

Ботанический журнал, 2021, T. 106, № 9, стр. 837-854
ПАЛЕОФИТОГЕОГРАФИЯ СИБИРСКОЙ ПАЛЕОФЛОРИСТИЧЕСКОЙ ОБЛАСТИ В РАННЕЙ ЮРЕ И ПЕРВОЙ ПОЛОВИНЕ СРЕДНЕЙ ЮРЫ
А. И. Киричкова 1, *, Е. И. Костина 2, **, Н. В. Носова 1, ***
1 Ботанический институт им. В.Л. Комарова РАН
197376 Санкт-Петербург, ул. Проф. Попова, 2, Россия
2 Геологический институт РАН
119017 Москва, Пыжевский переулок, 7, Россия
* E-mail: kirichkovaanna@gmail.com
** E-mail: kostina.gin@gmail.com
*** E-mail: nnosova@binran.ru
Поступила в редакцию 10.11.2020
После доработки 19.05.2021
Принята к публикации 01.06.2021
Аннотация
Рассматриваются результаты сравнительного анализа тафофлор ранней и первой половины средней юры Сибирской палеофлористической области. Для этого временного интервала выявлено большое сходство систематического состава тафофлор Западной Сибири и Северного Китая. На основе авторских и литературных данных откорректировано положение границы на юге и юго-западе Сибирской палеофлористической области и уточнены границы между ее провинциями – Западно-Сибирской и Северо-Китайской. Уточненный систематический состав палеофлор, постоянное наличие в отложениях многочисленных остатков листьев листопадных голосеменных и седиментологические особенности флороносных толщ – все это убедительно подтверждает, что климат на территории всей Сибирской области в ранней и в первой половине средней юры был гумидным с сезонными колебаниями температуры и влажности. Кратко рассмотрены палеогеографические особенности северо-востока Сибирской палеофлористической области.
История палеофитогеографических исследований юрско-меловых флор Евразии детально изложена в фундаментальных монографиях В.А. Вахрамеева (Vakhrameev, 1964, 1988, 1991; Vakhrameev et al., 1970). На основании проведенного им сравнительного анализа систематического состава юрских и меловых тафофлор впервые были составлены палеофитогеографические карты Евразии, получившие признание широкого круга палеоботаников и исследователей континентальных толщ. На территории Евразии Вахрамеев выделил две палеофлористические области – Евро-Синийскую (первоначально Индо-Европейскую) и Сибирскую. Российские флоры раннего мела Сибири рассматривались автором совместно с раннемеловыми флорами Северной Америки (Аляски и северной Канады), поэтому палеофлористическая область этого периода была названа Сибирско-Канадской.
Евро-Синийская палеофлористическая область в ранне- и среднеюрское время занимала огромную территорию Западной и Южной Европы, юга европейской части России, Средней Азии и Южного Китая до берегов Тихого океана. Область была подразделена на провинции: Европейскую, Среднеазиатскую и Восточно-Азиатскую. Позднее Среднеазиатская провинция была разделена на подпровинции: Закаспийскую, Гиссарскую и Ферганскую (Vakhrameev, 1988, 1991).
Сибирская палеофлористическая область занимала почти весь Урал, за исключением его южной части, Восточный Казахстан, всю Сибирь, Монголию, Западный, Северный и Северо-Восточный Китай, а также северо-восточную половину европейской части России и, возможно, северную часть Скандинавии. В области не были выделены палеофлористические провинции из-за недостаточной изученности на то время сибирских ранне- и среднеюрских тафофлор (Vakhrameev, 1988, 1991).
В течение последних 30 лет были получены новые данные о флорах ранней и средней юры, фитостратиграфии как закрытых седиментационных бассейнов, так и ряда открытых разрезов и угольных бассейнов, расположенных в пределах Сибирской области. Монографически были изучены коллекции растений нижне-среднеюрских отложений стратотипических и опорных разрезов скважин и обнажений в Кузнецком угленосном бассейне (Kiritchkova et al., 1992). Это позволило не только значительно уточнить стратиграфическую схему юрских отложений этого бассейна, но и впервые разработать региональную шкалу, основанную на сукцессионно сменяющихся по разрезу фитостратиграфических комплексах.
Была проведена ревизия коллекционного фонда и изучены вновь собранные коллекции макроостатков растений из юрских отложений, вскрытых многочисленными скважинами в Западной Сибири (Mogutcheva, 2003; Kiritchkova et al., 2005), в Канском угленосном бассейне (Kostina, 2004), в разрезах стратотипической местности Иркутского бассейна (Kiritchkova et al., 2017a, b, 2018, 2020) и Казахстана (Doludenko, Orlovskaya, 1976; Doludenko et al., 1989; Sevryugin et al., 1989; Kiritchkova, Doludenko, 1996). Были получены новые данные о юрской флоре Монголии (Kostina, Herman, 2013, 2016; Kostina et al., 2015) и Северного Китая (Zhou, 1995 и др.). Эти исследования послужили основой не только для корректировки границы Сибирской области в ранне- и среднеюрское время, но и для более уверенного обоснования ранее выделенных флористических провинций в этой области.
Сибирская палеофлористическая область и ее провинции
История изучения Сибирской палеофлористической области
После фундаментальных работ Вахрамеева, касающихся фитогеографии юрских и меловых флор Евразии (Vakhrameev, 1964, 1988, 1991; Vakhrameev et al., 1970), вопросы фитогеографии прошлых эпох интересовали многих палеоботаников и геологов. К сожалению, как отмечал Вахрамеев (Vakhrameev, 1964, 1988, 1991), ранне- и среднеюрские флоры Сибирской области долгое время оставались менее изученными по сравнению с флорами Евро-Синийской области. Сибирская область не была разделена на палеофлористические провинции, несмотря на отмечаемые автором некоторые различия систематического состава тафофлор ее северных и южных территорий.
Исследования по палеофитогеографии Сибирской области, начавшиеся в конце прошлого столетия и продолжающиеся до настоящего времени, проводятся в основном российскими и китайскими палеоботаниками. При этом китайские специалисты свои представления о палеофитогеографии по большей части ограничивают территорией Китая, отмечая при этом сходство систематического состава сибирских и северокитайских тафофлор.
В 1949 году китайский палеоботаник Сы (Sze, 1949) впервые отметил, что по систематическому составу флоры ранней и средней юры Северного Китая очень сходны с одновозрастными флорами Сибири, известными к тому времени лишь по работам О. Геера (Heer, 1876, 1878, 1880) и В.Д. Принады (Prynada, 1962). Флоры Китая этого возраста Сы разделил на Северные и Южные. Северные тафофлоры, флоры сибирского типа, с доминирующими в их составе папоротниками (Cladophlebis, Raphaelia, Coniopteris), гинкговыми (Ginkgo/Ginkgoides, Sphenobaiera, Baiera), лептостробовыми (Czekanowskia, Phoenicopsis) и хвойными (Podozamites и Elatocladus), Сы объединил во флору Coniopteris–Phoenicopsis. Южные тафофлоры, в составе которых присутствуют папоротники семейств Dipteridacеaе, Marattiacеaе, Matoniaceae, а голосеменные представлены разнообразными беннеттитовыми (Nilssoniopteris, Pterophyllum, Otozamites) и хвойными преимущественно с чешуевидными листьями, были объединены во флору Dictyophyllum–Clathropteris (Sze, 1949, 1956; Sze, Chow, 1962).
В 1983 году Чжоу Чжиянь (Zhou, 1983) на территории Евразии и востока Гренландии для ранне- и среднеюрского времени выделил две палеофлористические провинции: Континентальную (Continental Floristic Province) и Морскую (Maritime Floristic Province). К Континентальной провинции в пределах Северного Китая он отнес тафофлоры сибирского типа, т.е. флоры Coniopteris–Phoenicopsis, известные к тому времени на северо-востоке (Zhang, Zheng, 1987) и на территории Внутренней Монголии Китая. К этой же провинции Чжоу Чжиянь отнес ранне-среднеюрские флоры Южного Урала, южного Казахстана, Ферганы и юго-запада Киргизии, ранее относимые Вахрамеевым к Среднеазиатской провинции Евро-Синийской области (Vakhrameev, 1964; Vakhrameev et al., 1970). В связи с этим Чжоу Чжиянь считает, что поскольку среднеазиатские тафофлоры больше соответствуют сибирской флоре Coniopteris–Phoenicopsis, нежели тетическим флорам Южного Китая (т.е. флорам близ южного океана Тетис), то среднеазиатские флоры должны быть отнесены к Континентальной провинции, исключая Евро-Синийскую область (Zhou, 1995: 390–391). Ранне-среднеюрские флоры юго-восточного и южного Китая, флоры Dictyophyllum–Clathropteris, он отнес к Морской провинции (Zhou, 1983, 1995: 345, text-fig. 8-1).
В 1992 г. Сунь Чунь-Лин (Sun Ch. L., 1992) выделил для ранней – начала средней юры Китая три палеофлористические провинции. Флоры сибирского типа, т.е. флоры Coniopteris–Phoenicopsis, распространенные, главным образом, в пределах Северного Китая, составили две провинции: провинцию Дальнего Севера (Far North Floristic Province) и Северную провинцию (North Floristic Province). Во флористическую провинцию Дальнего Севера вошли тафофлоры северо-запада (тафофлоры Джунгарского бассейна) и часть тафофлор северо-востока Северного Китая. Южная граница провинции была проведена вдоль горных хребтов Тянь-Шаня и Иньшаня. В Северную флористическую провинцию вошли тафофлоры Внутренней Монголии и северо-востока Китая. Южная граница этой провинции отмечена вдоль южной границы Внутренней Монголии и горных хребтов Куньлунь, Циньлин и Даби. Южнее этой границы тафофлоры, соответствующие флоре Dictyophyllum–Clathropteris, выделены в Южно-Китайскую провинцию (South-China Province). При этом Сунь Чунь-Лин отмечал, что комплексы растений в тафофлорах двух северных провинций, хотя и соответствуют сибирскому типу, т.е. флоре Coniopteris–Phoeni-copsis, несколько различаются между собой. В тафофлорах Северной провинции, в отличие от тафофлор провинции Дальнего Севера, присутствуют немногочисленные, но постоянные Dipteridaceae и Cycadopsida. Предложенные Сунь Чунь-Лином палеофитогеографические построения на территории Китая в дальнейшем были использованы другими исследователями юрских флор Китая (Zhou, 1995; Wang et al., 2005; Huang W. et al., 2016; Deng et al., 2017; Na et al., 2017).
Тафофлоры ранней и начала средней юры Казахстана Вахрамеев относил к двум палеофлористическим областям (Vakhrameev, 1964, 1988, 1991; Vakhrameev et al., 1970). К Сибирской области были отнесены тафофлоры северо-востока Казахстана (Майкубень, Алаколь, Кендерлык), а к Евро-Синийской области (Среднеазиатская провинция) – тафофлоры южного Казахстана (хребет Каратау, Ленгерский бассейн, Илийский угольный бассейн).
На III-м Межведомственном стратиграфическом совещании (МСК) по Казахстану, проходившем в 1986 году в г. Алма-Ате (сейчас Алматы), в стратиграфии опорных разрезов ряда местонахождений Казахстана были выявлены значительные недостатки в построении и палеофлористическом обосновании разработанных к тому времени стратиграфических схем континентальных отложений региона. По рекомендации МСК группой местных геологов совместно с палеоботаниками были целенаправленно проведены стратиграфо-палеофлористические исследования принятых на совещании опорных разрезов нижне-среднеюрских отложений, в первую очередь хребта Каратау на юге (Таскомырсайское и Чокпакское месторождения угля), на юго-востоке (Ленгерская и Кельтемашатская впадины) и северо-востоке (Майкубенское месторождение угля) Казахстана. В результате этих исследований были значительно уточнены систематический состав комплексов растений, их возрастной уровень, стратиграфия соответствующих отложений, а также обосновано латеральное соотношение разнофациальных разрозненных стратонов (Sev-ryugin et al., 1989). Более того, было выявлено, что комплексы растений первой половины юры не только северо-восточного, но и южного Казахстана по систематическому составу доминирующих таксонов соответствуют флорам сибирского типа (Doludenko et al., 1989; Sevryugin et al., 1989, 1990). Полученные результаты послужили основанием для уточнения юго-западной границы Сибирской палеофлористической области (Kiritchkova, Doludenko, 1996).
Представления о положении южной границы Сибирской палеофлористической области и выделение в ee пределах палеофлористических провинций для ранне- и среднеюрского времени были опубликованы в монографии, посвященной юрской флоре Западной Сибири (Kiritchkova et al., 2005). Авторами были выделены фитостратиграфические комплексы, сукцессионно сменяющиеся по разрезу и датированные морской фауной (Shurygin et al., 2000). Результаты сравнительного анализа систематического состава западно-сибирских комплексов с тафофлорами юго-востока Средней Азии и Северного Китая показали, что ранне-среднеюрская флора сибирского типа занимала бóльшую территорию, чем предполагал Вахрамеев (Vakhrameev, 1964, 1988, 1991). В связи с этим было предложено проводить южную границу Сибирской области на уровне примерно 35°с.ш. вместо 40° с.ш., как предлагал Вахрамеев (1964). Кроме того, отмечалось, что систематический состав тафофлор в северных и южных частях Сибирской области несколько различен, что позволило впервые на территории области выделить две палеофлористические провинции: Западно-Сибирскую и Северо-Китайскую. Граница между провинциями была условно проведена с востока вдоль северной границы Китая до Аральского моря на западе (Kiritchkova et al., 2005).
К настоящему времени представления о юрской флористике Сибирской области значительно расширились (Kiritchkova et al., 1992, 2005; Kostina, 2004; Frolov, Mashchuk, 2018). На современном уровне изучена юрская флора Иркутского угленосного бассейна, широко известная со времен Геера (Heer, 1876, 1878, 1980), а затем Принады (Prynada, 1962). Кроме классических коллекций Геера материалом для исследований послужили послойно собранные образцы с макроостатками растений и миоспорами из естественных, но разрозненных разрезов стратотипической местности Иркутского бассейна. В результате был уточнен систематический состав иркутской палеофлоры из опорных разрезов Иркутского бассейна, впервые изученных еще в прошлом столетии. Особое внимание было уделено доминирующей группе голосеменных растений, получивших эпидермальную характеристику (Kiritchkova, Travina, 1993; Kiritchkova et al., 2016, 2017a, b, 2018, 2020; Nosova, Kiritchkova, 2018; Nosova et al., 2017a, b; 2018). Были выявлены флорогенетические изменения иркутской палеофлоры во времени, что послужило надежной основой для разработки первой региональной комплексной биостратиграфической шкалы. Это, в свою очередь, дало возможность обосновать латеральную корреляцию разнофациальных стратонов. Более того, впервые была дана послойная палинологическая характеристика отложений из этих же стратотипических разрезов (Kiritchkova et al., 2020).
За последние годы были детально изучены среднеюрские флоры Монголии, по своему систематическому составу соответствующие флорам сибирского типа (Kostina, Herman, 2013, 2016, Kostina et al., 2015). Комплексы растений, установленные в юрских отложениях Западной, Центральной и самого юга Южной Монголии, уверенно сопоставляются со среднеюрскими ангарским и прииркутско-ийским комплексами растений присаянской свиты Иркутского бассейна. Это послужило основанием отнести монгольские тафофлоры к Западно-Сибирской провинции, обозначив при этом ее южную границу вдоль южной границы Монголии, т.е. южнее, чем предполагалось ранее (Kiritchkova et al., 2005).
Были детально исследованы континентальные отложения верхнего триаса, нижней и средней юры некоторых опорных разрезов Северного Китая, таких как Джунгарский угольный бассейн (Sun G. et al., 2010; Li Sh. L. et al., 2014), северо-западная часть угленосного бассейна Цайдам в провинции Цинхай (Qinghai) (Li P. et al., 1988; Zhou, 1995; Wang et al., 2005), бассейн Ордос во Внутренней Монголии (Li Sh. L. et al., 2014; Sun Ch. L. et al., 2015b; Huang et al., 2016) и др.
Немалую роль для палеофитогеографических построений сыграли работы по детальному изучению таксонов, особенно доминирующих во флористическом комплексе той или иной палеофлоры. Для юрских флор Сибирской палеофлористической области это, в первую очередь, представители гинкговых – Ginkgo/Ginkgoites (Doludenko, Rasskazova, 1972; Sun Ch. L. et al., 2008; Kostina et al., 2015), Sphenobaiera (Sun G. et al., 2006; Wang et al., 2005; Kostina, Herman, 2013; Kiritchkova et al., 2016), Pseudotorellia (Krassilov, 1972; Kiritchkova, Nosova 2009; Nosova, Kiritchkova, 2010; Kostina et al., 2015), Leptotoma (Kiritchkova, Samylina, 1979; Travina, 1996; Kiritchkova et al., 2005; Nosova, 2010), лептостробoвые (чекановскиевые) – Phoenicopsis (Samylina, 1972; Samylina, Srebrodolskaya, 1986; Zhou, Zhang, 1998; Kiritchkova et al., 2002b, 2005; Li T. et al., 2014; Sun Ch. L. et al., 2015a; Huang et al., 2016) и Czekanowskia (Samylina, Kiritchkova, 1991; Sun Ch. L. et al., 2009, 2015b); из хвойных – Podozamites (Nosova et al., 2017b), Elatocladus (Kiritchkova et al., 2005; Nosova et al., 2017a), из папоротников – Coniopteris и Cladophlebis (Kiritchkova, Travina, 1993; Kiritchkova et al., 2002а, 2005; Deng et al., 2017).
Подобные исследования сделали таксоны более конкретными. Морфологические характеристики у многих представителей голосеменных были дополнены признаками эпидермально-кутикулярного строения их листьев, что позволило определять до вида растительные остатки со сходной морфологией. Это, в свою очередь, сделало возможным уточнить систематический состав палеофлор, возрастной диапазон характерных таксонов и их распространение по площади.
Границы Сибирской палеофлористической области
Особенности флористического комплекса11 юрской флоры сибирского типа были сформулированы Вахрамеевым при выделении Сибирской флористической области (Vakhrameev, 1964; Vakhrameev et al., 1970). Позднее монографически изученные остатки растений из юрских отложений многочисленных скважин Западной Сибири, особенно северной ее половины, и опорных разрезов Кузнецкого и Канского угленосных бассейнов сделали возможным не только значительно расширить и конкретизировать систематический состав сибирских тафофлор, но и уточнить их возрастной уровень и систематический состав флористического комплекса юрской флоры Сибирской палеофлористической области в целом (Kiritchkova et al., 1992, 2005; Kostina, 2004).
Основными элементами палеофлоры Сибирской области являются многочисленные хвощовые – Neocalamites (особенно в ранней юре), Equisetites и Phyllotheca; папоротники – Coniopteris (немногочисленные в ранней, но превалирующие в средней юре), многочисленные и разнообразные в видовом отношении Cladophlebis и Raphaelia, более редкие Hausmannia, Clathropteris, Phlebopteris; среди голосеменных преобладают разнообразные в родовом и видовом отношениях гинкгофиты – Ginkgo/Ginkgoites, Sphenobaiera, Baiera, Pseudotorellia, Leptotoma, Phoenicopsis, Czekanowskia; присутствуют неразнообразные и немногочисленные цикадофиты – Nilssonia, Anomozamites, Pterophyllum, Ctenis, Taeniopteris; среди хвойных преобладают Podozamites, Elatocladus, Pityophyllum, реже Pagiophyllum.
Обширная территория Сибирской палеофлористической области неизбежно предполагает региональную гетеротаксонность северных и южных тафофлор. На большей части Северного Китая систематический состав и соотношение доминирующих таксонов в ранне-среднеюрских тафофлорах сходны с сибирскими и более всего с комплексами растений Иркутского и Кузнецкого бассейнов. Китайскими палеоботаниками именно эти тафофлоры Северного Китая были выделены в Континентальную палеофлористическую провинцию (Zhou, 1983, 1995), позже – в провинции Дальнего Севера и Северную. К последней из них были отнесены также тафофлоры северо-востока Средней Азии (Sun Ch.L., 1992). При этом авторы отмечали, что в составе тафофлор южной части Северного Китая и северо-востока Средней Азии, наряду с перечисленными выше доминантами флор сибирского типа, чаще отмечается присутствие диптериевых и матониевых папоротников и цикадофитов (Sun Ch.L., 1992; Zhou, 1983, 1995; Huang et al., 2016; Na et al., 2017; Li et al., 2019).
На основе этих данных, а также результатов проведенного нами анализа тафофлор северо-востока Средней Азии, мы предлагаем южную границу Сибирской палеофлористической области для ранней и первой половины средней юры проводить в пределах Северного Китая вдоль горных хребтов Куньлунь – Циньлин – Даби, примерно на уровне 33–35° с.ш., что на 5–7° южнее, чем предполагал Вахрамеев (Vakhrameev et al., 1970), а юго-западную границу проводить, включая в пределы Сибирской области северо-восточную часть Средней Азии (рис. 1).
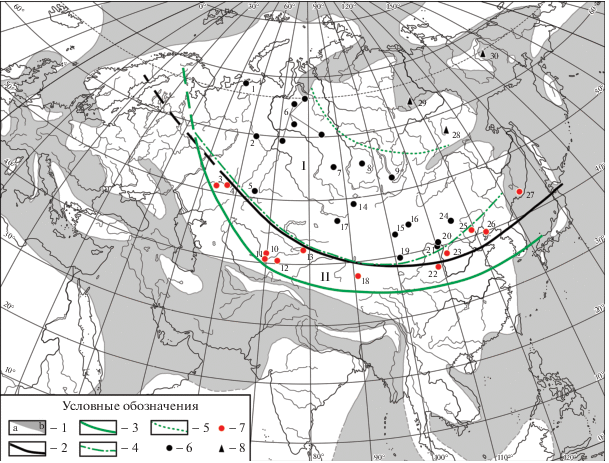
Рис. 1.
Схематическая карта Сибирской палеофлористической области и ее провинций: границы, местонахождения основных опорных разрезов ранне-среднеюрских отложений.
I – Западно-Сибирская провинция, II – Северо-Китайская провинция.
1: a – суша, b – море; 2 – граница между Сибирской и Евро-Синийской палеофлористическими областями по В.А. Вахрамееву (Vakhrameev et al., 1970); 3 – граница Сибирской флористической области (уточненная в данной статье); 4 – граница между Западно-Сибирской и Северо-Китайской провинциями (уточненная в данной статье); 5 – условная граница Западно-Сибирской провинции на северо-востоке Сибирской области; 6–8 – основные опорные разрезы ранне-среднеюрских отложений: 6 – в Западно-Сибирской провинции, 7 – в Северо-Китайской провинции, 8 – на северо-востоке Сибирской области.
Цифры на карте: 1–30 – местонахождения тафофлор: 1 – бассейн р. Печоры; 2 – Восточный Урал; 3 – бассейн р. Илек; 4 – Орский угольный бассейн; 5 – Тургай; 6 – Западня Сибирь; 7 – Кузнецкий угольный бассейн; 8 – Канский угольный бассейн; 9 – Иркутский угольный бассейн; 10 – южный Казахстан; 11 – Ангренский угольный карьер; 12 – Восточно-Ферганский угольный бассейн; 13 – Иссык-Кульская впадина; 14–16 – Монголия: 14 – Ошин-Боро-Удзюр-Ула, 15 – угольное месторождение Нарийн-Сухайт, 16 – угольный карьер Цаган-Ово; 17–26 – Северный Китай: 17 – Джунгарский бассейн, 18 – бассейн Цайдам, 19 – Алашань-Юци, 20 – бассейн Шигуай, 21 – бассейн Ордос, 22 – Шэньси, 23 – бассейн Шаньси, 24 – угольный бассейн Шилин-Хото, 25 – Даохугу, 26 – провинция Ляонин; 27 – бассейн р. Петровки (Южное Приморье), 28 – Южно-Якутский угольный бассейн, 29 – р. Вилюй, нижнее течение, 30 – бассейн р. Колымы.
Fig. 1. Schematic map of the Siberian palaeofloristic region and provinces: boundaries, localities of the main key stratigraphic sections of the Early-Middle Jurassic deposits.
I – West Siberian province, II – North Chinese province.
1: a – land, b – sea; 2 – boundary between the Siberian and Euro-Sinian palaeofloristic regions (after Vakhrameev et al., 1970); 3 – boundary of the Siberian palaeofloristic region (emended in the present paper); 4 – boundary between the West Siberian and the North Chinese provinces (emended in the present paper); 5 – conventional boundary of the West Siberian province in the north-east of the Siberian region; 6–8 – main key stratigraphic sections: 6 – in the West Siberian province, 7 – in the North Chinese province, 8 – in the north-east of the Siberian region.
Figures on the map: 1–30 – localities of taphofloras: 1 – Pechora River Basin; 2 – Eastern Urals; 3 – Ilek River Basin; 4 – Orsk Coal Basin; 5 – Turgay; 6 – West Siberia; 7 – Kuznetsk Coal Basin; 8 – Kansk Coal Basin; 9 – Irkutsk Coal Basin; 10 – Southern Kazakhstan; 11 – Angren coal mine; 12 – East Fergana Coal Basin; 13 – Issyk-Kul Depression; 14–16 – Mongolia: 14 – Oshin-Boro-Udzyur-Uul, 15 – Nariin-Shukhait Coalfield, 16 – Tsagaan-Ovoo coal quarry; 17–26 – North China: 17 – Junggar Basin, 18 – Qaidam Basin, 19 – Area Alxa Right Banner, 20 – Shiguai Basin, 21 – Ordos Basin, 22 – Shaanxi, 23 – Shanxi Basin, 24 – Xilinhot Coal Basin, 25 – Daohugou Area, 26 – Liaoning Province; 27 – Petrovka River Basin (South Primorye); 28 – South Yakutsk Coal Basin, 29 – lower reaches of the Vilyuy River, 30 – Kolyma River Basin.
Западно-Сибирская палеофлористическая провинция
Новые данные о фитостратиграфии юрских континентальных отложений и тафофлорах Северного Китая значительно расширили наши представления о ранее выделенных на территории Сибирской области палеофлористических провинциях. Выявлено, что тафофлоры Западно-Сибирской провинции на территории Северного Китая имели бóльшее распространение. Южная граница провинции проходила заметно южнее, чем мы предполагали ранее (Kiritchkova et al., 2005; Kostina, Herman, 2016). Одним из опорных разрезов Западно-Сибирской провинции в пределах Северного Китая является разрез юрских отложений Джунгарского угольного бассейна (рис. 1). Первый исследователь юрских растений этого бассейна А. Сьюорд (Seward, 1911) считал, что они подобны сибирским. Вахрамеевым эта флора была отнесена к флорам Сибирской палеофлористической области (Vakhrameev et al., 1970). Позже китайские палеоботаники также определяли палеофлору Джунгарского бассейна как сибирскую и относили ее к Континентальной палеофлористической провинции (Zhou, 1983) или к провинции Дальнего Севера (Sun G. et al., 2010; Li Sh.-L. et al., 2014).
Отложения нижней юры в Джунгарском бассейне (формации Badaowan и Sangonghe) залегают на континентальных отложениях верхнего триаса и охарактеризованы флорой Neocalamites–Marattiopsis (рис. 2). Комплексы растений этих формаций немногочисленны и в систематическом плане неразнообразны. Это хвощовые (Neocalamites, Equisetites), папоротники (Cladophlebis, Clathropteris, Phlebopteris, единичные Coniopteris), а также единичные гинкговые (Ginkgoites, Sphenobaiera) и хвойные (Podozamites, Pityophyllum). Джунгарская палеофлора Neocalamites–Marattiopsis по соотношению характерных таксонов сопоставима с раннеюрскими комплексами растений Западной Сибири (с ягельным комплексом) и Иркутского бассейна (c белореченским, но в основном с черемхово-азейским комплексом) (рис. 2). Наличие в джунгарской тафофлоре теплолюбивых папоротников Clathropteris и Phlebopteris было обусловлено не только более южным ее расположением, по сравнению с другими тафофлорами сибирского типа, но и, как предполагали китайские палеоботаники, влиянием флор Морской палеофлористической провинции, существовавшей в это время вдоль берегов океана Тетис (Zhou, 1983, 1995).
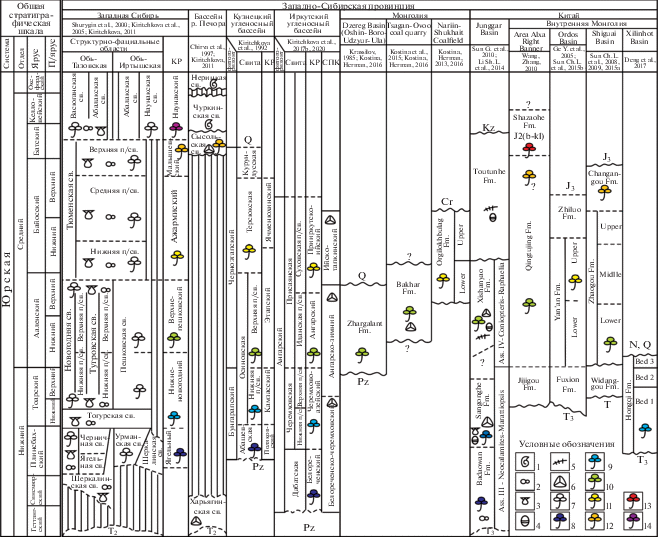
Рис. 2.
Корреляция континентальных отложений ранней и первой половины средней юры Западно-Сибирской провинции Сибирской палеофлористической области по палеоботаническим данным.
Сокращения: п/я – подъярус; св. – свита; п/св. – подсвита; Fm – Formation; Ass. – Assemblage; КР – комплекс растений; СПК – спорово-пыльцевой комплекс.
Условные обозначения: 1–7 – ископаемые остатки: 1 – аммоноидеи, 2 – фораминиферы, 3 – двустворки, 4 – конхостраки, 5 – наземные позвоночные, 6 – споры и пыльца, 7 – макроостатки растений; 8–14 – возрастной уровень тафофлор: 8 – нижняя половина ранней юры, 9 – верхняя половина ранней юры, 10 – аален, 11 – байос, 12 – байос-бат, 13 – бат-келловей, 14 – келловей-оксфорд.
Fig. 2. Correlation of the Lower and first half of the Middle Jurassic continental deposits of the West Siberian province of the Siberian palaeofloristic region based on the palaeobotany data.
Abbreviations: п/я – substage; св. – suite; п/св. – subsuite; Fm – Formation; Ass. – Assemblage; КР – plant complex; СПК – spores and pollen complex.
Legend: 1–7 – fossils: 1 – ammonites, 2 – foraminifera, 3 – bivalves, 4 – conchostraca, 5 – terrestrial vertebrates, 6 – spores and pollen, 7 – plant macroremains; 8–14 – floras ages: 8 – lower half of the Early Jurassic, 9 – upper half of the Early Jurassic, 10 – Aalenian, 11 – Bajocian, 12 – Bajocian-Bathonian, 13 – Bathonian-Callovian, 14 – Callovian-Oxfordian.
Тафофлора из отложений начала средней юры джунгарского разреза (формация Xishanyao) относится к флоре Coniopteris–Phoenicopsis и характеризуется разнообразием Coniopteris и многочисленными лептостробовыми (чекановскиевыми). Это делает ее сопоставимой с ангарским комплексом Иркутского бассейна и верхнепешковским комплексом Западной Сибири (рис. 2).
Сравнительный анализ систематического состава других ранне-среднеюрских тафофлор сибирского типа, известных из отложений на территории Внутренней Монголии Китая, выявил их сходство более всего с тафофлорами Иркутского угленосного бассейна (Kiritchkova et al., 2018, 2020). Тафофлоры второй половины ранней юры формации Hongqi угольного бассейна Шилин-Хото на юго-востоке Внутренней Монголии (Deng et al., 2017) по систематическому составу мало отличимы от раннеюрского черемхово-азейского комплекса Иркутского бассейна. Тафофлоры начала средней юры формации Zhaogou бассейна Шигуа (Sun Ch. L. et al., 2008, 2009, 2015a) и формации Qingtujing района Алашань-Юци (Wang, Zhang, 2010) оказались сопоставимы с ангарским комплексом Иркутского бассейна (рис. 1, 2). Систематический состав комплекса растений формации Yan’an бассейна Ордос (Ge et al., 2005; Sun Ch. L. еt al., 2015b) соответствует прииркутско-ийскому комплексу Иркутского бассейна и комплексу растений формации Orgilokhbulag угольного бассейна Нарийн Сухайт Монголии (Kostina, Herman, 2013, 2016) (рис. 1, 2).
Подобная сопоставимость китайских тафофлор, относимых к двум Северным провинциям (Sun Ch. L., 1992), с тафофлорами Западной Сибири и особенно Иркутского бассейна предполагает, что в раннеюрское и аален-байосское время средней юры тафофлоры Западно-Сибирской палеофлористической провинции существовали не только в пределах провинции Дальнего Севера, но и на большей части Северной провинции Китая. Это послужило основанием южную границу Западно-Сибирской провинции провести южнее границы Внутренней Монголии, условно в пределах 38–42°с.ш. (рис. 1).
Северо-Китайская палеофлористическая провинция
Тафофлоры первой половины юры (ранняя юра – аален-байос средней юры) южной части Северного Китая китайскими палеоботаниками определялись как флоры сибирского типа и входили в Северную палеофлористическую провинцию (Sun Ch. L., 1992; Zhou, 1995). Опорным разрезом нижне-среднеюрских отложений этого региона признан разрез бассейна Цайдам (рис. 1). Руководящими в систематическом составе тафофлор первой половины ранней юры (формации Xiaomeigou и Huoshaoshan) являются хвощовые рода Neocalamites и папоротники, составляющие местами до 50% от общего числа таксонов и представленные, главным образом, родом Cladophlebis (15 видов). Кроме них присутствуют немногочисленные цикадофиты – Ctenis, Pseudoctenis, Anomozamites, Zamites, Cycadolepis; единичные гинкговые; из хвойных преобладают Podozamites и Pityophyllum. Тафофлоры второй половины ранней юры этого же бассейна (формация Tianshuigou) менее разнообразны в видовом отношении. Доминирующими в их составе остаются папоротники Cladophlebis с редкими, но постоянно присутствующими Coniopteris совместно с Clathropteris и Phlebopteris. По систематическому составу характерных таксонов тафофлоры бассейна Цайдам сопоставимы с комплексами растений формации Yongdinzhuang бассейна Шанси и не менее близки с тафофлорами северо-востока Средней Азии (рис. 1, 3).
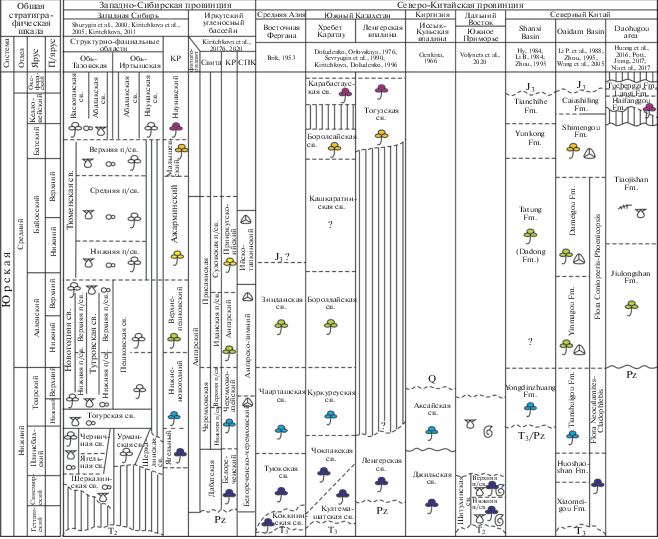
Рис. 3.
Корреляция континентальных отложений ранней и первой половины средней юры Западно-Сибирской и Северо-Китайской провинций Сибирской палеофлористической области по палеоботаническим данным.
Сокращения и условные обозначения см. рис. 2.
Fig. 3. Correlation of the Lower and first half of the Middle Jurassic continental deposits of the West Siberian and North Chinese provinces of the Siberian palaeofloristic region based on the palaeobotany data.
For explanations and legend see Fig. 2.
Тафофлоры начала средней юры южной части Северного Китая более разнообразны по сравнению с раннеюрскими и проявляют заметно большее сходство между собой. В составе тафофлор опорного разреза бассейна Цайдам (формации Yinmagou и Dameigou) превалируют папоротники из родов Conioptеris, Gonatosorus, Raphaelia, с менее частыми Cladophlebis. Кроме папоротников присутствуют хвощовые Equisetites, более разнообразные гинкговые – Ginkgo, Baierа, Sphenobaiera, Eretmophyllum, Pseudotorellia. Лептостробовые и хвойные немногочисленны и представлены тем же составом, что и в раннеюрских комплексах (Li P. et al., 1988; Zhou, 1995). По систематическому составу этим тафофлорам соответствуют тафофлоры формации Jiulongshan местонахождения Даохугоу на северо-востоке Китая (Huang et al., 2016; Na et al., 2017; Pott, Jiang, 2017) (рис. 1, 3). Тафофлоры этого местонахождения представлены значительным количеством папоротников Coniopteris (до 10 видов), Cladophlebis (4–6 видов), Dictyophyllum, Marattiopsis. Более разнообразными здесь являются беннеттитовые (Pterophyllum, Anomozamites, Ptilophyllum и др.), гинкговые и лептостробовые.
О том, что ранне-среднеюрские тафофлоры северо-востока Средней Азии по систематическому составу соответствуют флорам Сибирской палеофлористической области, неоднократно высказывались многие палеоботаники (Li B., Hu B., 1984; Sun Ch.L., 1992; Kiritchkova, Doludenko, 1996; Kiritchkova et al., 2005). Отмеченные выше новые данные о тафофлорах южной части Северного Китая выявили несомненное сходство их с тафофлорами ранней и начала средней юры северо-востока Средней Азии, ранее относимыми к Среднеазиатской провинции Евро-Синийской области (Vakhrameev et al., 1970). Очевидно сходство систематического состава тафофлор бассейнов Цайдам, Шаньси, местонахождения Даохугу провинции Хэбэй (рис. 2) с тафофлорами Южного Казахстана (Ленгерская и Кельтемашатская впадины) (Kiritchkova, Doludenko, 1996) и Киргизии (Иссык-Кульская впадина) (Genkina, 1966).
Не менее сходны с китайскими тафофлорами флоры ранней и первой половины средней юры Ферганы (рис. 3). В составе ферганских тафофлор первой половины ранней юры (коккиинская и туюкская свиты) (Brik, 1935, 1937, 1952, 1953; Markovich, 1971; Genkina, 1977) также многочисленны хвощовые – Neocalamites, многочисленны и разнообразны папоротники – крупнолистные Cladophlebis (около 10 видов), единичные Coniopteris, Osmundopsis, Phlebopteris, Gonatosorus, Clathropteris. Цикадофиты представлены довольно редкими Nilssonia, Taeniopteris, Pterophyllum, Ctenis. Гинкгофиты не отличаются особым разнообразием – это Ginkgo, Sphenobaiera, Phoenicopsis. В тафофлорах верхней половины нижней юры Восточной Ферганы (чаарташская свита) соотношение таксонов остается прежним, но среди папоротников ведущее место занимает род Coniopteris (9 видов), Cladophlebis менее разнообразен (4 вида) и представлен мелколистными формами, присутствуют Marattiopsis, Klukia и Eboracia. Более часты цикадофиты, кроме отмеченных выше родов присутствуют Ptilophyllum, Otozamites, Cycadites. Из гинкгофитов в отложениях многочисленны листья Phoenicopsis и Ginkgo с сильно рассеченной пластинкой. Хвойные неразнообразны – это Podozamites, Pityophyllum, Pagiophyllum. Ферганский комплекс растений начала средней юры (зинданская свита) отличается меньшим разнообразием систематического состава, хотя в нем по-прежнему преобладает род Сoniopteris, а Cladophlebis представлен небольшим количеством видов, присутствуют редкие цикадофиты (Nilssonia, Taenio-pteris, Anomozamites) и тот же набор хвойных. Как предполагала М.И. Брик, подобное обеднение систематического состава восточно-ферганских тафофлор в начале средней юры, начавшееся, по всей вероятности, еще в конце ранней юры, связано с изменением экологических условий, обусловленных большей дифференциацией рельефа (Brik, 1937, 1952). Но, несмотря на отмеченные особенности среднеюрских тафофлор Восточной Ферганы, их систематический состав и состав доминирующих таксонов сопоставимы с палеофлорами сибирского типа южной части Северного Китая (рис. 3), т.е. с тафофлорами Северо-Китайской палеофлористической провинции.
К Северо-Китайской провинции следует отнести и раннеюрскую флору Южного Приморья (Дальний Восток России), недавно изученную Е.Б. Волынец с соавторами (Volynets at al., 2020) (рис. 1). Комплексы растений происходят из континентальных прослоев в морских отложениях шитухинской свиты бассейна реки Петровка. Нижнеюрский (геттанг–синемюр) возраст отложений обоснован находками аммонитов. Как отмечают авторы, в составе раннеюрских тафофлор Южного Приморья преобладают папоротники, главным образом род Cladophlebis, наряду с нередкими представителями теплолюбивых Matoniaceaе, Marattiaceae, Dipteridacae. Голосеменные представлены цикадофитами, в основном Ctenis и Pterophyllum, более редкими лептостробовыми и гинкговыми, среди хвойных преобладают Podozamites, Elatocladus, Pityophyllum. В целом тафофлоры шитухинской свиты Южного Приморья – это флоры сибирского типа и мало отличаются от тафофлор южных территорий Северного Китая. По систематическому составу южно-приморские тафофлоры сопоставимы с раннеюрскими тафофлорами формации Yihe провинции Цзилинь на северо-востоке Китая (Sun G. et al., 2010) и с тафофлорами опорного разреза бассейна Цайдам формаций Xiaomeigou и Huoshaoshan (Li P. et al., 1988; Zhou, 1995). Не менее сходна раннеюрская шитухинская флора с чокпак-кельтемашатским комплексом растений хребта Каратау Южного Казахстана (Kiritchkova, Doludenko, 1996). Волынец с соавторами, проведя широкомасштабный сравнительный анализ систематического состава южно-приморских тафофлор с тафофлорами ранней юры не только Китая, но и Северной Европы, отнесли исследуемые шитухинские тафофлоры к флорам экотона, т.е. переходным от флор сибирского к флорам евросинийского типа. Это вполне вероятно, поскольку еще Вахрамеев, определяя характер ранне-среднеюрских флор южной части Северного Китая, отмечал, что “…их можно с небольшой натяжкой рассматривать как переходные от флор Сибирской к флорам Евро-Синийской области, т.е. как флоры экотона” (Vakhrameev, 1988: 40).
Палеогеографические особенности северо-восточной части Сибирской палеофлористической области
Северо-восточная часть Сибирской области в ранне-среднеюрское время по большей части была занята морем (Paleogeografiya…, 1975). Максимум морской трансгрессии зафиксирован в конце ранней юры, когда море (“Ленское море”) занимало территорию не только современного Приверхоянья, но и бассейна р. Вилюй (“Вилюйский залив”). Сушей оставались лишь Среднесибирская и Таймырская возвышенности и Байкальское нагорье. На занятой морем территории в конце ранней и в начале средней юры формировались морские отложения с аммонитовой фауной. Однако, в одном из береговых обнажений среднего течения р. Вилюй в нижнеюрских отложениях морского генезиса (сунтарская свита, тоар) были найдены остатки листьев, принадлежащие Ptilophyllum sibiricum Samyl. – одному из характерных таксонов флор Евро-Синийской области (Kiritchkova, 1984). Остатки листьев евросинийских таксонов (Thaumatopteris schenkii Nath., Ptilophyllum sibiricum Samyl., Dicroidium sp.) были найдены также в морских отложениях средней юры (аален) в береговых обнажениях р. Колымы (Samylina, Efimova, 1968). Казалось, что будет трудно объяснить присутствие характерных для евросинийской тропической флоры растений в составе флор Сибирской палеофлористической области, на территории которой преобладал гумидный климат (Vakhrameev, 1991; Zhou, 1995). А.Л. Тахтаджян при анализе условий расселения современных растений, в частности некоторых таксонов прибрежных ценозов, предположил возможность расселения их по миграционному “коридору”, т. е. вдоль побережья или по островным дугам (Takhtajan, 1970). Вполне вероятно, что вдоль тихоокеанского побережья Восточной Сибири и по островным дугам в течение позднего триаса и первой половины юры могли проникнуть далеко на север некоторые наиболее мобильные таксоны из южных территорий, в частности из Южно-Китайской провинции Евро-Синийской области.
Комплекс растений сибирского типа начала ранней юры на территории северо-восточной части Сибирской палеофлористической области известен из укугутской свиты бассейна р. Вилюй. В опорном разрезе правого берега верхнего течения р. Вилюй свита представлена пресноводными осадками. Она несогласно залегает на иреляхской свите (условно верхний триас – начало нижней юры (геттанг)), и также несогласно перекрывается морскими отложениями тюнгской свиты (нижний плинсбах) (Kirina, 1966; Kiritchkova, 1966, 1985). Типично сибирский укугутский комплекс растений насчитывает небольшое количество таксонов: Annulariopsis sp., Neocalamites sp., Equisetites lindensis Kiritchk., E. viluensis Kiritchk., Hausmannia ussuriensis Krysht., Cladophlebis vaccensis Ward., C. cf. whitbiensis (Brongn.) Brongn., Cladophlebis sp., Raphaelia diamensis Sew., Ginkgo celebris Kiritchk., Sphenobaiera ex gr. spectabilis (Nath.) Flor., Czekanowskia viluensis Kiritchk. et Samyl., Phoeni-copsis sp., Schizolepidopsis glumaceus (Kiritchk.) Doweld, S. liassica (Kiritchk.) Doweld, Platylepidium vilujense Kiritchk. (Kiritchkova, 1972, 1985).
Тафофлоры сибирского типа начала средней юры на северо-востоке Сибирской палеофлористической области происходят в основном из якутской свиты (аален–бат) бассейна р. Вилюй. В северо-западной части бассейна якутская свита представлена прибрежно-морскими отложениями с редкими макроостатками растений (р. Марха, левый приток р. Вилюй) и морскими отложениями с обильной фауной аален–батского возраста. Южнее, на западе и юго-западе бассейна, свита сложена континентальными отложениями с многочисленными макроостатками растений (Kirina, 1966). Комплекс растений из опорных разрезов якутской свиты по правому берегу верхнего течения р. Вилюй и нижнего течения р. Марха по-прежнему немногочисленный, но в его составе отсутствуют древние хвощовые Neocalamites и Annulariopsis, а папоротники более разнообразны по сравнению с укугутским комплексом. В состав якутского комплекса растений входят: Equisetites beanii (Bunb.) Sew., Coniopteris cf. hymenophylloides (Brongn.) Sew., Cladophlebis cf. parvifolia Genk., C. vaccensis Ward., Cladophlebis sp., Raphaelia diamensis Sew., R. kirinae Kiritchk., Heilungia sp., Sphenobaiera ex gr. czekanowskiana Heer, Ginkgo ex gr. sibirica Heer, G. cf. tapkensis Dolud. ex Rasskaz., Pseudotorellia cf. nordenskioldii (Nath.) Flor., Сzekanowskia sp., Phoenicopsis irkutensis Dolud. et Rasskaz., Ixostrobus heeri Pryn., Podozamites sp., Pityophyllum sp. (Kiritchkova, 1985).
В целом, оба комплекса растений – укугутский (начало ранней юры) и якутский (начало средней юры), по систематическому составу близки между собой, являясь типичными представителями палеофлоры сибирского типа. Однако их трудно соотнести с западносибирскими тафофлорами. По всей вероятности, это были мобильные первопроходцы, мигрировавшие на северо-восток по Ангаро-Вилюйской аллювиальной равнине вслед за отступавшим Ленским морем (Paleogeografiya…, 1975). Это были предвестники флоры Ленской палеофлористической провинции, сформировавшейся лишь к началу поздней юры.
Подобное впечатление создается и при сравнительном анализе среднеюрских тафофлор Западной Сибири и тафофлор юга Якутии. Тафофлоры Южно-Якутского угольного бассейна, приуроченные к среднеюрским (бат-келловей?) чулымской и дурайской свитам (Vlasov, Marko-vich, 1979), к сожалению, недостаточно изучены, но известный из этих свит список растений также соответствует сибирским тафофлорам. Это уже сформировавшиеся тафоценозы с многочисленными папоротниками (9 видов Coniopteris, 11 видов Cladophlebis, Osmundiella) и цикадофитами (Taeniopteris, Nilssonia, Ctenis, Heilungia, Pterophyllum, Butefia, Jakutiella). По систематическому составу они близки к позднеюрским флорам бассейна р. Буреи Ленской палеофлористической провинции (Vakhrameev, Doludenko, 1961).
По представлениям многих исследователей климат большей части Сибирской палеофлористической области в период ранней и первой половины средней юры был умеренно теплым и влажным, возможно, с сезонными колебаниями температуры и влажности (Vakhrameev et al., 1970; Vakhrameev, 1988; Zhou, 1995; Li Sh.-L. et al., 2014; Na et al., 2017; Li Y.-F. et al., 2019). Об этом свидетельствуют не только седиментологические особенности отложений с частыми угольными залежами, но и систематический состав сибирских тафофлор, а также многочисленные остатки листопадных/веткопадных гинкговых, лептостробовых и хвойных родов Elatocladus, Pityophyllum, Podozamites, встречающиеся наряду с разнообразными папоротниками, в основном осмундовыми и диксониевыми (Vakhrameev, 1962; Zhou, 1995; Sun Ch. L. et al., 2009, 2015a, b; Kiritchkova et al., 2005; Kostina, Herman, 2016; Huang et al., 2016; Deng et al., 2017; Pott, Jiang, 2017). Лишь на юге Западно-Сибирской провинции (южная часть Внутренней Монголии, Китай) и на территории Севeро-Китайской провинции климат был более теплым, приближающимся к субтропическому, о чем свидетельствуют часто присутствующие в составе тафофлор папоротники из семейств Dipteridaceae, Matoniaceae, Marattiaceae, цикадофиты и хвойные с чешуевидными листьями.
ЗАКЛЮЧЕНИЕ
Проводимые с конца прошлого века до настоящего времени исследования по стратиграфии континентальных отложений юры и палеофлористики Сибирской палеофлористической области значительно расширили представления не только об области в целом, но и о практической значимости выделяемых в ее пределах фитостратиграфических комплексов.
Немалую роль в определении возрастного уровня юрских тафофлор Сибирской области в целом и обосновании латеральной корреляции разнофациальных литостратонов сыграла региональная унифицированная комплексная биостратиграфическая шкала юрских отложений Западной Сибири (Kiritchkova et al., 2005), где возраст выделенных фитостратиграфических комплексов обоснован морской фауной.
Результаты проведенного сравнительного анализа систематического состава ранне–среднеюрских тафофлор Сибири и Северного Китая позволяют значительно уточнить положение границы между ранее выделенными на территории области палеофлористическими провинциями – Западно-Сибирской и Северо-Китайской.
Новые данные о систематическом составе тафофлор из юрских отложений ряда местонахождений Северного Китая еще раз убедительно показали, что флора сибирского типа, т.е. Сибирской палеофлористической области, была распространена на значительно бóльшей территории Китая, чем предполагалось ранее.
Палеогеографические особенности северо-востока Сибирской платформы не способствовали широкому распространению западносибирских тафоценозов в этом направлении. Растения, остатки которых приурочены к нижне-среднеюрским, по большей части морским, отложениям бассейна р. Вилюй, рассматриваются нами как первопроходцы, мигрировавшие по Ангаро-Вилюйской аллювиальной равнине на северо-восток вслед за отступавшим Ленским морем. Это были предвестники флоры Ленской палеофлористической провинции, сформировавшейся лишь к концу средней – началу поздней юры.
Климат на территории всей Сибирской палеофлористической области, от северных окраин Западной Сибири и почти до середины Китая, в период ранней и первой половины средней юры был гумидным, с сезонными колебаниями температуры и/или влажности, о чем свидетельствуют систематический состав тафофлор Сибирской области и характер захоронения многочисленных остатков листьев листопадных голосеменных. Такой климат в период наибольшей морской трансгрессии лишь способствовал миграции на север отдельных таксонов тропической флоры Евро-Синийской палеофлористической области вдоль тихоокеанского побережья или по островным дугам (по миграционному “коридору”), проникая вглубь континента.
Список литературы
[Brik] Брик М.И. 1935. Ископаемая флора Южной Ферганы. Ч. I. Ташкент. 36 с.
[Brik] Брик М.И. 1937. Ископаемая флора Южной Ферганы. Ч. II. Ташкент. 74 с.
[Brik] Брик М.И. 1952. Ископаемая флора и стратиграфия нижнемезозойских отложений среднего течения р. Илек в Западном Казахстане. М. 115 с.
[Brik] Брик М.И. 1953. Мезозойская флора Восточно-Ферганского каменноугольного бассейна. М. 111 с.
[Chirva et al.] Чирва С.А., Куликова Н.К., Яковлева С.П. 1997. Новые данные по стратиграфии триаса и юры Печорской синеклизы. – В сб.: Вопросы совершенствования стратиграфической основы фанерозойских отложений нефтегазоносных регионов России. СПб. С. 72–81.
Deng Sh. H., Zhao Y., Lu Y.Z., Shang P., Fan R., Li X., Dong S.X., Liu L. 2017. Plant fossil from the Lower Jurassic coal bearing formation of central Inner Mongolia of China and their implication for palaeoclimate. – Palaeoworld. 26: 279–316.
[Doludenko et al.] Долуденко М.П., Киричкова А.И., Саккулина Г.В., Фокина Н.И. 1989. О флоре боролсайской свиты хребта Каратау (Южный Казахстан). – Палеонтол. журн. 4: 88–100.
[Doludenko, Orlovskaya] Долуденко М.П., Орловская Э.Р. 1976. Юрская флора Каратау. М. 259 с.
[Doludenko, Rasskazova] Долуденко М.П., Рассказова Е.С. 1972. Гинкговые и чекановскиевые Иркутского бассейна. – В кн.: Мезозойские растения (гинкговые и чекановскиевые) Восточной Сибири. М. С. 7–43.
[Frolov, Mashchuk] Фролов А.О., Мащук И.М. 2018. Юрская флора и растительность Иркутского угленосного бассейна. Иркутск. 541 с.
Ge Y.H., Sun C.L., Liu M.X. 2005. Stratigraphic division and correlation of the Yan’an Formation in the northeastern margin of the Ordos basin. – J. Stratigraphy. 31(2): 151–156.
[Genkina] Генкина Р.З. 1966. Ископаемая флора и стратиграфия нижнемезозойских отложений Иссык-Кульской впадины (Северная Киргизия). М. 133 с.
[Genkina] Генкина Р.З. 1977. Стратиграфия юрских континентальных отложений Ферганского хребта и палеоботаническое обоснование их возраста. – Сов. Геология. 9: 63–79.
Heer O. 1876. Beitrӓge zur Jura-Flora Ostsibiriens und des Amurlandes. V. 22. N. 12. St.-Petersbourg. 122 S.
Heer O. 1878. Beitrӓge zur fossilen Flora Sibiriens und des Amurlandes. V. 25. N. 6. St.-Petersbourg. 58 S.
Heer O. 1880. Beitrӓge zur Jura-Flora Ostsibiriens gegrundet auf die von Herrn Richard Maak in Ust-Balei gesammelten Pflanzen. V. 27. N. 10. St.-Petersbourg. 34 S.
Hu Y. 1984. Fossil plants from the original “Huairen Group” in Meiyukou, Datong, Shanxi, and correlation of their age. – Geol. Rev. 30 (6): 569–574.
Huang W., Sun C.L., Wang H.S., Na Y.L., Li Y.F., Li T. 2016. New Phoenicopsis (Czekanowskiales) from the Middle Jurassic Daohugou Biota, China and their roles in phytogeographic and paleoclimatic reconstruction. – Palaeoworld. 25: 388–398.
[Kirina] Кирина Т.И. 1966. Стратиграфия нижнеюрских отложений западной части Вилюйской синеклизы. – В сб.: Геология и нефтегазоносность Западной Якутии. Л. С. 18–71.
[Kiritchkova] Киричкова А.И. 1966. О находке нижнеюрской флоры в Восточной Сибири. – В сб.: Геология и нефтегазоносность Западной Якутии. Л. С. 120–128.
[Kiritchkova] Киричкова А.И. 1972. К систематике Equisetites из юрских и нижнемеловых отложений Западной Якутии. – Бот. журн. 57: 1116–1120.
[Kiritchkova] Киричкова А.И. 1984. Саговниковые и беннеттитовые в юрской и раннемеловой флоре Ленского бассейна – Ежегодник Всесоюзного палеонтологического общества. XXVII: 172–189.
[Kiritchkova] Киричкова А.И. 1985. Фитостратиграфия и флора юрских и нижнемеловых отложений Ленского бассейна. Л. 222 с.
[Kiritchkova] Киричкова А. И. 2011. Триас Западной Сибири: литостратоны опорных разрезов. – Разведка и охрана недр. 4: 27–33.
[Kiritchkova et al.] Киричкова А.И., Батяева С.К., Быстрицкая Л.И. 1992. Фитостратиграфия юрских отложений юга Западной Сибири. М. 216 с.
[Kiritchkova et al.] Киричкова А.И., Быстрицкая Л.И., Травина Т.А. 2002а. Значение Coniopteris и Czekanowskiales для стратиграфии континентальной юры Западной Сибири. – Стратиграфия. Геол. корреляция. 10 (3): 35–52.
[Kiritchkova, Doludenko] Киричкова А.И., Долуденко М.П. 1996. Новые данные по фитостратиграфии юрских отложений Казахстана – Стратиграфия. Геол. корреляция. 4(5): 32–52.
[Kiritchkova et al.] Киричкова А.И., Костина Е.И., Быстрицкая Л.И. 2005. Фитостратиграфия и флора юрских отложений Западной Сибири. СПб. 378 с.
[Kiritchkova et al.] Киричкова А.И., Костина Е.И., Носова Н.В. 2016. Род Sphenobaiera Florin (Ginkgoales) и его значение для стратиграфии юрских отложений Иркутского угленосного бассейна. – Нефтегазовая геология. Теория и практика. 11 (3): 1–34. http:// dx.doi.org/https://doi.org/10.17353/2070-5379/30_2016
[Kiritchkova et al.] Киричкова А.И., Костина Е.И., Носова Н.В. 2017а. Юрские континентальные отложения в разрезах страторегиона Иркутского угленосного бассейна. – Стратиграфия. Геол. корреляция. 25 (5): 17–40.
[Kiritchkova et al.] Киричкова А.И., Костина Е.И., Носова Н.В. 2018. Юрская флора Иркутского угленосного бассейна. – Бот. журн. 103 (1): 36–63.
[Kiritchkova, Nosova] Киричкова А.И., Носова Н.В. 2009. Род Pseudotorellia Florin (Ginkgoales): таксономический и стратиграфический аспекты. – Стратиграфия. Геол. коррел. 17 (6): 59–76.
[Kiritchkova et al.] Киричкова А.И., Носова Н.В., Костина Е.И., Ярошенко О.П. 2020. Континентальная юра Иркутского угленосного бассейна. СПб. 288 с.
[Kiritchkova, Samylina] Киричкова А.И., Самылина В.А. 1979. Об особенностях листьев некоторых мезозойских гинкговых и чекановскиевых. – Бот. журн. 64 (11): 1529–1538.
[Kiritchkova, Travina] Киричкова А.И., Травина Т.А. 1993. О сфеноптероидных папоротниках юры Иркутского бассейна. – Палеонтол. журн. 4: 106–114.
[Kiritchkova et al.] Киричкова А.И., Травина Т.А., Быстрицкая Л.И. 2002b. Род Phoenicopsis. Систематика, история, распространение, значение для стратиграфии. – Биостратиграфия и корреляция фанерозоя нефтегазоносных бассейнов России. Вып. 3. СПб. 205 с.
[Kiritchkova et al.] Киричкова А.И., Ярошенко О.П., Костина Е.И., Носова Н.В. 2017b. Мeтодолoгические основы стратиграфической схемы континентальных отложений Иркутского угленосного бассейна. – Нефтегазовая геология. Теория и практика. 3: 1–21. https://doi.org/10.17353/2070-5379_2017
[Kostina] Костина Е.И. 2004. Юрская флора Канского угленосного бассейна. М. 165 с.
Kostina E.I., Herman A.B. 2013. Middle Jurassic flora of South Mongolia: composition, age and phytogeographic position. – Rev. Palaeobot. Palynol. 93: 82–98.
Kostina E.I., Herman A.B. 2016. Middle Jurassic flora of Mongolia: composition, age and phytogeographic position. – Palaeontol. J. 50 (12): 1437–1450.
Kostina E.I., Herman A.B., Kodrul T.M. 2015. Early Jurassic (possibly Aalenian) Tsagan-Ovoo Flora of Central Mongolia. – Rev. Palaeobot. Palynol. 220: 44–68.
[Krassilov] Красилов В.А. 1972. Мезозойская флора р. Буреи (Ginkgoales и Czekanowskiales). М. 151 с.
[Krassilov] Красилов В.А. 1985. Юрская флора Ошин-Боро-Удзюр-Ула и Жаргаланта (МНР). – В сб.: Юрские континентальные биоценозы Южной Сибири и сопредельных территорий. М. 80–84.
Li B., Hu B. 1984. Fossil plants from the Youngdingzhuang Formation of the Datong Coalfield, notheren Shanxi – Acta Palaeontol. Sinica. 23 (2): 135–147.
Li P., He Y., Wu X., Mei S., Li B. 1988. Early and Middle Jurassic strata and their floras from northeastern border of Qaidam Basin, Qinghai. Nanjing University. 31 (2): 151–156.
Li Sh. L., Yu X.H., Tan C.P. 2014. Jurassic sedimentary evolution of southern Junggar Basin: Implication for palaeoclimate changes in northern Xinjang Uygur Autonomous Region, China. – J. Palaeogeogr. 3 (2): 145–161.
Li T., Li Y.F., Na Y.J., Huang W., Sun Ch. 2014. Discovery of subgenus Phoenicopsis (Gulgoweria) from the Middle Jurassic of Ordos Basin, Inner Mongolia. – Global Geology. 33 (3): 524–534.
Li Y.F., Wang H., Dilcher D.L., Bugdaeva E., Tan. X., Li T., Na Y.L., Sun Ch.L. 2019. Middle Jurassic Plant and Climate in the Ordos Basin, China. – Palaeontol. J. 53 (11): 1216–1235.
[Markovich] Маркович Е.М. 1971. Юрская флора и растительность Орь-Илекского района. Л. 126 с.
[Mogutcheva] Могучева Н.К. 2003. Материалы к стратиграфии триас-среднеюрских отложений Западной Сибири. – В сб.: Проблемы стратиграфии мезозоя Западно-Сибирской плиты. Новосибирск. С. 25–31.
Na Y.L., Sun Ch.L., Wang H.S., Dilcher D.L., Li Y.F., Li T. 2017. A brief introduction to the Middle Jurassic Daohugou Flora from Inner Mongolia, China. – Rev. Palaeobotany, Palynology. 247: 53–67.
[Nosova] Носова Н.В. 2010. Род Leptotoma Kiritch. et Samylina: систематика и особенности эпидермального строения листьев. – Палеоботаника. 1: 22–44.
[Nosova, Kiritchkova] Носова Н.В., Киричкова А.И. 2010. Новый вид рода Pseudotorellia Florin (Ginkgoales) из среднеюрских отложений Восточного Урала. – Бот. журн. 2 (95): 251–256.
[Nosova, Kiritchkova] Носова Н.В., Киричкова А.И. 2018. Новый вид Marskea Florin (Pinopsida) из среднеюрских отложений Иркутского угленосного бассейна (Восточная Сибирь). – Палеонтол. журн. 5: 101–109.
[Nosova et al.] Носова Н.В., Киричкова А.И., Костина Е.И. 2017a. Облиственные побеги и микростробилы хвойных из юрских отложений Иркутского угольного бассейна. – Палеоботаника. 8: 5–27.
Nosova N., van Konijnenburg-van Cittert J.H.A., Kiritchkova A. 2017b. New data on the epidermal structure of the leaves of Podozamites Braun. – Rev. Palaeobot. Palynol. 238: 88–104.
[Nosova et al.] Носова Н.В., Завьялова Н.М., Киричкова А.И., Костина Е.И. 2018. Sorosaccus sibiricus Prynada (Ginkgoales) из среднеюрских отложений Иркутского угленосного бассейна, Восточная Сибирь. – Палеоботаника. 9: 5–17.
[Paleogeografiya SSSR] Палеогеография СССР. Объяснительная записка к Атласу литолого-палеогеографических карт СССР. 1975. Т. 3. Триасовый, юрский и меловой периоды. М. 199 с.
[Popov] Попов М.Г. 1963. Основы флорогенетики. М. 134 с.
Pott C., Jiang B. 2017. Plant remains from the Middle-Late Jurassic Daohugou site of the Yanliao Biota in Inner Mongolia, China. – Acta Palaeobotanica. 57 (2): 185–222.
[Prynada] Принада В.Д. 1962. Мезозойская флора Восточной Сибири и Забайкалья. М. 368 с.
[Samylina] Самылина В.А. 1972. Систематика рода Phoenicopsis. – В кн.: Мезозойские растения (гинкговые и чекановскиевые) Восточной Сибири. М. С. 44–81.
[Samylina, Efimova] Самылина В.А., Ефимова А.Ф. 1968. Первые находки раннеюрской флоры в бассейне р. Колымы. – Докл. АН СССР. 1 (1): 166–168.
[Samylina, Kiritchkova] Самылина В.А., Киричкова А.И. 1991. Род Czekanowskia (Систематика, история, распространение, стратиграфическое значение). Л. 143 с.
[Samylina, Srebrodolskaya] Самылина В.А., Сребродольская И.Н. 1986. Новые виды рода Phoenicopsis из мезозоя Азиатской части СССР. – Бот. журн. 71 (9): 1262–1266.
[Sevryugin et al.] Севрюгин Н.Н., Долуденко М.П., Киричкова А.И., Сакулина Г.В., Федоренко О.А., Фокина Н.И. 1989. Стратиграфия и флора юрских отложений хребта Каратау (Южный Казахстан). М. 58 с.
[Sevryugin et al.] Севрюгин Н.Н., Федоренко О.А., Долуденко М.П., Киричкова А.И., Сакулина Г.В. 1990. Юрские отложения хребта Каратау (Южный Казахстан). – Известия АН СССР. Сер. геол. 7: 63–77.
Seward A.C. 1911. Fossil plants from Chinese Dzungaria, collected by Prof. Obrutschew. – Mem. Com. Geol. St-Petersbourg. 75: 1–61.
[Shurygin et al.] Шурыгин Б.Н., Никитенко Б.Л., Девятов В.П., Ильина В.И., Меледина С.В., Гайдебуров Е.А., Дзюба О.С., Казаков А.М., Могучева Н.К. 2000. Стратиграфия нефтегазоносных бассейнов Сибири. Юрская система. Новосибирск. 480 с.
Sun Ch.L. 1992. The division of the Early Jurassic floristic province of the Eurasia continent. – Journ. Changchun Univ. Earth Sci. Quarterly. P. 178–187.
Sun Ch.L., Dilcher D.L., Wang H., Sun G., Ge Y. 2008. Study of Ginkgo leaves from the Middle Jurassic of Inner Mongolia, China. – Int. J. Plant Sci. 162 (8): 1119–1128.
Sun Ch.L., Dilcher D.L., Wang H., Sun G., Ge Y. 2009. Czekanowskia from the Jurassic of Inner Mongolia, China. – Int. J. Plant Sci. 170 (9): 1183–1194.
Sun Ch.L., Na Y., Dilcher D.L., Wang H., Li T., Li Y. 2015a. A new species of Phoenicopsis Subgenus Windwardia (Florin) Samylina (Czekanowskiales) from the Middle Jurassic of Inner Mongolia, China. – Acta Geologica Sinica. 89 (1): 55–69.
Sun Ch.L., Wang H., Dilcher D.L., Li T., Li Y., Na Y. 2015b. A new species of Czekanowskia (Czekanowskiales) from the Middle Jurassic of Ordos Basin, China. – Botanica Pacifica. 4 (2): 149–155.
Sun G., Miao Y., Chen Y.I. 2006. A new species Sphenobaiera from Middle Jurassic Junggar Basin, Xinjang, China. – J. Jilin University (Earth Science Edition). 36 (5): 717–722.
Sun G., Miao Y., Mosbrugger V., Ashraf A.R. 2010. The Upper Triassic to Middle Jurassic strata and floras of the Junggar basin, Xinjiang, Northwest China. – Palaeobio. and Palaeoenv. 90 (3): 203–214. https://doi.org/10.1007/s12549-010-0039-8
Sze H.C. 1949. Die mesozoische Flora aus der Hsiangchi Kohlen Serie in Westhupeh. – Palaeontol. Sinica. A. 2: 1–71.
Sze H.C. 1956. The fossil flora of the Mesozoic oil-bearing deposits of the Dzungaria-Basin, northwestern Sinkiang. – Acta Palaeontol. Sinica. 4 (4): 461–476.
Sze H.C., Chou T.Y. 1962. Mesozoic Continental Deposits of China. Science Press. Beijing. 180 p.
[Takhtajan] Тахтаджян А.Л. 1970. Происхождение и расселение цветковых растений. Л. 145 с.
[Travina] Травина Т.А. 1996. Систематика рода Leptotoma (Ginkgoales). – Бот. журн. 81 (6): 103–109.
[Vakhrameev] Вахрамеев В.А. 1964. Юрские и раннемеловые флоры Евразии и палеофлористические провинции этого времени. М. 261 с.
[Vakhrameev] Вахрамеев В.А. 1988. Юрские и меловые флоры и климаты Земли. М. 214 с.
Vakhrameev V.A. 1991. Jurassic and Cretaceous floras and climates of the Earth. Cambridge. 318 p.
[Vakhrameev et al.] Вахрамеев В.А., Добрускина И.А., Заклинская Е.Д., Мейен С.В. 1970. Палеозойские и мезозойские флоры Евразии и фитогеография этого времени. М. 424 с.
[Vakhrameev, Doludenko] Вахрамеев В.А., Долуденко М.П. 1961. Верхнеюрская и нижнемеловая флора Буреинского бассейна. М. 135 с.
[Vlasov, Markovich] Власов В.М., Маркович Е.М. 1979. Корреляция юрских и нижнемеловых отложений восточной части Южно-Якутского угольного бассейна. – Сов. Геолог. 1: 72–80.
Volynets E.B., Sun G., Shorokhova S.A., Salyukova E.N. 2020. Early Jurassic flora from South Primorye, Far East, Russia. – Palaeoworld. 29 (3): 590–605. https://doi.org/10.1016/j.palwor.2019.09.002
Wang Y., Mosbrugger V., Zhang H. 2005. Earlly to Middle Jurassic vegetation and climatic events in the Qaidam Basin, Northwest China – Palaeogeogr. Palaeoclimatol. Palaeoecol. 224: 200–216.
Wang Y., Zhang H. 2010. Fertile organs and in situ spores of a new dipteridaceous ferns Hausmannia sinensis from the Jurassic northern China. – Proc. R. Sci. B. 277: 311–320.
Zhang W., Zheng S. 1987. Early Mesozoic fossil plants western Liaoning, Northeast China. – In: Mesozoic stratigraphy and palaeontology of weastern Liaoning. Beijing. P. 239–368.
Zhou Z. 1983. Early Jurassic plants from southwest Hunan, China. – Palaeontol. Sinica. N 165. N S A, (7): 1–85.
Zhou Z. 1995. Jurassic Floras. Chapter 8. – In: Fossil floras of China through the geological ages; Guangzhou. P. 343–410.
Zhou Z., Zhang B. 1998. Tianshia patens gen. et sp. nov., a new type shoots associated with Phoenicopsis from the Middle Jurassic Yima Formation, Henan, China. – Rev. Palaeobot. Palynol. 102: 165–178.
Дополнительные материалы отсутствуют.
Инструменты
Ботанический журнал