

Ботанический журнал, 2021, T. 106, № 3, стр. 255-271
ПОКОЙ ЖИЗНЕСПОСОБНЫХ СЕМЯН В ПОЧВЕ И ИХ ПРОРАСТАНИЕ ПОСЛЕ ПОЖАРОВ В ШИРОКОЛИСТВЕННО-КЕДРОВЫХ ЛЕСАХ ЮЖНОГО СИХОТЭ-АЛИНЯ
Т. А. Комарова 1, *, Н. В. Терехина 2, **, Т. П. Орехова 1, ***
1 Федеральный научный центр биоразнообразия наземной биоты Восточной Азии ДВО РАН
690022 Приморский край, Владивосток, пр. 100-летия Владивостоку, 159, Россия
2 Институт наук о Земле СПбГУ
199178 Санкт-Петербург, 10-я линия Васильевского острова, 33/35, Россия
* E-mail: mata41@mail.ru
** E-mail: n.terehina@spbu.ru
*** E-mail: tp.orekhova@mail.ru
Поступила в редакцию 21.06.2020
После доработки 02.11.2020
Принята к публикации 10.11.2020
Аннотация
Обобщены собранные ранее материалы более 20-летних исследований почвенных банков семян на ранних, средних и поздних стадиях демутационных сукцессий после устойчивых низовых пожаров в широколиственно-кедровых лесах Южного Сихотэ-Алиня. Прослежены изменения в видовом составе, численности и вертикальном распределении всхожих семян в подстилке и почве, начиная с однолетней гари до старовозрастного широколиственно-кедрового леса более 350 лет. Установлено закономерное снижение состава и численности всхожих семян от ранних к поздним стадиям послепожарных демутационных сукцессий и резкое снижение числа погребенных покоящихся семян в старовозрастном лесу. Лесные пожары рассматриваются как своеобразный сигнал для активизации покоящихся в почве семян.
Важное место в возобновлении многих популяций растений после пожаров принадлежит покоящимся в почве семенам, выполняющим роль резерва, который активизируется в случае нарушения растительного покрова и подстилки. Свойство семян сохранять жизнеспособность в почве в течение длительного времени выработалось в процессе эволюции и служит характерным приспособлением для сохранения устойчивости популяций во времени при циклической смене условий произрастания (Rabotnov, 1981, 1986). Длительный покой семян обеспечивает растениям возможность пережить неблагоприятные условия среды на эмбриональной стадии развития как наиболее устойчивой к внешним воздействиям (Poptsov, 1968; Juan et al., 2006; Wellstein et al., 2007).
Совокупность жизнеспособных семян, содержащихся в подстилке и почве, большинством отечественных и зарубежных исследователей названа почвенным банком семян (для краткости ПБС). Изучение ПБС в лесных биогеоценозах европейской части СССР было начато с 60-х годов прошлого столетия (Karpov, 1960, 1969; Rysin, Rysina, 1965; Petrov, 1989). Анализировался запас семян в биогеоценозах сукцессионных рядов после рубки коренных лесов в европейской части России (Karpov, 1969; Petrova, 1999; Sidorova, 2009). Особенности экологии ПБС в лесных биогеоценозах Саратовской обл. исследовала О.Н. Торгашкова (Torgashkova, 2003, 2013).
Многочисленные исследования ПБС, особенно в последние два десятилетия, проведены за рубежом. Получены сведения о ПБС всех растительных зон от пустынь до Арктики и в растительных сообществах с широким экологическим диапазоном (Baskin, Baskin, 2001; Bossuyt, Hermy, 2001; Fenner, Thompson, 2005). Изучена динамика видового состава и численности ПБС в ходе вторичных сукцессий после лесных рубок в разных формациях (Grandin, 2001; Bossuyt et al., 2002; Plue et al., 2010). Менее изученными остались особенности динамики ПБС в процессе демутационных сукцессий после лесных пожаров. Большая часть исследований динамики ПБС после пожаров проведена либо на начальных стадиях сукцессий (Ferrandis et al., 1996; Kwiatkowska-Falińska et al., 2014), либо в саванах, редколесьях и пустошах (Ne’eman, Izhaki, 1999; Buhk, Hensen, 2005).
Нами, начиная с 1982 г., в течение более 20 лет проводилось изучение видового состава и запасов всхожих семян, а также спор папоротников в почвенных образцах, взятых в широколиственно-кедровых и кедрово-темнохвойных лесах Южного Сихотэ-Алиня, подвергавшихся пожарам разной давности. Результаты 4-летних и 8-летних наблюдений за прорастанием семян и спор папоротников в почвенных образцах, содержатся в опубликованных работах (Komarova, 1986, 1992).
В настоящей работе проводится более детальный анализ собранных ранее материалов по изучению ПБС, взятых с 6 участков широколиственно-кедровых лесов, подвергавшихся пожарам от 1 до 220 лет назад. Кроме того, впервые дается анализ видового состава и запасов ПБС в старовозрастном перестойном (более 350 лет) широколиственно-кедровом лесу, а также в 9‑летнем осиново-березовом молодняке, сформировавшемся после сплошной экспериментальной рубки в этом перестойном лесу. Проведен сравнительный анализ ПБС в 9-летних осиново-березовых молодняках, образовавшихся после сплошной рубки и в результате устойчивого низового пожара широколиственно-кедрового леса.
ХАРАКТЕРИСТИКА РАЙОНА ИССЛЕДОВАНИЯ
Исследования проводили в 1982–2010 гг. на территории Верхнеуссурийского биогеоценотического стационара ФНЦ Биоразнообразия ДВО РАН площадью около 4.5 тыс. га, и на прилегающих к нему территориях (43°09'–44°01'с.ш.). Район исследования находится на западном макросклоне Южного Сихотэ-Алиня в бассейнах рек Правая Соколовка и Извилинка (притоки р. Уссури), в пределах высотных отметок 450–1050 м над ур. моря.
Основные растительные сообщества, где проводились работы, – широколиственно-кедровые леса, представленные различными этапами послепожарных сукцессионных смен: осиново-березовый молодняк на месте мезофитного широколиственно-кедрового лианово-разнокустарникового василистниково-папоротниково-осокового леса, осиново-березовый лес на месте ксеромезофитного лимонниково-лещинного разнотравяно-мелко-осокового кедровника с дубом, ксеромезофитный кедровник с темнохвойными породами лимонниково-лещинный низкотравно-мелкоосоковый разных возрастов и перестойный ксеромезофитный кедровник с дубом лимонниково-лещинный разнотравно-мелкоосоковый.
Характеристики почв и подстилок изучаемых биогеоценозов приведены по материалам, полученным почвоведами на Верхнеуссурийском стационаре (Sapozhnikov, Gavrenkov, 1980; Sapozhnikov, Kostenkova, 1984; Soil formation, 1993).
Почвы под широколиственно-кедровыми лесами и их производными сообществами относятся к горно-лесным бурым, развивающимся на делювиальных склоновых отложениях. Гумусовый горизонт мощностью от 7 до 16 см, темно-бурого цвета, легко- или среднесуглинистый; содержание гумуса в нем колеблется от 15 до 32% (Sapozhnikov, Gavrenkov, 1980).
Для подстилок характерно сочетание трех подгоризонтов (О1-О2-О3), отличающихся друг от друга степенью разложения опада. Нижний подгоризонт (О3) представляет собой наиболее разложившийся плотный и влажный опад, постепенно переходящий в гумусовый горизонт почвы (AY). Подстилка в ходе послепожарных демутационных сукцессий меняется от фрагментарной до сплошной, мощностью до 10 см.
Бурые горно-лесные почвы значительно изменяются в зависимости от рельефа и характера растительного покрова. На выпуклых крутых инсолируемых склонах под мезоксерофитными дубово-кедровыми редкопокровными лесами и их производными сообществами (п.п. 30-1982) характерны слаборазвитые, сильноскелетные, легкосуглинистые почвы с резко выраженным переменным режимом увлажнения.
Хорошо развитые глубокие дренированные почвы с pH 5.9–6.2 развиваются под мезофитными лианово-разнокустарниковыми широколиственно-кедровыми лесами и их производными сообществами (п.п. 6-1975, секц. 2).
Под ксеромезофитным кедровником с дубом (п.п. 12-1976, секц. 2) и кедровником с темнохвойными (п.п. 3-1974; 2-1986) и их производными сообществами развиваются маломощные хорошо дренированные свежие, периодически сухие почвы с непостоянным увлажнением и pH 6.1–6.3. Мощность гумусового горизонта изменяется в ходе демутационных сукцессий от незначительной на фрагментарных пятнах между обломками горной породы на первых этапах послепожарных сукцессий (п.п. 36-1983) до 12 см в коренных лесах (п.п. 3-1974; 2-1986) (Sapozhnikov, Gavrenkov, 1980).
Для всех бурых горно-лесных почв характерно наличие большого количества копролитов, образующихся после прохождения почвы через кишечник червей и придающие почве зернистую структуру. Это указывает на наличие в этих почвах большого количества червей. Известна огромная роль дождевых червей, участвующих в образовании почвенного плодородия в результате измельчения растительных остатков, минерализации органического материала и обогащения почвы гумусом, обеспечении дренажа и вентиляции почвы благодаря своим многочисленным ходам (Chekanovskaya, 1960; Kaydun, 2018).
Степень воздействия пожаров на подстилки и почву зависит от их интенсивности и характера лесорастительных условий. Все исследуемые нами послепожарные биогеоценозы образовались после устойчивых низовых пожаров. В наибольшей степени воздействие огня происходит в местообитаниях с явным дефицитом влаги в почве. В мезоксерофитных кедровниках и их производных лесах интенсивное горение обычно распространяется по всему напочвенному покрову и выгорает почти весь растительный покров и подстилка, сильно повреждается гумусовый горизонт с образованием “корки спекания” (Sapozhnikov, Kostenkova, 1984). В свежих и влажных экотопах характерно мозаичное выгорание. На молодых гарях можно выделить микробиотопы со слабой, умеренной, сильной и очень сильной степенью выгорания растительности и подстилки.
Для каждого типа послепожарных микробиотопов характерны определенные комплексы почвенно-гидрологических, микробиологических и фитоценотических условий. После воздействия огня слабой интенсивности обычно уничтожаются рыхлый сухой опад, ветошь и отдельные растения. При этом несколько улучшаются физические, химические и микробиологические свойства почвы. Под влиянием огня умеренной интенсивности повышается азотофиксирующая активность почвы, возрастает pH, увеличивается содержание водорастворимых минеральных соединений. При сильной интенсивности горения отмечается временная избыточность щелочности и высокая концентрация легкорастворимых минеральных соединений. При очень сильной интенсивности горения ухудшаются физические свойства почвы, уменьшается их порозность, разрушается структура и происходит спекание почв (Sapozhnikov, Kostenkova, 1984).
На однолетней гари кедровника с дубом (п.п. 36-1983) в микробиотопах со слабой степенью выгорания средний показатель pH водной вытяжки составил 5.4, а в микробиотопах с очень сильной степенью выгорания возрос до 7.4. Результаты химического анализа почвенных образцов, взятых на однолетней гари в “корковом слое спекания” и в слабо нарушенных микробиотопах, проведенный Т.М. Ильиной (Komarova, Il`ina, 2002), приведены в табл. 2.
Таблица 1.
Общие сведения о местах сбора почвенных образцов в послепожарных биогеоценозах Table 1. General information on the sites of soil samples collection in post-fire biogeocenoses
| Лет после пожара Years after fire |
№ пробной площади (п.п.) № of sample plot |
Местоположение, экспо-зиция и крутизна склона Location, exposure and slope steepness |
Типы сукцессионных рядов Succession series types |
|---|---|---|---|
| Тепло-сухие дубово-кедровые леса Warm-dry oak-Korean-pine forests | |||
| 70 | 30-1982 | Верхняя часть крутого ЮЗ склона; 630–650 м над ур. м. Upper part of steep SW slope; 630–650 m a.s.l. | Мезоксерофитный осиново-березовый редкопокровный лес на месте дубово-кедрового редкопокровного леса/Mesoxerophytic Populeto-Betuletum sparse forest on the site of Pinus koraiensis + Quercus mongolica sparse forest |
| Умеренно тепло-свежие широколиственно-кедровые леса Moderately warm and mesophytic broadleaved-Korean-pine fresh forests |
|||
| 1 | 36-1983 | Верхняя часть крутого ЮЗ склона; 620–640 м над ур. м. Upper part of steep SW slope; 620–640 m a.s.l. | Однолетняя гарь на месте ксеромезофитного кедровника с дубом лимонниково-лещинного разнотравно-мелкоосокового/Annual burnt site of xeromesophytic Pinus koraiensis + Quercus mongolicaSchisandra chinen- sis + Corylus mandshurica herbal-shallow sedge forest |
| 9 | 6-1975 секц. 2 section 2 |
Средняя часть З склона средней крутизны (20–25°); 560–580 м над ур. м. Middle part of W slope, medium steepness (20–25°); 560–580 m a.s.l. |
Осиново-березовый молодняк на месте мезофитного широколиственно-кедрового лианово-разнокустарникового василистниково-папоротниково-осокового леса/Young Populeto-Betuletum stands on the site of a mesophytic broadleaved-Pinus koraiensis forest with lianas, different shrubs and meadow-rue, ferns and sedges |
| 45 | 8-1982 | Средняя часть крутого ЮЗ склона; 450–470 м над ур. м., 15 км южнее стационара Middle part of steep SW slope; 450–470 m a.s.l., 15 km S of the station | Осиново-березовый лес на месте ксеромезофитного лимонниково-лещинного разнотравно-мелкоосокового кедровника с дубом/Populeto-Betuletum on the site of a xeromesophytic Pinus koraiensis with Quercus mongolica forest with Schisandra chinensis + Corylus mandshurica and herbal-shallow sedges |
| 200 | 3-1974 | Средняя часть крутого ЮЗ склона; 650–670 м над ур. м. Middle part of the steep SW slope; 650–670 m a.s.l. |
Ксеромезофитный кедровник с темнохвойными породами лимонниково-лещинный низкотравно-мелкоосоковый/Xeromesophytic Pinus koraiensis forest with dark coniferous species, Schisandra chinensis + Corylus mandshurica and low-grass-shallow sedge |
| 210–220 | 2-1986 | Верхняя часть крутого ЮЗ склона; 670–690 м над ур. м. Upper part of steep SW slope; 670–690 m a.s.l. |
Ксеромезофитный кедровник с темнохвойными породами лимонниково-лещинный низкотравно-мелкоосоковый/Xeromesophytic Pinus koraiensis forest with dark-coniferous species, Schisandrachinensis + Corylus mandshurica and low-grass-shallow sedge |
| >350 | 12-1976 секц. 2 Section 2 |
Средняя часть крутого З склона; 560–580 м над ур. м. Middle part of steep W slope; 560–580 m a.s.l. | Перестойный ксеромезофитный кедровник с дубом лимонниково-лещинный разнотравно-мелкоосоковый/Overmatured xeromesophytic Pinus koraiensis forest with Quercus mongolica, Schisandra chinensis + Corylus mandshurica and herbal-shallow sedge |
Таблица 2.
Валовое содержание зольных элементов в образцах, взятых в микробиотопах с очень сильным и слабым выгоранием (в % на абс. сух. вещество) (Komarova, Il’ina, 2002) Table 2. Total content of ash elements in samples taken in microbiotopes with very strong and weak burnt (% of absolute dry matter) (Komarova, Il’ina, 2002)
| Характер микробиотопов Microbiotopes nature |
Чистая зола, % Pure ash,% |
Si | Fe | Al | Ca | Mg | P | K | Na |
|---|---|---|---|---|---|---|---|---|---|
| “Корковый слой спекания”, n = 8 “Sintering crust”, n = 8 |
16.09 | 1.38 | 0.88 | 1.24 | 4.53 | 0.58 | 0.14 | 0.28 | 0.05 |
| Сохранившиеся подстилка и почва, n = 8 Preserved litter and soil, n = 8 |
4.11 | 0.81 | 0.59 | 0.47 | 1.74 | 0.38 | 0.16 | 0.2 | 0.02 |
Как следует из показателей этой таблицы, процентное содержание зольных элементов в чистой золе “коркового слоя спекания” значительно выше их содержания в слабо нарушенных микробиотопах. Наиболее резко возросло содержание кальция и алюминия. При этом известно, что алюминий является наиболее токсичным элементом для прорастания семян растений и особенно хвойных пород.
Динамику послепожарной трансформации подстилки и почвы в разных микробиотопах после устойчивого низового пожара в мезофитном широколиственно-кедровом лесу (п.п. 6-1975. секц. 2) в течение 7 лет наблюдали А.П. Сапожников и А.Ф. Костенкова (Sapozhnikov, Kostenkova, 1984). Микробиотопы с очень сильной степенью горения с образованием “коркового слоя спекания” они отнесли к органо-деструктивному типу повреждения. Гумусовый горизонт при этом не претерпевал внешних морфологических изменений, но заметно обогащался примесью углистых частиц. Через три года после пожара “корковый слой спекания” трансформировался в пиромуллевый подгоризонт подстилки. Затем этот подгоризонт сменился пирогумусным подгоризонтом, и через 7 лет после пожара подстилка утратила черты пирогенеза и восстановила свое первоначальное строение. Показатели pH вытяжки подстилок и верхнего слоя почвы в микробиотопах с разной интенсивностью выгорания выровнялись. На основании проведенных исследований был сделан вывод, что устойчивые низовые пожары в широколиственно-кедровых лесах практически не затрагивают минеральную часть почвы, а вызывают травматизм подстилок, которые в довольно короткий период (до 7–10 лет) проходят путь трансформации и восстановления (Sapozhnikov, Kostenkova, 1984; Soil formation, 1993).
В опубликованной коллективной монографии (Komarova, et al., 2017) дана детальная характеристика состава, строения и динамики растительных сообществ, а также почвы и условий произрастания всех анализируемых в данной работе сукцессионных рядов.
МАТЕРИАЛЫ
Отбор почвенных образцов проводился в биогеоценозах, находящихся на разных стадиях послепожарных демутационных сукцессий. В связи с этим в качестве основной классификационной единицы был принят тип сукцессионных рядов, включающий биогеоценозы на всех стадиях послепожарных лесовосстановительных смен, свойственных данному типу условий местопроизрастания (Komarova, 1992).
Почвенные образцы были взяты с 7 пробных площадей (п.п.) 50 × 50 м, растительные сообщества которых принадлежат 7 типам послепожарных сукцессионных рядов, относящихся к 2 экологическим комплексам, выделенным на основе близости условий местопроизрастания по режимам прямодействующих факторов – увлажнению и теплу (табл. 1).
Давность пожара устанавливалась по возрасту деревьев верхнего полога древостоя, у которых были взяты керны с помощью бура Пресслера, а у модельных деревьев брали спилы. О послепожарном происхождении фитоценозов судили также по наличию скопления углей в разных слоях почвы.
Почвенные образцы были взяты также в осиново-березовом молодняке, который образовался на 9 год после экспериментальной сплошной рубки перестойного лимонниково-лещинного кедровника с дубом (п.п. 12-1976, секц. 1). ПБС на 9-летней вырубке изучался с целью сравнительного анализа с ПБС одновозрастного осиново-березового молодняка (п.п. 6-1975, секц. 2), сформировавшегося после устойчивого низового пожара лианово-разнокустарникового широколиственно-кедрового леса. Оба участка расположены в средней части западного склона около 560–580 м над ур. м. Вместе с тем, возраст допожарного древостоя на п.п. 6-1975, секц. 2 не превышал 160–180 лет, а экспериментальная рубка (п.п. 12-1976, секц. 1) была проведена в перестойном лесу, где большинство крупных деревьев Pinus koraiensis были с гнилой сердцевиной; возраст единичных деревьев с сохранившейся стволовой сердцевиной превышал 350 лет, одно дерево имело максимальный возраст 450 лет.
МЕТОДЫ
Существуют разные методы и способы изучения почвенных запасов семян, которые детально рассмотрены в работах Т.А. Работнова (Rabotnov, 1958; 1982). Для изучения ПБС мы использовали метод “проращивания семян в почве” (Rabotnov, 1982). Минусы и плюсы этого метода были отмечены разными авторами (Petrov, 1989; Lyanguzova, 2002; Ivanova, 2004).
На всех исследуемых участках мы выбирали по 15–20 точек, расположенных регулярно по диагоналям пробных площадей (п.п.), и вырезали там монолиты размером 20 × 20 см из подстилки и гумусового горизонта почвы. Для установления вертикальной структуры ПБС образцы брали послойно из разных подгоризонтов подстилки (О1, О2 и О3) и верхних минеральных горизонтов почвы через каждые 2 см. Исследовали семена и односемянные плоды. Для проращивания семян были использованы пластиковые кюветы, размером 20 × 20 × 6 см, дно которых покрывалось слоем прокаленного песка 2–3 см, а сверху размещались образцы из разных подгоризонтов подстилки и почвы. Почвенные образцы содержали в специально оборудованном неотапливаемом лабораторном корпусе на Верхнеуссурийском стационаре в условиях рассеянного и достаточного света. Образцы поддерживали во влажном состоянии (около 60%). В зимний период полив не производился.
Наблюдения за прорастанием семян в образцах, взятых на 9-летней гари широколиственно-кедрового леса (п.п. 6-1975, секц. 2), начали проводить 8 июля 1982 г. и продолжали в течение 5 лет. На 7 остальных участках исследования проводились в последующие годы с середины мая до конца сентября со средней продолжительностью наблюдений за образцом 3–4 года. Это позволило установить характер и длительность выхода семян разных видов растений из покоя. О численности семян судили по числу появившихся всходов.
Появляющиеся проростки регулярно подсчитывались, результаты записывали в журнал; проводились морфологическое описание и зарисовка различных фаз развития. Проростки не установленных видов нумеровались и пересаживались в деревянные ящики с почвой или в открытый грунт для дальнейшего доращивания и уточнения видовой принадлежности. Краткая морфологическая и биологическая характеристика плодов, семян, проростков и их рисунки у 91 вида растений, появившихся в почвенных образцах, приведены в работе Т.А. Комаровой (Komarova, 1986). Кроме того, в этой книге дано морфологическое и биологическое описание гаметофитов и первых фаз развития спорофитов у 10 видов папоротников и представлены их рисунки.
Латинские названия растений в данной работе приведены по сводке “Сосудистые растения Советского Дальнего Востока” (Sosudistye, 1985–1996).
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
Начало прорастания семян в почвенных образцах тесно связано с температурой в лабораторном корпусе и характером участка, на котором были взяты образцы. Раньше и дружнее всех прорастали всхожие семена в образцах, взятых 8-го июля в 9-летнем пионерном кустарниково-осоково-разнотравном сообществе (п.п.6-1975, секц. 2). Первыми (на 5–6-й день) появились проростки Epilobium davuricum, Erigeron canadensis и Chelidonium asiaticum в подстилке и верхнем слое гумусового горизонта почвы. Средняя суточная температура при этом составляла 14°С. Массовое прорастание их семян началось на 10–12 день от начала полива образцов, и через 20 дней активное прорастание семян уже закончилось у Epilobium davuricum. Семена у Chelidonium asiaticum активно прорастали до конца августа, а в сентябре проростки уже не появлялись. У большинства остальных видов семена в образцах с этого участка начали активно прорастать на 12–15 дни и по всему почвенному профилю. Позже всех (на 30 день) здесь начали прорастать семена у Artemisia rubripes в верхнем слое гумусового горизонта почвы.
В почвенных образцах, взятых в середине мая последующих лет с других участков, прорастание семян начиналось обычно в подстилке или верхнем слое гумусового горизонта почвы на 13–15 день от начала полива при средней суточной температуре 8–10°С. При этом семена одних и тех же видов, взятых в различных биогеоценозах, начинали прорастать в разное время. Так, семена Chelidonium asiaticum начали прорастать в образцах, взятых в 45-летнем осиново-березовом лесу (п.п. 8-1982), на 12 день, в 70-летнем осиново-березовом лесу (п.п. 30-1982) – на 15 день, на однолетней гари кедровника с дубом (п.п. 36-1983) – на 21 день, а в коренных кедровниках с темнохвойными (п.п. 3-1974 и 2-1986) – только на 33 и 35 день.
Результаты нашего проращивания семян в течение пяти лет в почвенных образцах, взятых под 9-летним осиново-березовым молодняком, приведены в табл. 3 для видов, прораставших более двух лет. На 4-й год проводили процесс стимуляции прорастания семян путем повторного высушивания и намачивания почвенных образцов, а также полива слабым раствором KNO3. Это вызвало у Epilobium davuricum, Erigeron сanadensis и Philadelphus tenuifolius некоторое увеличение числа проросших семян в этот же год по сравнению с предыдущим годом, а у Artemisia rubripes – на следующий 5-й год наблюдений. Надо отметить, что 84–90% всех семян проросло в течение первых двух лет. Вероятно, этим обуславливается принятая другими исследователями двухлетняя продолжительность наблюдений за прорастанием семян. Вместе с тем, и более длительные наблюдения не дают полного представления обо всем составе жизнеспособных семян в подстилках и почве из-за ограниченной суммарной площади почвенных образцов (не более 1 м2). Кроме того, жизнеспособные семена у некоторых видов растений требуют дополнительную химическую или термическую обработку.
Таблица 3.
Динамика ежегодного прорастания всхожих семян (экз./м2) на п.п. 6-1975, секц. 2 (n = 20 для каждого года) Table 3. Dynamics of annual germination of viable seeds/m2 on sample plot 6-1975, section 2 (n = 20 for each year)
| Вид растений/ Plant species |
Годы проращивания/Years of germination | Всего Total |
||||
|---|---|---|---|---|---|---|
| 1-й | 2-й | 3-й | 4-й | 5-й | ||
| Artemisia rubripes | 85 | 116 | 15 | 8 | 15 | 239 |
| Erigeron canadensis | 391 | 592 | 146 | 210 | 6 | 1345 |
| Epilobium davuricum | 167 | 78 | 14 | 114 | – | 373 |
| Philadelphus tenuifolius | 146 | 423 | 26 | 49 | – | 644 |
| Chelidonium asiaticum | 2306 | 210 | 52 | 6 | – | 2604 |
| Carex campylorhina | 398 | 49 | 8 | – | – | 455 |
В отечественной и зарубежной литературе были предложены различные классификационные схемы для почвенных банков семян. За рубежом первую классификационную схему для почвенных банков семян предложили D.E. Schafer и D.O. Chilcote (1969), а затем еще были разработаны 9 классификационных систем (Thompson, Grime, 1979; Grime, 1981; и др.). В эти системы входили от 3 до 12 категорий, включая длительность жизни семян, устойчивость или постоянство в семенных банках, вес, размеры и форма семян, способы их распространения и др. Наибольшее признание в зарубежной литературе получила система классификации, предложенная К. Thompson et al. (1979, 1993). В этой системе выделены три типа почвенных банков семян: 1) временные банки семян; 2) краткосрочные постоянные (persistent) семенные банки; 3) долгосрочные постоянные банки семян. Т.А. Работнов (Rabot-nov, 1982) выделил 3 типа: 1) автохтонные семена, поступившие в результате обсеменения растений, входящих в состав данного сообщества; 2) реликтовые семена, входившие в состав сообществ, существовавших в данном месте в прошлом; 3) инвазионные семена, принесенные извне. Сравнительный анализ всех предложенных классификационных схем провели ряд авторов (Csontos and Tamás, 2003; Yu et al., 2007).
Составленная нами система ПБС близка к классификационным схемам Т.А. Работнова (Rabotnov, 1982) и К. Thompson et al. (1993), но в ее основу был взят динамический аспект применительно к сукцессионным рядам. По длительности пребывания и характеру участия всхожих семян в ПБС послепожарных сукцессионных рядов все виды мы распределили на 3 группы, у которых всхожие семена: 1) всходят в ПБС в течение короткого времени после обсеменения растений и не накапливаются в почве; 2) более или менее постоянно присутствуют в ПБС сукцессионных рядов за счет периодического поступления извне и относительно длительного сохранения всхожести в почве; 3) постоянно присутствуют в ПБС сукцессионных рядов в результате массового поступления на ранних стадиях сукцессий и длительного покоя при погребении в почве (Komarova, 1986).
К первой группе относятся все лесообразующие древесные породы коренных сообществ (Pinus koraiensis, Picea ajanensis, Quercus mongolica, Tilia taquetii и др.), а также все представители семейства Salicaceae (Salix caprea, S. taraikensis, Po-pulus tremula, P. koreana). Кроме того, к этой группе относятся многие травянистые анемохоры (Chamerion angustifolium, Picris koreana, Aster ageratoides, Lactuca triangulata).
Семена у представителей второй группы могут сохранять жизнеспособность в почве довольно длительное время и постоянно пополняют ПБС за счет распространения семян с помощью ветра и других агентов. К этой группе относятся семена древесно-кустарниковых пород: Aralia elata, Sambucus racemosa, Philadelphus tenuifolius, виды рода Betula (B. costata, B. lanata, B. platyphylla), а также многие представители семейств Cyperaceae (Carex campylorhina, C. xyphium, C. reventa, C. ussuriensis и др.), Poaceaeа (Cinna latifolia, Neomolinia mandshu-rica, Melica nutans и др.), однолетние и двулетние растения (Erigeron canadensis, Epilobium davuricum, Impatiens noli-tangere), жизнеспособные семена которых содержатся не только в подстилке и верхнем слое почвы, но и в более глубоких слоях гумусового горизонта почвы.
Третью группу образуют растения, семена которых поступают в большом количестве в первые годы после пожара и длительное время находятся в покоящемся состоянии погребенными в почве, пока не создадутся благоприятные условия для их прорастания. В эту группу входят раннесукцессионные травянистые растения (Chelidonium asiaticum, Artemisia rubripes, Lamium barbatum, Urtica angustifolia). Семена их рассеиваются преимущественно барохорно вблизи материнского растения, что в сочетании с длительным покоем семян обеспечивает им устойчивое положение во времени на одних и тех же участках.
Видовой состав всхожих семян в ПБС биогеоценозов отдельных сукцессионных рядов изменяется от 19 до 47 видов, а общая численность всхожих семян варьирует от 0.95 до 7.3 тыс. экз./м–2. Состав и численность всхожих семян в 6 послепожарных сукцессионных рядах приведены в опубликованных работах (Komarova, 1986, 1992).
Как показали результаты исследований, видовой состав и численность всхожих семян существенно изменяются в зависимости от положения биогеоценозов в сукцессионных рядах (рис. 1).
Рис. 1.
Изменения запасов всхожих семян (экз./м2) в подстилке и почве в ходе послепожарного восстановления широколиственно-кедровых лесов. Легенда: подгоризонты подстилки (О1, О2, О3) и двухсантиметровых слоев почвенного горизонта (AY). Ось Х – количество семян (экз./кв.м). Ось Y – возраст сообщества после пожара (лет). Fig. 1. Changes in viable seeds/m2 in the litter and soil during the post-fire restoration of broadleaved-Korean-pine forests. Legend: subhorizons of litter (O1, O2, O3) and two-centimeter layers of the soil horizon (AY). Х-axis – number of seeds/m2. Y-axis – age after fire (years).
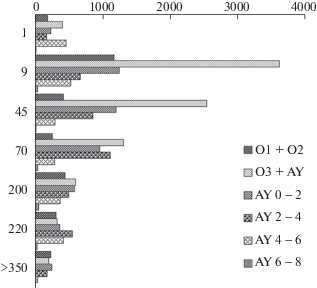
Наибольшие показатели численности и видового разнообразия всхожих семян отмечены в ПБС 9-летнего послепожарного осиново-березового молодняка. При этом максимальная концентрация семян была сосредоточена в нижнем слое подстилки (O3). Сформировавшаяся после пожара подстилка способствовала накоплению и консервации в приповерхностном слое почвы жизнеспособных семян растений, активно плодоносивших в первые годы после пожара. В наибольшем числе здесь были представлены семена Chelidonium asiaticum, Erigeron canadensis, Epilobium davuricum, Artemisia rubripes, Lamium barbatum, Sonchus arvensis, Urtica angustifolia и других раннесукцессионных видов, плодоносящие растения которых отсутствовали в растительном покрове.
В ПБС 45- и 70-летних осиново-березовых лесов, образовавшихся на месте ксеромезофитных кедровников с дубом, общая численность всхожих семян уменьшилась и особенно резко во всех слоях подстилки, увеличение численности семян отмечалось только в слое гумусового горизонта AY2-4. Всхожие семена новых видов, по сравнению с ПБС 9-летнего осиново-березового молодняка, не были обнаружены. Последовательное уменьшение численности всхожих семян и перемещение зоны их максимальной концентрации из подстилки и верхнего слоя гумусового горизонта почвы в более глубокие слои почвы характерно и в дальнейшем ходе послепожарных демутационных сукцессий. Постепенное заглубление семян связано с ежегодным накоплением на поверхности почвы огромного количества отмершего органического материала, который постепенно гумифицируется и минерализуется с помощью почвенной фауны и микрофлоры, включающихся в процесс почвообразования. Некоторые авторы (Petrov, 1989; Regnier et al., 2008; Eisenhauer et al., 2009; и др.) отмечают важную роль дождевых червей в перемещении семян с поверхности вглубь почвы и увеличении числа покоящихся семян из-за их погребения.
Для ПБС в биогеоценозах на разных стадиях послепожарных сукцессий и в различных типах сукцессионных рядов характерны свои “верные” виды (Petrov, 1989; Ivanova, 2004); в зарубежной литературе им соответствует термин “persistent” (постоянные) виды. На очень ограниченный состав в почвенных банках “верных” (постоянных) доминирующих по числу семян видов указывали разные исследователи (Petrov, 1989; Ivanova, 2004; Erenler еt al., 2010). Согласно исследованиям Erenler H.E. и др. (2010), из 81 установленного вида в почвенных банках семян на четырех участках искусственных и естественных лесов центральной Англии, к постоянным доминантам относился лишь один вид – Juncus effusus, на долю семян которого приходилось 80% от общего числа семян.
По мнению ряда исследователей (Holzel et al., 2004; Wellstein et al., 2007) к постоянным доминантам ПБС относятся преимущественно однолетние и малолетние растения с длительностью онтогенеза не более 5 лет. Семена их преимущественно мелкие, легко проникающие вглубь почвы. По мнению S.L. Ju et al. (2007) такие виды растений с мелкими и легкими семенами относятся к группе постоянных доминантов в регионах с влажным климатом, тогда как в районах с сухим климатом к группе постоянных доминантов обычно относятся растения с крупными семенами.
Как следует из показателей табл. 4 в ПБС всех производных сообществ широколиственно-кедровых лесов максимально представлены всхожие семена раннесукцессионного вида Chelidonium asiaticum. Род Chelidonium долгое время считался монотипным, представленным одним видом Chelidonium majus L. На основании кариологических различий A. Krahulcova (1982) предложила различать два вида: Chelidonium majus c числом хромосом 2n = 12, распространенный в Европе и Азии, и Chelidonium asiaticum (Hara) Krahulcova с числом хромосом 2n = 10, встречающийся в Восточной Азии и Японии. Несмотря на сходные биологические и морфологические особенности у этих видов, произрастают они в разных условиях местообитания. Четкую экологическую дифференциацию у этих видов отметили также В.П. Селедец и Н.С. Пробатова (Seledets, Probatova, 2018). Всхожие семена Chelidonium majus редко встречаются в лесных почвах. Сравнительно высокое содержание всхожих семян этого вида было отмечено под искусственными белоакациевыми насаждениями (Al`bitskaya, 1965) и искусственными ясеневыми насаждениями Днепровщины (Al`bitskaya, Moroz, 1965), где растения Chelidonium majus господствовали в травяном покрове.
Таблица 4.
Характеристика основных “верных” доминантов в ПБС исследуемых биогеоценозов (n = 20 для каждой п.п.) Table 4. Characterization of the main persistent dominants in the soil seed bank of the studied biogeocenoses (n = 20 for each plot)
| Лет после пожара Years after fire |
№ пробной площади (п.п.) № of sample plot |
Общее число семян, экз./м2 Total number of seeds/m2 |
Число видов Number of species |
Постоянные доминирую-щие виды по числу семян Permanent dominant species by number of seeds |
Число семян, экз./м2 Number of seeds/m2 |
% от всех семян % of the total number of seeds |
|---|---|---|---|---|---|---|
| 1 | 36-1983 | 1512 | 41 | Chelidonium asiaticum | 214 | 14.2 |
| Philadelphus tenuifolius | 208 | 13.7 | ||||
| 9 | 6-1975, секц. 2 section 2 |
7306 | 47 | Chelidonium asiaticum | 2604 | 35.6 |
| 45 | 8-1982 | 5361 | 33 | Chelidonium asiaticum | 1948 | 36.3 |
| Erigeron сanadensis | 1758 | 32.7 | ||||
| 70 | 30-1982 | 4008 | 35 | Chelidonium asiaticum | 1358 | 33.9 |
| 200 | 3-1974 | 2593 | 33 | Betula platyphylla + B. costata | 528 | 20.4 |
| Carex campylorhina | 515 | 19.8 | ||||
| 220 | 2-1986 | 2057 | 32 | Betula platyphylla + B. costata | 315 | 15.3 |
| Carex campylorhina | 192 | 9.3 | ||||
| Philadelphus tenuifolius | 173 | 8.4 | ||||
| Более 350 > 350 |
12-1976, секц. 2 section 2 |
946 | 19 | Betula platyphylla + B. costata | 219 | 23.1 |
| Philadelphus tenuifolius | 100 | 11 | ||||
| Carex campylorhina | 90 | 10.5 |
В горных кедрово-широколиственных лесах Южного и Среднего Сихотэ-Алиня растениям Chelidonium asiaticum принадлежит одно из ведущих мест в формировании послепожарных сообществ, благодаря большим запасам всхожих семян в лесных почвах (Комарова, 1984). После устойчивых низовых пожаров зарастание освободившихся участков в течение первых трех лет идет чаще всего через тип чистотеловых гарей. Массовое появление всходов этого вида обычно отмечается только в первые два года после пожара. В течение 1-го года его растения проходят виргинильный период и в середине июля формируют первичные репродуктивные побеги до 50–60 см выс. В конце августа – начале сентября происходит массовое осыпание семян. Коробочки Chelidonium asiaticum обладают слабо выраженным разбрасывающим механизмом и семена опадают вблизи материнских растений. Автобарохорное осыпание семян сочетается с распространением муравьями, которых привлекают мягкие и сочные придатки на семенах, но разносят они семена не более, чем на 2–3 м. На 2-й год большая часть его растений представлена средневозрастными генеративными особями, достигающими максимальных размеров и продуктивности. Они обычно обильно плодоносят и обсеменяют территорию, формируя новый многочисленный почвенный банк семян. На 3-й год большинство растений Chelidonium asiaticum переходит в онтогенетическое состояние старых генеративных особей, а на 4-й год – сенильных растений. Второе поколение у этого вида обычно отсутствует или представлено единичными ослабленными растениями, которые чаще всего погибают на стадиях проростков или ювенильных особей. Сильное тормозящее влияние на прорастание семян и рост молодых растений Chelidonium asiaticum оказывает токсичность колинов из их же надземных и подземных частей (Komarova, 1986). При этом токсичное влияние ветоши и подстилки из сухих остатков растений Chelidonium asiaticum сохраняется в подстилке и верхних слоях почвы достаточно длительное время. Там, где росли растения Chelidonium asiaticum, нами не были встречены молодые растения этого вида. В то же время небольшая концентрация водных экстрактов из надземных частей Chelidonium asiaticum (1–2%) может даже стимулировать прорастание семян и рост молодых растений у некоторых видов (Picea ajanensis, Populus koreana, Salix schwerinii и др.), но увеличение концентрации водных экстрактов до 5% подавляло прорастание семян у исследуемых древесных видов (Komarova, 1986).
В ПБС 9-летнего осиново-березового молодняка всхожие семена Chelidonium asiaticum значительно превосходили по численности все остальные виды и их максимум находился в уплотненном переходном горизонте (О3AY). При этом горизонтальное распределение его семян в этом переходном слое отличалось четкой мозаичностью. Количество его всхожих семян варьировало в образцах переходного горизонта от 0 до 105 семян на 20 площадках (20 × 20 см), представляющих регулярную выборку через 5 м по диагоналям участка. Мозаичность в горизонтальном распределении почвенных запасов семян Chelidonium asiaticum отражала мозаичный характер размещения синузий с преобладанием его растений на двухлетней гари этого участка (Komarova, 1992). Наибольшее число семян этого вида отмечено в почвенных образцах, взятых в микробиотопах с сильным и очень сильным выгоранием, на которых в первые три–четыре года доминировали растения Chelidonium asiaticum. На 2-й год после пожара синузии с доминированием этого вида занимали около 70% от общей площади гари, образовавшейся на месте мезофитного широколиственно-кедрового леса (п.п. 6-1975, секц.2), 56% – на гари ксеромезофитного кедровника с дубом (п.п. 36-1983) и около 85% – на гари мезоксерофитного лещинно-рододендронового кедровника с дубом (п.п. 42-1984). Вероятно, распределение семян этого вида по площади может дать определенное представление о будущем размещении его растений после очередного пожара.
В распределении всхожих семян Chelidonium asiaticum по вертикальному почвенному профилю для ПБС каждого участка характерны свои зоны максимальной концентрации (рис. 2), причем расположение их в почвенном профиле связано с размещением сообществ в сукцессионном ряду.
Рис. 2.
Распределение всхожих семян Chelidonium asiaticum (экз./м2) в подстилке и почве биогеоценозов на разных стадиях послепожарных демутационных сукцессий. Легенда: подгоризонты подстилки (О1, О2, О3) и двухсантиметровых слоев почвенного горизонта (AY). Ось Х – количество семян, экз./кв.м. Ось Y – возраст сообщества после пожара (лет). Fig. 2. Distribution of Chelidonium asiaticum viable seeds/m2 in the litter and soil of biogeocenoses at different stages of post-fire demutation successions. Legend: subhorizons of litter (O1, O2, O3) and two-centimeter layers of the soil horizon (AY). Х-axis – number of seeds/m2. Y‑axis – age after fire (years).
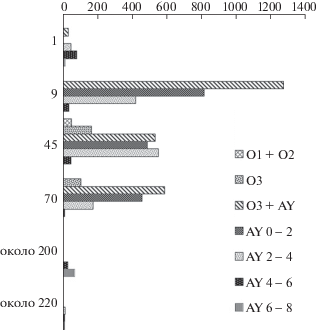
В подстилке, сформировавшейся после пройденных пожаров, семена его полностью отсутствовали. На однолетней гари (п.п. 36-1983) расположение всхожих семян Chelidonium asiaticum обусловлено главным образом распределением их в допожарном сообществе и неоднородностью выгорания подстилки и почвы. В верхнем слое почвы (0–2 см) семена его находились преимущественно в микробиотопах с сильной степенью выгорания. Они составили активный запас семян, согласно Т.А. Работнову (Rabotnov, 1983), находящихся в слое прорастания. В более глубоких слоях почвы (4–8 см) семена Chelidonium asiaticum содержались преимущественно в образцах, взятых в микробиотопах со слабой степенью выгорания. Они составили потенциальный запас семян, лежащих на недоступной для прорастания глубине (Rabotnov, 1983). В ходе демутационных сукцессий от пионерных сообществ до сомкнутых коренных лесов зоны максимальной концентрации всхожих семян Chelidonium asiaticum постепенно перемещаются в более глубокие слои почвы, что согласуется с заключением некоторых исследователей (Petrov, 1989; Petrov, Belyaeva, 1981): чем глубже залегают семена, тем дольше они покоятся в почве.
Почвенный запас семян на однолетней гари кедровника с дубом состоял преимущественно из семян, сохранившихся в почве во время пожара и занесенных семян с соседних участков (Betula platyphylla, B. costata, Populus tremula, Carex campylorhina), поэтому в нем не выделялись преобладающие по числу семян виды. Незначительно по численности преобладали всхожие семена Chelidonium asiaticum и широко распространенного в лесах и на нарушенных участках кустарника Phi-ladelphus tenuifolius. Растения этого кустарника обильно цветут и плодоносят только на освещенных участках. Уже в первый год после пожара его сохранившиеся растения начинают цвести и плодоносить. При созревании семян коробочки широко раскрываются, и легкие семена разносятся ветром или рассыпаются при раскачивании их пружинящих плодоножек. Всхожие семена Philadelphus tenuifolius присутствуют по всему почвенному профилю на всех стадиях послепожарных сукцессий в широколиственно-кедровых лесах. Также широко и по всему почвенному профилю представлены всхожие семена у другого крупного кустарника – Sambucus racemosa и деревянистой лианы – Actinidia kolomikta. Семена у них распространяются эндозоохорно и постоянно пополняют ПБС во всех сообществах.
В ПБС 45-летнего (п.п. 8-1983) и 70-летнего (п.п. 30-1982) послепожарных осиново-березовых лесов наибольшее количество семян также принадлежало Chelidonium asiaticum, а зоны его максимальной концентрации находились в верхнем слое гумусового горизонта почвы.
В ПБС на всех этапах послепожарных сукцессий присутствуют семена других постоянных, но не доминирующих, а сопутствующих раннесукцессионных видов (Erigeron сanadensis, Artemisia rubripes, Impatiens noli-tangere, Lamium barbatum, Sonchus arvensis, Urtica angustifolia и др.) и пионерных видов деревьев и кустарников (Betula рlatyphylla, Aralia elata, Sambucus racemosa). Всхожие семена их присутствовали в ПБС не только производных, но и коренных лесов, но численность в них была значительно меньше, чем у “верных” доминирующих видов.
Как было отмечено ранее рядом авторов (Karpov, 1969; Rabotnov, 1983), благоприятные условия для длительного сохранения всхожести семян создают развитая подстилка и высокое содержание гумуса в почве. Для проверки возможности содержания всхожих семян в более глубоких слоях почвы, не содержащих гумуса, мы взяли почвенные образцы из коренного кедровника с темнохвойными (п.п. 3-1974) с площадок размером 20 × 20 см в трехкратной повторности из иллювиального горизонта (В) на глубине 40–42 см и в желтовато-буром дресвянисто-каменистом горизонте коренной породы (С) на глубине 60–62 см от поверхности почвы. Наблюдения в течение трех летних месяцев за прорастанием семян в пластиковых кюветах показали полное отсутствие в них жизнеспособных семян.
Минимальный видовой состав (19 видов) и низкая численность всхожих семян (946 экз./м2) в климатическом комплексе широколиственно-кедровых лесов были отмечены в достаточно мощной подстилке и развитой почве с высоким содержанием гумуса в ПБС старовозрастного кедровника с дубом (п.п. 12-1976, секц. 2), возраст которого более 350 лет. Явно доминирующие виды по численности семян здесь отсутствовали, а наибольшим числом были представлены всхожие семена Viola selkirkii, Betula costata и B. рlatyphylla, Philadelphus tenuifolius и разных видов Carex (С. campylorhina, C. xyphium, C. reventa), растения которых присутствовали в растительном покрове этого леса. Раннесукцессионные виды были представлены здесь только 5 видами (Sambucus racemosa, Artemisia rubripes, Saussurea ussuriensis, Epilobium davuricum и Erigeron canadensis), общая численность которых составляла всего 114 экз./м2 и распределены они были преимущественно в нижнем подгоризонте подстилки и в верхних слоях гумусового горизонта почвы. Полностью здесь отсутствовали всхожие семена Chelidonium asiaticum. В коренных климаксовых кедровниках с темнохвойными породами (п.п. 3-1974 и 2-1986) всхожие семена Chelidonium asiaticum (с численностью всхожих семян 100 и 27 экз./м2 соответственно) были отмечены только в гумусовом горизонте на глубине 4–8 см. Отсутствие всхожих семян Chelidonium asiaticum в перестойном лесу (п.п. 12-1976, секц. 2), вероятно, объясняется тем, что долговечность его семян в почве не достигает 350 лет.
Ограниченный видовой состав всхожих семян, их низкая численность и незначительное участие в ПБС раннесукцессионных видов, возможно, связано с действием многочисленных фитотоксических веществ – антибиотиков, выделяемых плесневыми грибами, актиномицетами, водорослями и другими микроорганизмами, а также токсичными продуктами выделения надземных и подземных частей растений, которые обильно скапливаются в перестойных насаждениях. Все это приводит к известному явлению – почвенному утомлению (Bublitz, 1953; Grodzinskiy, 1965).
Для установления различий между банками семян на вырубке перестойного леса и участке после пожара 160–180-летнего широколиственно-кедрового леса был проведен сравнительный анализ видового состава и численности всхожих семян в образцах, взятых в одновозрастных осиново-березовых молодняках, образовавшихся 9 лет назад в результате устойчивого низового пожара широколиственно-кедрового леса (п.п. 6-1975, секц. 2) и после сплошной экспериментальной рубки перестойного кедровника с дубом (п.п. 12-1976, секц. 1). Расстояние между этими участками не превышает 1 км, и находятся они в сравнительно близких условиях местопроизрастания.
В результате 3-летних наблюдений за прорастанием всхожих семян в почвенных образцах, взятых на вырубке перестойного леса (п.п. 12-1976, секц. 1), выявлены 1428 экз./м2, относящихся к 29 видам. Все виды всхожих семян, отмеченные в ПБС перестойного леса, присутствовали и в ПБС 9-летней вырубки. Добавились преимущественно семена пионерных видов (Plantago major, Geum aleppicum, Gnaphalium uliginosum, Sonchus arvensis). Доминировали по числу семян два раннесукцессионных вида (Epilobium davuricum и Artemisia rubripes), совместная численность всхожих семян которых составляла 726 экз./м2, или 51% от общего числа проросших семян.
Семена Chelidonium asiaticum отсутствовали в ПБС этой вырубки. Вероятно, это связано с отсутствием его всхожих семян в ПБС перестойного леса. Как уже было отмечено ранее, семена этого автобарохорного вида не заносятся извне никакими агентами на значительные расстояния.
Низкую численность семян раннесукцессионных анемохорных видов (Erigeron canadensis, Sonchus arvensis, Lactuca triangulatа, Cirsium pendulum) в ПБС вырубки можно объяснить тем, что рубка производилась только с помощью бензопил без тяжелой техники, поэтому на значительной площади сохранилась подстилка, препятствующая расселению и прорастанию семян анемохорных растений.
Низкие показатели видового состава и численности всхожих семян в ПБС на вырубке (29 видов и 1.4 тыс. экз./м2) по сравнению с ПБС послепожарного осиново-березового молодняка (47 видов и 7.3 тыс. экз./м2), очевидно, связаны с различиями возрастных состояний биогеоценозов, бывших до пожара и рубки леса, а также стимулирующей ролью огня, о которой будет сказано ниже.
Роль отдельных факторов, вызывающих сокращение численности покоящихся в почве семян, анализировали многие исследователи (Taulorson, 1970; Fenner, Thompson, 2005). По их мнению, к основному фактору, приводящему к истощению численности семян, относится прерывание покоя и прорастание семян in situ, что сопровождается появлением всходов или чаще их гибелью. Снижение количества всхожих семян может быть связано с повреждением их патогенными грибами и почвенными беспозвоночными, а также с естественным процессом их старения. Причинами уменьшения численности ПБС в верхних частях крутых горных склонов служит скатывание семян по склону и смыв их водными потоками (Torgashkova, 2003; Morimoto et al., 2009), а мелкие гладкие семена сползают по многочисленным ходам дождевых червей вглубь почвы (Regnier et al., 2008; Eisenhauer et al., 2009).
ФАКТОРЫ, ВЛИЯЮЩИЕ НА ПОКОЙ И ПРОРАСТАНИЕ СЕМЯН ПОСЛЕ ПОЖАРА
С первых этапов послепожарных сукцессий вступают в действие особые механизмы, ограничивающие прорастание семян и обусловливающие накопление их в почве. Покой семян может наступать из-за недостатка влаги, света, кислорода, избытка углекислого газа и других причин. Прорастанию семян, содержащихся в почве, могут препятствовать густой травяной покров и мощный слой подстилки (Karpov, 1969; Rysin, Rysina, 1965; Torgashkova, 2003). Механическое препятствие, созданное почвой или лесной подстилкой, не дает прорастать семенам даже в случае их выхода из состояния вынужденного покоя (Torgashkova, 2003; Ivanova, 2004).
Благоприятные условия для массового прорастания покоящихся в почве семян создаются после пожаров и особенно велика роль устойчивых низовых пожаров. В результате их выгорает напочвенный покров, уничтожается подстилка, грубый гумус и обнажается поверхность почвы. При этом погребенные на глубине семена оказываются ближе к поверхности почвы, что благоприятствует их прорастанию.
Как известно, ход прорастания семян включает три необходимые фазы: набухания, стимуляции и роста (Lyr et al., 1974). Первая и необходимая фаза прорастания покоящихся в почве семян связана с поглощением определенного количества воды и зависит от количества почвенной влаги. После пожаров улучшаются условия почвенного увлажнения благодаря образованию открытых участков и свободному проникновению атмосферных осадков на поверхность почвы. Наши исследования на свежих гарях показали, что наиболее высоким содержанием влаги отличается верхний слой почвы, который представляет ложе для прорастающих семян (Komarova, 1986). Это обусловлено высокой влагоемкостью верхнего слоя гумусового горизонта бурых горно-лесных почв (Kostenkova, 1978).
Для второй фазы прорастания семян – стимуляции – важным условием служит выдерживание набухших семян при пониженных и переменных температурах. Это способствует более глубоким биохимическим превращениям, подготавливающим более быстрое прорастание семян (Nikolaeva, 1967). Наибольшие перепады температур характерны для оголенной поверхности почвы, лишенной подстилки и развитого напочвенного покрова. Согласно нашим наблюдениям, амплитуда суточных колебаний температуры воздуха на открытой поверхности двухлетней гари мезофитного широколиственно-кедрового леса (п.п. 6-1975, секц. 2), в середине июля достигала 20°С в то время, как под пологом сомкнутого древостоя (пр. пл. 6-1975, секц. 4) она не превышала 10°С.
Семена у раннесукцессионных видов обычно чувствительны к освещению и для их прорастания необходим свет. Уничтожение пожаром травяного покрова, подлеска, подроста и изреживание древесного полога способствуют проникновению прямого солнечного света к поверхности почвы и повышению ее освещенности.
Покоящиеся в почве семена обычно мелкие с небольшим запасом питательных веществ, поэтому их прорастание лучше происходит на плодородных почвах. Приток нитратов может действовать как “сигнал” для прорастания семян, а также как источник питания для молодых растений (Bazzaz, 1979). Вследствие уменьшения кислотности, повышения степени насыщенности основаниями, обогащения почвы элементами минерального питания, освободившихся от сгорания растений и подстилки происходит улучшение условий для прорастания семян и роста молодых растений.
Интенсивность роста семян зависит также от концентрации кислорода. Уничтожение огнем напочвенного покрова и мощного слоя подстилки уменьшает концентрацию углекислого газа и увеличивает доступ кислорода к поверхностным слоям почвы и содержащимся в них семенам.
Важную роль для прорастания семян, покоящихся в подстилке и почве, играет также устранение огнем фитотоксических веществ. Семена некоторых видов растений нуждаются в скарификации, поэтому обогрев во время пожара может стимулировать их прорастание.
ЗАКЛЮЧЕНИЕ
В результате многолетних исследований почвенных банков семян (ПБС) в ходе демутационных сукцессий после пожаров в широколиственно-кедровых лесах Южного Сихотэ-Алиня были установлены основные изменения в видовом составе, численности и вертикальном распределении всхожих семян в подстилке и почве, начиная с однолетних гарей до перестойного широколиственно-кедрового леса. Численность ПБС варьирует в пределах 0.95–7.3 тыс. семян на 1 м2, а видовой состав изменяется от 19 до 47 видов на разных этапах послепожарных сукцессий в широколиственно-кедровых лесах. В ходе сукцессии общая численность всхожих семян уменьшается с возрастом сообществ, а зона максимальной концентрация семян смещается из подстилки в более глубокие слои гумусового горизонта почвы.
Весь сложный комплекс прямых и косвенных воздействий огня – уничтожение напочвенного покрова и мощного слоя подстилки, в результате чего покоящиеся семена оказываются ближе к поверхности почвы, повышение на обнаженной поверхности почвы освещенности, влажности, амплитуды температурных колебаний, доступа кислорода, обогащение почвы элементами минерального питания, устранение действия фитотоксикантов, ослабление корневой конкуренции и т.д. – служит своеобразным “сигналом” для массового прорастания семян и активной вспышки семенного возобновления.
Список литературы
[Al`bitskaya] Альбицкая М.А. 1965. Засоренность почвы белоакациевых и дубовых искусственных насаждений зоны обыкновенного чернозема УССР (Комиссаровский лес). – Бот. журн. 50 (8): 1092–1104.
[Al`bitskaya, Moroz] Альбицкая М А., Мороз О.Б. 1965. О видовом составе и количестве семян в почве искусственных ясеневых насаждений на Днепровщине. – Бот. журн. 50 (6): 856–861.
Baskin C.C., Baskin J.M. 2001. Seeds. Ecology, biogeography, and evolution of dormancy and germination. Academic Press, San Diego. 680 p.
Bazzaz F.A. 1979. The physiological ecology of plant succession. – Annu. Rev. Ecol. Syst.. 10: 351–371.
Bossuyt B., Hermy M. 2001. Influence of land use history on seed banks in European temperate forest ecosystems: a review. – Ecography. 24: 225–238/
Bossuyt B., Heyn M., Hermy M. 2002. Seed bank and vegetation composition of forest stands of varying age in central Belgium: consequences for regeneration of ancient forest vegetation. – Plant Ecol.162: 33–48.
Bublitz W. 1953. Über den Einfluss von AuBenfaktoren auf die keimung und ihre Bedeutung fur die Naturverjungung. – Allg. Forstzeitschrift. 8 (53): 594–596.
Buhk C., Hensen I. 2005. Lack of hard-seeded species in pre-fire and post-fire seed banks in the region of Murcia (south-eastern Spain). – Anales de Biología. 27: 29–37.
[Chekanovskaya] Чекановская С.В. 1960. Дождевые черви и почвообразование. М. 212 с.
Csapody V. 1968. Keimlingsbestimmungsbuch der Dicotyledonen. Bp. Akad. Kiado. 286 p.
Csontos P., Tamás J. 2003. Comparisons of soil seed bank classification systems. – Seed Sci. Res. 13: 101–111.
Eisenhauer N., Straube D., Johnson E., Parkinson D., Scheu S. 2009. Exotic Ecosystem Engineers Change the Emergence of Plants from the Seed Bank of a Deciduous Forest. – Ecosystems. 12: 1008–1016.
Erenler H.E., Ashton P.A., Gillman M.P., Ollerton J. 2010. Factors determining species richness of soil seed banks in lowland ancient woodlands. – Biodivers. Conserv. 19: 1631–1648.
Fenner M., Thompson K. 2005. The ecology of seeds. Cambridge University Press. 260 p.
Ferrandis P., Herranz J.M., Martínez Sanchez J.J. 1996. The role of soil seed bank in the early stages of plant recovery after fire in a Pinus pinaster forest in SE Spain. – Int. J. Wildland Fire. 6 (1): 31–35.
Grandin U. 2001 Short-term and long-term variation in seed bank/vegetation relations along an environmental and successional gradient. – Ecography. 24: 731–741.
Grime J.P. 1981. The role of seed dormancy in vegetation dynamics. – Ann. Appl. Biol. 98: 555–558.
[Grodzinskiy] Гродзинский А.М. 1965. Аллелопатия в жизни растений и их сообществ. Основы химического взаимодействия растений. Киев. 200 с.
Hölzel N., Otte A. 2004. Interannual variations in the soil seed bank of flood meadows over two years with different flooding patterns. – Plant Ecol. 174: 279–291.
[Ivanova] Иванова Т.В. 2004. Методические аспекты изучения видового разнообразия почвенного банка семян. – Методы популяционной биологии. Сборник материалов VII Всероссийского популяционного семинара. Ч. 1. Сыктывкар. С. 90–92.
Juan T., Azcárate F.M., Peco B. 2006. The fate of seeds in Mediterranean soil seed banks in relation to their traits. – J. Veg. Sci. 17: 5–10.
[Kaydun] Кайдун П.И. 2018. Влияние дождевых червей на доступность растениям элементов минерального питания: азота, железа, цинка, марганца и кремния: Автореф. дис. … канд. биол. наук. CПб. 23 с.
[Karpov] Карпов В.Г. 1960. О видовом составе живых семян и запасе их в почве ельника-черничника. – Тр. МОИП. Т. 3. С. 131–140.
[Karpov] Карпов В.Г. 1969. Экспериментальная фитоценология темнохвойной тайги. Л. 335 с.
[Komarova] Комарова Т.А. 1984. Развитие Chelidonium asiaticum (Papaveraceae) при восстановлении кедрово-широколиственных лесов Южного Сихотэ-Алиня после пожара. – Бот. журн. 69 (8): 1052–1062.
[Komarova] Комарова Т.А. 1986. Семенное возобновление растений на свежих гарях (леса Южного Сихотэ-Алиня). Владивосток. 224 с.
[Komarova] Комарова Т.А. 1992. Послепожарные сукцессии в лесах Южного Сихотэ-Алиня. Владивосток. 224 с.
[Komarova, Il`ina] Комарова Т.А., Ильина Т.М. 2002. Изменение всхожести семян ели аянской в зависимости от характера послепожарного субстрата. – В кн.: Динамика и состояние лесных ресурсов Дальнего Востока. Хабаровск. С. 115–121.
[Komarova] Комарова Т.А., Прохоренко Н.Б., Глушко С.Г., Терехина Н.В. 2017. Послепожарные сукцессии в лесах Сихотэ-Алиня с участием Pinus koraiensis Siebold et Zucc. СПб. 402 с.
[Kostenkova] Костенкова А.Ф. 1978. Роль опада хвойно-широколиственных лесов Приморья в биологическом круговороте веществ. – В кн.: Вопросы генезиса, плодородия и охраны почв Дальнего Востока. Владивосток. С. 100–124.
Krahulcova A. 1982. Chelidonium majus L.s.1. Cytotaxno-mic study of Chelidonium majus L.s.1. – Folia Geobot. Et Phytotaxon. (17) 3: 237–268.
Kwiatkowska-Falińska A., Jankowska-Błaszczuk M., Jaroszewicz B. 2014. Post-Fire changes of soil seed banks in the early successional stage of Pine forest. – Polish J. Ecol. 62 (3): 455–466.
[Lyr et al.] Лир Х., Польстер Г., Фидлер Г. 1974. Физиология древесных растений. М. 421 с.
[Lyanguzova] Лянгузова И.В. 2002. Методика изучения банка семян в почве. – В кн.: Методы изучения лесных сообществ. СПб. С. 114–118.
Morimoto J., Kominami R., Koike T. 2009. Distribution and characteristics of the soil seed bank of the black locust (Robinia pseudoacacia) in a headwater basin in northern Japan. – Landscape Ecol. Eng. (6) 2: 193–199.
Ne’eman G., Izhaki I. 1999. The effect of stand age and microhabitat on soil seed banks in Mediterranean Aleppo pine forests after fire. – Plant Ecology. 144: 115–125.
[Nikolaeva] Николаева М.Г. 1967. Физиология глубокого покоя семян. Л. 206 с.
[Petrov] Петров В.В. 1989. Банк семян в почвах лесных фитоценозов Европейской части СССР. М. 197 с.
[Petrov, Belyaeva] Петров В.В., Беляева Ю.Е. 1981. Запас семян растений в почве под пологом леса. – Лесоведение. 5: 44–49.
[Petrova] Петрова А.Н. 1999. Банк семян в почвах лесных фитоценозов подзоны смешанных (елово-широколиственных) лесов. – Вестн. Нижегород. ун-та им. Н.И. Лобачевского. 1: 26–28.
Plue J., Verheyen K., Van Calster H., Marage D., Thompson K., Kalamees R., Jankowska-Blaszczuk M., Bossuyt B., Hermy M. 2010. Seed banks of temperate deciduous forests during secondary succession. – J. Veg. Sci. (21) 5: 965–978.
[Poptsov] Попцов А.В. 1968. К вопросу о роли и значении покоя семян в семенном размножении растений. – Уч. зап. Ульяновск. гос. пед. ин-та. Вопросы биологии семенного размножения. Ульяновск. 23 (3): 134–142.
[Rabotnov] Работнов Т.А. 1958. К методике изучения содержания всхожих семян в почвах лугов. – Бот. журн. 43 (11): 86–92.
[Rabotnov] Работнов Т.А. 1981. Жизнеспособные семена в составе ценотических популяций как показатель стратегии жизни видов растений. – Бюл. МОИП. Отд. Биол. 86 (3): 68–78.
[Rabotnov] Работнов Т.А. 1982. Жизнеспособные семена в почвах природных биогеоценозов СССР. – В кн: Теоретические и прикладные аспекты биогеографии. М. С. 35–39.
[Rabotnov] Работнов Т.А. 1983. Фитоценология. 2-е изд. М. 292 с.
[Rabotnov] Работнов Т.А. 1986. Жизнеспособные семена в почвах природных биогеоценозов. – Бюл. МОИП. Отд. биол. 91 (3): 3–17.
[Rysin, Rysina] Рысин Л.П., Рысина Г.П. 1965. Почвенный запас семян травянистых растений в лесу и факторы, влияющие на их прорастание. – В кн.: Леса Подмосковья. М. С. 5–27.
[Sapozhnikov] Сапожников А.П., Гавренков Г.И. 1980. О достоверности различий между почвами кедровников и ельников на Верхнеуссурийском стационаре. – В кн.: Комплексные исследования лесных биогеоценозов. Владивосток. С. 74–89.
[Sapozhnikov] Сапожников А.П., Костенкова А.Ф. 1984. Воздействие пожаров на подстилку в кедрово-широколиственных лесах Южного Сихотэ-Алиня и особенности их восстановления. – В кн.: Динамические процессы в лесах Дальнего Востока. Владивосток. С. 139–147.
Schafer D.E., Chilcote D.O. 1969. Factors influencing persistence and depletion in buried seed populations. I. A model for analysis of parameters of buried seed persistence and depletion. – Crop Sci. 9: 417–419.
[Seledets, Probatova] Селедец В.П., Пробатова Н.С. 2018. Экологическая дифференциация видов чистотела Chelidonium (Papaveraceae). – Ученые записки ЗапГУ. 13 (1): 41– 49.
[Sidorova] Сидорова О.В. 2009. Запас семян в почвах сосновых лесов и их вырубок. – В кн.: Экологические проблемы Севера: межвуз. сб. науч. тр. Архангельск. 12: 58–60.
[Soil formation…] Почвообразование и особенности круговорота веществ в горных лесах Южного Сихотэ-Алиня (на примере Верхнеуссурийского стационара). 1993. Хабаровск. 269 с.
[Sosudistye…] Сосудистые растения Советского Дальнего Востока. 1985–1996. Т. 1–8. СПб.
Taylorson R.B. 1970. Changes in dormancy and viability of weed seeds in soils. –Weed Sci. (18) 2: 265–269.
Thompson K., Grime J. 1979. Seasonal variation in the seed banks of herbaceous species in ten contrasting habitats. – J. Ecol. 67: 893–921.
Thompson K., Band S.R., Hodgson J.G. 1993. Seed size and shape predict persistence in soil. – Funct. Ecol. 7: 236–241.
[Torgashkova] Торгашкова О.Н. 2003. Основные закономерности формирования и реализации банка семян в лесах Саратовского Правобережья. – Бюл. Бот. сада Сарат. гос. ун-та. 2: 60–67.
[Torgashkova] Торгашкова О.Н. 2013. Процессы функционирования банков семян лесных сообществ Саратовского Правобережья. – Изв. Саратовского ун-та. Новая серия. Серия: Химия. Биология. Экология. 13 (4): 89–95.
Wellstein C., Otte A., Waldhardt R. 2007. Seed bank diversity in mesic grasslands in relation to vegetation type, management and site conditions. – J. Veg. Sci. 18: 153–162.
Yu S.L., Chen H.W., Lang N.J. 2007. The classification systems of soil seed banks and seed persistence in soil. – Acta Ecol. Sin. 5 (27): 2099–2108.
Дополнительные материалы отсутствуют.
Инструменты
Ботанический журнал