иной тип секторальной приуроченности. Так, ельники крупнопапоротниково-морошково-хвощовые var. Dryopteris expansa встречаются лишь в Карелии и на северо-западе Архангельской обл. Ельники болотнохвощовые распространены от Карелии до бассейна Мезени, а морошково-хвощовые subass. typicum и subass. Dryopteris carthusiana var. typica – до левобережья Печоры. В то же время ельники деренно-хвощовые subass. empetrosum var. Abies sibirica встречаются только в Предуралье, морошково-хвощовые пихтовой субассоциации – также на Урале.
Распространение и взаимное замещение большинства синтаксонов однозначно соответствуют изменениям континентальности климата (табл. 2). С этой точки зрения ельники морошково-хвощовые сфагновые существенно отличаются от черничных сфагновых, ареалы которых не связаны с изменениями континентальности (Kucherov, Kutenkov, 2019b). Напрашивается предположение, что произрастание сообществ на богатых почвах способствует проявлениям влияния континентализации климата, тогда как тяготение к бедным почвам нивелирует последнее.
С точки зрения приуроченности синтаксонов к почвообразующим породам различных типов, ельники воронично-деренно-хвощовые var. typica, намного чаще встречаясь на силикатных породах, изредка растут и на карбонатных. Леса всех остальных субассоциаций и вариантов деренно-хвощовой ассоциации строго приурочены к силикатным породам. Ельники папоротничково-морошково-хвощовые изредка растут не только на силикатах, но и на гипсах, а типичные морошково-хвощовые – также на известняках. Леса оставшихся трех синтаксонов в равной мере успешно произрастают и на силикатных, и на карбонатных почвообразующих породах.
Стоит особо остановиться на том, что при разграничении синтаксонов наравне с эпигейными видами известную диагностическую роль играют также эпифитные мхи и лишайники, обычно исключаемые из синтаксономического анализа.
Список литературы
[A checklist…] Список лихенофлоры России. 2010. СПб. 194 с.
[Andreyev] Андреев В.Н. 1935. Лесная растительность южного Тимана. – Тр. Полярной комиссии СОПС АН СССР. 24: 7–64.
Brandt A. 1933. Hiisjarven luonnonpuiston kasvillisuudesta. – Silva Fenn. 32: 1–108.
Cajander A.K. 1913. Studien über die Moore Finnlands. – Acta Forest. Fenn. 2 (3): 1–208.
[Czerepanov] Черепанов С.К. 1995. Сосудистые растения России и сопредельных государств (в пределах бывшего СССР). СПб. 991 с.
[Dmitrieva] Дмитриева Е.В. 1973. Ельники северо-западной части Карельского перешейка. – Лесоведение. 2: 51–64.
Eurola S., Hicks S., Kaakinen E. 1984. Key to Finnish mire types. – In: European Mires. L. 117 p.
[Gavrilov, Karpov] Гаврилов К.А., Карпов В.Г. 1962. Главнейшие типы леса и почвы Вологодской области в районе распространения карбонатной морены. – Тр. ин-та леса и древесины. 52: 5–119.
[Gromtsev] Громцев А.Н. 2008. Основы ландшафтной экологии европейских таежных лесов России. Петрозаводск. 238 с.
Ignatov M.S., Afonina O.M., Ignatova E.A. 2006. Check-list of mosses of East Europe and North Asia. – Arctoa. 15: 1–130.
[Isachenko, Lavrenko] Исаченко Т.И., Лавренко Е.М. 1980. Ботанико-географическое районирование. – В кн.: Растительность европейской части СССР. Л. С. 10–22.
Kielland-Lund J. 1981. Die Waldgesellschaften SO Norwegens. – Phytocoenologia. 9 (1–2): 53–250.
[Kolesnikov] Колесников Б.П. 1985. Лесная растительность юго-восточной части бассейна Вычегды. Л. 216 с.
[Kolesnikov et al.] Колесников Б.П., Зубарева Р.С., Смологонов Е.П. 1973. Лесорастительные условия и типы леса Свердловской области. Свердловск. 176 с.
[Korchagin] Корчагин А.А. 1929. К вопросу о типах леса по исследованию в Тотемском уезде Вологодской губернии. – В кн.: Очерки по фитосоциологии и фитогеографии. М. С. 287–327.
[Korchagin] Корчагин А.А. 1940. Растительность северной половины Печорско-Ылычского заповедника. – В кн.: Труды Печорско-Ылычского заповедника. Т. 2. С. 5–415.
[Korchagin] Корчагин А.А. 1956. Еловые леса западного Притиманья в бассейне р. Мезенской Пижмы. – Учен. записки Ленингр. гос. ун-та. Сер. Геогр. 11: 111–239.
[Kucherov] Кучеров И.Б. 2019. Ценотическое и экологическое разнообразие светлохвойных лесов средней и северной тайги Европейской России. СПб. 568 с.
[Kucherov et al.] Кучеров И.Б., Разумовская А.В., Чуракова Е.Ю. 2010. Еловые леса национального парка “Кенозерский” (Архангельская область). – Бот. журн. 95 (9): 1268–1301.
[Kucherov, Kutenkov] Кучеров И.Б., Кутенков С.А. 2019a. Олиготрофные сфагновые и сфагново-зеленомошные ельники Европейской России и Урала. – Бот. журн. 104 (1): 12–40. https://doi.org/10.1134/S000681361901006X
[Kucherov, Kutenkov] Кучеров И.Б., Кутенков С.А. 2019b. Мезотрофные кустарничковые сфагновые и сфагново-зеленомошные ельники Европейской России и Урала. – Бот. журн. 104 (6): 859–887. https://doi.org/10.1134/S0006813619060115
[Kutenkov, Kuznetsov] Кутенков С.А., Кузнецов О.Л. 2013. Разнообразие и динамика заболоченных и болотных лесов Европейского Севера России. – В кн.: Разнообразие и динамика лесных экосистем России. Кн. 2. М. С. 152–206.
[Medvedeva et al.] Медведева В.М., Корнилова Л.И., Вайнблат В.З. 1980. Основные типы заболоченных ельников Карелии. – В кн.: Болотно-лесные системы Карелии и их динамика. Л. С. 78–99.
[Morozova, Korotkov] Морозова О.В., Коротков В.Н. 1999. Классификация лесной растительности Костомукшского заповедника. – Заповедное дело. 5: 56–78.
[Morozova] Морозова Р.М. 1991. Лесные почвы Карелии. Л. 184 с.
NASA surface meteorology and solar energy. 2006. http://eosweb.larc.nasa.gov (Accessed 15.05.2006).
[Naumova] Наумова С.Н. 1929. Ботанико-географические исследования 1926 года в предгорьях Северного Урала (р. Большая Сыня и Большой Оранец). – Тр. О-ва по изуч. Урала, Сибири и Дальнего Востока. М. 1: 103–157.
[Nekrasova] Некрасова Т.П. 1935. Очерк растительности Лапландского заповедника. – Тр. Ленингр. о-ва естествоиспыт. 64 (2): 239–272.
[Nepomilueva] Непомилуева Н.И. 1970. Кедр сибирский (Pinus sibirica Tour.) на северной границе ареала в Коми АССР. – Бот. журн. 55 (7): 1011–1025.
[Neshatayev et al.] Нешатаев В.Ю., Потокин А.Ф., Томаева И.Ф., Егоров А.А., Добрыш А.А., Чернядьева И.В., Потемкин А.Д. 2002. Растительность, флора и почвы Верхне-Тазовского государственного заповедника. СПб. 154 с.
Påhlsson L. (ed.) 1994. Vegetationstyper i Norden. Köpenhavn. 627 s.
[Popov] Попов П.П. 2005. Ель европейская и сибирская: структура, интерградация и дифференциация популяционных систем. Новосибирск. 231 с.
[Rutkovskii] Рутковский В.И. 1933. Типы лесов Кемского края АКССР. – Тр. Ин-та изучения леса АН СССР. 1: 1–97.
[Rysin, Savelieva] Рысин Л.П., Савельева Л.И. 2002. Еловые леса России. М. 335 с.
[Saburov] Сабуров Д.Н. 1972. Леса Пинеги. Л. 173 с.
[Safronova, Yurkovskaya] Сафронова И.Н., Юрковская Т.К. 2015. Зональные закономерности растительного покрова равнин Европейской России и их отображение на карте. – Бот. журн. 100 (11): 1121–1141.
[Sambuk] Самбук Ф.В. 1927. Растительные ассоциации на желто-подзолистых почвах Коношской дачи Вологодской губернии. – Журн. Русск. Бот. о-ва. 12 (1–2): 33–58.
[Sambuk] Самбук Ф.В. 1932. Печорские леса. – Тр. Бот. музея АН СССР. 24: 63–245.
[Smirnova] Смирнова А.Д. 1954. Типы еловых лесов крайнего севера Кировской области. Ч. II. – Учен. записки Горьковск. ун-та. 25: 191–226.
[Soczava] Сочава В.Б. 1930. К фитосоциологии темнохвойного леса. – Журн. Русск. бот. о-ва. 15 (1–2): 7–41.
[Sukachev] Сукачев В.Н. 1931. Руководство к исследованию типов леса. 3-е изд. М. 328 с.
Tukhanen S. 1980. Climatic parameters and indices in plant geography. – Acta Phytogeogr. Suec. 67: 1–105.
[Vasilevich, Bibikova] Василевич В.И., Бибикова Т.В. 2004. Сфагновые ельники Европейской России. – Бот. журн. 89 (5): 734–748.
[Yudin] Юдин Ю.П. 1948. Темнохвойные леса Коми АССР (геоботаническая характеристика): Дис. … докт. биол. наук. Сыктывкар. 323 с.
[Yudin] Юдин Ю.П. 1954. Темнохвойные леса. – В сб.: Производительные силы Коми АССР. М.; Л. Т. 3. Вып. 1. С. 42–125.
[Zinserling] Цинзерлинг Ю.Д. 1932. География растительного покрова северо-запада европейской части СССР. Л. 376 с.
Дополнительные материалы отсутствуют.
Инструменты
Ботанический журнал