

Ботанический журнал, 2022, T. 107, № 11, стр. 1067-1082
ФЛУКТУАЦИИ ЭКОЛОГО-ЦЕНОТИЧЕСКИХ ХАРАКТЕРИСТИК РАСТИТЕЛЬНОСТИ ЛУГОВОЙ КАТЕНЫ (КАРЕЛЬСКИЙ ПЕРЕШЕЕК)
О. В. Созинов 1, *, К. В. Щукина 2, **, А. П. Кораблёв 2, ***, Д. С. Кессель 2, Н. С. Ликсакова 2, М. Ю. Пукинская 2
1 Гродненский государственный университет им. Янки Купалы
230023 Гродно, ул. Ожешко, 22, Республика Беларусь
2 Ботанический институт им. В.Л. Комарова РАН
197376 Санкт-Петербург, ул. Проф. Попова, 2, Россия
* E-mail: ledum@list.ru
** E-mail: vyatka_ks_72@mail.ru
*** E-mail: korablev.anton@yandex.ru
Поступила в редакцию 29.05.2022
После доработки 29.08.2022
Принята к публикации 20.09.2022
- EDN: BWXJWV
- DOI: 10.31857/S0006813622110060
Аннотация
На территории научно-опытной станции “Отрадное” БИН РАН в 2018–2021 гг. проведены геоботанические исследования изменчивости состава и структуры луговых сообществ на пологом склоне террасы озера Отрадное (Приозерский район, Ленинградская область, Россия).
Изученные луговые сообщества относятся к пяти ассоциациям. Вверх по склону террасы сообщества этих ассоциаций закономерно сменяют друг друга: Filipenduletum ulmariae alopecurosum, Phalaridetum arundinaceae filipendulosum, Alopecuretum pratensis anthriscosum, Arrhenatheretum elatioris geraniosum, Calamagrostietum epigeji.
Общее количество выявленных высших растений на профиле за 4 года составило 84 вида при доминирующем участии травянистых растений (68 видов). В среднем видовое богатство луговых фитоценозов составило 20 ± 5 видов/100 м2. Относительно высокая выравненность и видовое богатство формируются в сообществах середины катены. Выявлены следующие закономерности: увеличение роли доминирующих видов в сложении фитоценоза ведет к снижению выравненности (rp = –0.79); увеличение суммарного проективного покрытия ведет к снижению видового богатства сообществ (rp = –0.63). Для изученных сообществ характерна высокая сезонная и разногодичная изменчивость суммарного проективного покрытия и высоты травостоя: коэффициент вариабельности >20%. Количество диагностических видов ассоциаций увеличивается от вершины склона к подножию с 3 до 8. Спектр эколого-ценотических групп диагностических видов луговой катены зависит от положения сообществ на склоне, и, соответственно, от почвенно-грунтовых условий, а также от территориальной близости к фитоценозу определенного типа леса (черноольшанику или сосняку). В целом 60% всех видов травянистых растений в сообществах принадлежат к луговому экоценотипу. Выявлено, что ведущими факторами дифференциации луговых сообществ являются плодородие и увлажнение почв по орографическому градиенту.
Научно-опытная станция Ботанического института им. В.Л. Комарова РАН “Отрадное” (далее – станция) расположена в Приозерском районе Ленинградской области (60°50' с.ш. и 30°15' в.д.). Она находится на Карельском перешейке в 100 км к северу от г. Санкт-Петербурга, в 25 км к югу от г. Приозерска и в 12 км к западу от Ладожского озера.
Земельный участок станции расположен на полуострове, глубоко вдающемся в оз. Отрадное (Пюхе-Ярви). Он неширокой полосой вытянут вдоль берега озера по пологому склону юго-западной экспозиции. В восточной части станции “Отрадное” находится луговой массив, ограниченный с одной стороны сосновым лесом, а с другой – прибрежными черноольшаниками. На базе станции “Отрадное” со второй половины 50-х годов XX века проводились интенсивные экспериментальные геоботанические исследования луговой растительности (Shennikov, 1963; Makarevich, 1963; 1970). В то время основная задача заключалась в проработке теоретических вопросов и научных обоснований основных положений фитоценологии. Программа исследований шла по двум основным направлениям: изучение взаимодействий между растениями при их сосуществовании в ценозе и изучение влияния ценозов и их отдельных компонентов на исходную среду (Makarevich, 1970; Vasilevich, Yurkovskaya, 2007). Наиболее активная фаза работ на луговом стационаре пришлась на 60-е–70-е гг. прошлого века. За период исследований было опубликовано более 70 научных статей по экспериментальной геоботанике (Latmanizova, 2017). Работы в этом направлении продолжались в Отрадном до середины 1990-х годов (Vasilevich, Yurkovskaya, 2007).
С 2018 года сотрудниками лаборатории общей геоботаники БИН РАН возобновлено изучение лугов – заложен эколого-ценотический профиль с 5 пробными площадями (100 м2) на пологом склоне террасы оз. Отрадное юго-западной экспозиции, занятом луговой растительностью.
Исторически луга станции “Отрадное” возникли на месте лесов на озерных террасах; данные по строению почвенного профиля за разные годы (1963–1970), наличие частых дренажных канав на опытном участке, мозаика лесной растительности сравнительно молодого возраста позволяют говорить о длительной истории окультуривания рассматриваемой территории (Shennikov, Makarevich, 1963). До 1938–1940 гг. луга распахивались и засевались многолетними ценными кормовыми растениями, некоторые участки были заняты огородами (Makarevich, 1963). В конце 90-х – начале 2000-х на этих лугах велся активный выпас скота (овец). Интенсивное использование данных лугов прекратилось в начале 2000-х гг.
Луга являются азональными экосистемами, их ценотические особенности определяются локальными, в первую очередь, почвенно-грунтовыми условиями биотопа и режимом антропогенного использования (Rabotnov, 1984; Znamenskiy, 2015). Наиболее эффективный подход к изучению эколого-ценотических характеристик лугов во времени и пространстве основан на использовании понятия “катена”. Катеной считают последовательность взаимозависимых местообитаний, вытянутых цепочкой вдоль геоморфологического профиля с закономерным изменением экологических условий, определяемым рельефом местности (Mordkovich, 1985; Zhukov et al., 2016). Таким образом, катена (= цепочка местообитаний) формируется на градиенте экологических факторов геоморфологического профиля. Рельеф относится к орографической комплексной группе факторов (Walter, 1982). Рельеф меняет режим увлажнения, трофности почвы и иные экологические показатели (эти изменения проявляются на уровне отдельных ландшафтов), и, зачастую, выступает основным комплексным фактором, приводящим к локальной дифференциации растительного покрова (Ramenskiy, 1971). В растительном покрове соответствующие хорологические единицы называют “фитокатенами” (Katenin, 1988; Kholod, 1991; Zaugol’nova, 2001). Использование понятия “катена” в геоботанических исследованиях позволяет в полной мере выразить естественные пространственные и временные свойства экосистем, которые характеризуют их разнообразие и динамику (Didukh, 2008). Катенная организация местообитаний широко распространена на суше: на равнинах, занимающих 2/3 земной суши, 90% площади составляют склоны, в том числе 60% с уклоном поверхности менее 10°. Этого минимального уклона достаточно для локального перераспределения вдоль склона потоков припочвенного воздуха, тепла, атмосферных осадков и грунтовых вод вместе с растворенными в них биогенами, в результате чего формируется катена (Mordkovich, 2017). Поэтому катена является удобной моделью территории, с помощью которой можно оценить изменение видов и экосистем вдоль избранного градиента среды (Kaprus, 2011).
Для изучаемой территории характерен элювиально-аккумулятивный (делювиальный) тип увлажнения. В связи с этим, а также по причине преобладания глинистых и суглинистых почв, при освоении данной территории было проведено поверхностное осушение (мелиоративная сеть по всему склону долины перпендикулярно берегу озера) (Shennikov, Makarevich, 1963). Соответственно, в данных условиях должен проявляться катенный эффект – это реакция растительных сообществ на фактор рельефа, в результате перераспределения влаги и биогенов почв по мезорельефу в рамках конкретной местности.
Многие исследования луговой растительности на различных формах рельефа посвящены многолетнему изучению (мониторингу): временной и пространственной динамики растительных сообществ, оценке состояния и степени загрязнения эдафотопа травяных сообществ; характера внешнего воздействия и степени угроз существованию лугов (Rabotnov, 1987; Guzova, Tikhodeeva, 2016; Stepanovich et al., 2019). Представленная работа раскрывает новые аспекты влияния катенного градиента на состав и структуру луговых экосистем на фоне разногодичной и сезонной изменчивости метеорологических условий с использованием комплекса статистических методов на территории с многолетней историей стационарных наблюдений.
МАТЕРИАЛЫ И МЕТОДЫ
Исследования проводились авторами в течение 2018–2021 гг. в разные периоды вегетационного сезона на территории станции “Отрадное”. По геоботаническому районированию (Geobotanicheskoe…, 1989) территория станции расположена в Балтийско-Ладожском округе среднетаежной подзоны, для растительности которого характерно преобладание сосняков на песчаных водно-ледниковых отложениях. Позднее положение территории было пересмотрено – теперь она относится к южной тайге (Safronova, Yurkovskaya, 2015).
Для характеристики погодных условий с сайтов https://rp5.ru/ и метеостанции Сосново (расположена на 60°33' с.ш., 30°15' в.д.) нами взяты общие (среднегодовые) показатели и показатели для вегетационных сезонов 2018–2021 гг. Для оценки изменения теплообеспеченности территории использовали сумму активных температур (температур воздуха ≥+10°С) и гидротермический коэффициент Селянинова (ГТК) (Selyaninov, 1930) (табл. 1).
Таблица 1.
Метеорологические условия района исследования Table 1. Meteorological conditions of the research area
| Параметр/Parameter | Год/Year | |||
|---|---|---|---|---|
| 2018 | 2019 | 2020 | 2021 | |
| Сумма осадков за год, мм Annual amount of precipitation, mm |
790 | 708 | 753 | 792 |
| Кол-во дней с осадками Number of days with precipitation |
225 | 247 | 234 | 241 |
| Кол-во осадков в период со среднесуточной температурой ≥+10°С, мм Amount of precipitation in the period with an average daily temperature ≥+10 °C, mm |
256 | 177 | 338.8 | 407.4 |
| Сумма осадков за три месяца до проведения описаний, мм Amount of precipitation for three months before the relevés, mm |
184 | 81 | 135 | 190 |
| Кол-во дней с осадками за 3 месяца до проведения описаний Number of days with precipitation for 3 months before the relevés |
42 | 35 | 48 | 46 |
| Среднегодовая температура, t°С Average annual temperature, t°С |
+5.4 | +5.7 | +6.7 | +4.9 |
| Средняя температура за 3 месяца до проведения описаний, t°С Average temperature for 3 months before the relevés, t°C |
+17.1 | +10.9 | +9.2 | +11 |
| Сумма температур за период со среднесуточной температурой ≥+10°С, t°С Sum of temperatures for the period with an average daily temperature ≥+10°C, t°C |
2262 | 1922 | 2048 | 2102 |
| Кол-во дней со среднесуточной температурой ≥+10°С Number of days with an average daily temperature ≥+10°C |
140 | 128 | 139 | 127 |
| ГТК (по Селянинову) Hydrothermal coefficient (according to Selyaninov) |
1.13 | 0.92 | 1.65 | 1.94 |
Изучена изменчивость показателей α- и β-разнообразия луговых сообществ на пологом склоне озерной террасы за 4-летний период (2018–2021 гг.). Для этого на склоне юго-западной экспозиции (азимут 208°) на второй приозерной террасе был заложен профиль длиной 150 м. Вдоль профиля расположены 5 пробных площадей 10 × 10 м (100 м2). Почвы на склоне дерново-слабоподзолистые суглинистые на безвалунных пылеватых суглинках (Baranovskaya, 1963). Ежегодно на каждой пробной площади проводили по одному геоботаническому описанию (сентябрь 2018, июль 2019, июнь 2020, июнь и июль 2021 – всего по 5 описаний на каждой из 5 площадок) с учетом проективного покрытия всех высших растений. Анализ изменчивости видового богатства был проведен по обилию и встречаемости только травянистых видов растений. Мохообразные и древесные виды в оценке строения растительности не использованы вследствие их малочисленности.
Для оценки репрезентативности выборки видов растений на пробных площадях применялся метод (эстиматор) Jackknife2 (bootstrapping: n = 1000) (Sushko, 2021).
Индекс Маргалефа (видового богатства), индекс Пиелу (выравненности) и индекс Бергера–Паркера (доминирования) рассчитывали на основе обилия всех выявленных видов травянистых растений (Magurran, 2004).
Эколого-ценотические группы понимаются нами в соответствии с работой О.В. Смирновой с соавторами (Smirnova et al., 2004) для таежной зоны.
Фитоиндикация экологических режимов биотопов проводилась по шкалам Г. Элленберга (Ellenberg, 1996).
Измерение высот особей доминирующих видов травостоя проводили выборочно на каждой пробной площади с высокой повторностью (от 25 и более) линейкой (±1 мм).
Кислотность почвенного раствора корнеобитаемого горизонта измеряли с помощью Rapitest Digital № 1835 (±0.1) в трехкратной повторности на каждой пробной площади.
Анализ сходства и различий растительных сообществ проводили с помощью Analysis of similarities (ANOSIM) и Permutational multivariate analysis of variance (PERMANOVA) на основе расстояния Брея–Кёртиса (Sushko, 2021). Для выявления диагностических видов сообществ применяли Indicator Value (IndVal) – метод индикаторных видов М. Дюфрена и П. Лежандра (Dufrêne, Legendre, 1997).
Ординация сообществ в системе экологических координат проведена по методу неметрического многомерного шкалирования (nMDS) на основе расстояния Брея–Кёртиса (Hammer et al., 2001). Ординационные оси интерпретированы с помощью анализа корреляций с факторами среды.
Статистическую обработку геоботанических описаний и морфометрических параметров проводили в программе: PAST 4.10 (Hammer et al., 2001) и R v. 4.1.1 (R Core Team, 2021).
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
В период начала экспериментальных исследований на станции “Отрадное” в 1955 г. здесь были распространены полидоминантные мелкозлаково-разнотравные луга с доминированием Agrostis tenuis, Anthoxanthum odoratum, Alopecurus pratensis, Alchemilla monticola. Было отмечено большое сходство флористического состава по всему профилю луга, от основания склона до его вершины (Shennikov, Makarevich, 1963). Причину этого исследователи видели в сглаживании эдафических различий при распашке территории под сеянные луга (Shennikov, Makarevich, 1963).
За прошедшие с начала исследований 60 лет травостой кардинально изменился. На основании анализа полученных данных геоботанических описаний 2018–2021 гг. по луговому фрагменту катены (от опушки прибрежного черноольшаника к вершине склона, занятой сосняком зеленомошным) на основе принципов эколого-фитоценотической классификации (Shennikov, 1941) мы выделили 5 ассоциаций: 1) Filipenduletum ulmariae alopecurosum, 2) Phalaridetum arundinaceae filipendulosum, 3) Alopecuretum pratensis anthriscosum, 4) Arrhenatheretum elatioris geraniosum, 5) Ca-lamagrostietum epigeji, относящихся соответственно к формациям: 1) Filipenduleta ulmariae, 2) Phala-rideta arundinaceae, 3) Alopecureta pratensis, 4) Arrhenathereta elatioris, 5) Calamagrostieta epigeji. Эти формации принадлежат к группам формаций: крупнозлаковые (Magnogramineta) (2–5) и крупноразнотравные луга (Magnoherbeta) (1).
Сообщества первых 3 формаций, относящихся к классу болотистых (Filipenduleta ulmariae, Phalarideta arundinaceae) и настоящих (Alopecureta pratensis) лугов, на территории Северо-Запада не занимают больших площадей, но встречаются часто (Ramenskaya, 1958; Matveeva, 1967; Lopatin, 1971; Vasilevich, Belyaev, 2005). Сообщества этих формаций предпочитают богатые почвы с проточным увлажнением. Таволговые луга возникают на месте вырубленных заболоченных лесов, но являются очень устойчивыми длительно производными сообществами (Vasilevich, Belyaev, 2005). В настоящее время из прежних доминирующих видов свое обилие на части склона сохранил только Alopecurus pratensis L. На обследуемой территории лисохвост луговой мог остаться в травостое от прежних сеяных лугов, поскольку в естественных сообществах он характерен преимущественно для речных пойм (Shennikov, Makarevich, 1963; Vasilevich, Bibikova, 2007). Фитоценозы с таволгой формируются в случае постоянного избытка застойной воды (Matveeva, 1967). Еще один доминант – Arrhenatherum elatius (L.) J. Presl et C. Presl, скорее всего, был занесен на территорию станции “Отрадное” позднее, в результате его посева на экспериментальных площадках (Shennikov, Serafimovich, 1963). Arrhenatherum elatius – европейский вид, внедрялся в качестве кормового и пастбищного растения во многих странах, в 14 регионах на европейской части России вид имеет инвазионный статус (Vinogradova et al., 2015). Тем не менее, ассоциации с доминированием Arrhenatherum elatius достаточно давно описаны в литературе для севера Карельского перешейка (Serafimovich, 1960; Matveeva, 1967). Сообщества с доминированием райграса отличаются достаточно высоким видовым разнообразием (Panasenko, 2019). Формацию Calamagrostieta epigeji описывают на территории Северо-Запада, как типичную при зарастании опушек, вырубок и гарей сосновых лесов (Nitsenko, 1961; Matveeva, 1967). Отмечено, что Calamagrostis epigeios появляется в составе травостоя на лугах при прекращении их хозяйственного использования (Somodi et al., 2008; Samsonova et al., 2016). Таким образом, на обследуемом участке вверх по склону идет смена сообществ болотистых лугов фитоценозами настоящих мезофитных, а выше – бедных мезопсаммофитных лугов.
Порядок смены ассоциаций отражает экологический градиент по склону террасы озера (табл. 2). Вверх по склону отмечено снижение увлажнения и кислотности почвы (pH почвенного раствора изменялось от 6.3 до 6.7); для уровня освещенности и трофности (по азоту) отмечено возрастание показателей в середине профиля. Отмечен невысокий уровень разногодичной изменчивости градаций экологических факторов с тенденцией возрастания их коэффициента вариабельности вверх по склону (табл. 2).
Таблица 2.
Экологические характеристики биотопов ассоциаций (2018–2021 гг.) Table 2. Ecological characteristics of biotopes of the distinguished associations (2018–2021)
| Ассоциация Association |
F | N | R | L | ||||
|---|---|---|---|---|---|---|---|---|
| Балл/Point | Cv | Балл/Point | Cv | Балл/Point | Cv | Балл/Point | Cv | |
| 1 | 5.9 ± 0.05 | 1.9 | 4.3 ± 0.13 | 6.8 | 2.6 ± 0.18 | 14.5 | 5.9 ± 0.08 | 2.9 |
| 2 | 5.1 ± 0.1 | 4.5 | 3.3 ± 0.08 | 5.7 | 2.5 ± 0.14 | 12.5 | 5.5 ± 0.16 | 6.6 |
| 3 | 5.3 ± 0.09 | 3.7 | 4.6 ± 0.23 | 11.5 | 2.9 ± 0.16 | 11.8 | 6.6 ± 0.11 | 3.6 |
| 4 | 5.3 ± 0.11 | 4.6 | 4.8 ± 0.14 | 6.5 | 3.7 ± 0.29 | 17.5 | 6.6 ± 0.16 | 5.3 |
| 5 | 4.5 ± 0.14 | 6.7 | 3.2 ± 0.24 | 16.8 | 3.1 ± 0.15 | 10.8 | 5.4 ± 0.13 | 5.3 |
Примечание. Ассоциации: 1 – Filipenduletum ulmariae alopecurosum; 2 – Phalaridetum arundinaceae filipendulosum; 3 – Alopecuretum pratensis anthriscosum; 4 – Arrhenatheretum elatioris geraniosum; 5 – Calamagrostietum epigeji.
Cv – коэффициент вариабельности, %; F – увлажнение; N – трофность по азоту; R – кислотность; L – освещенность (по Г. Элленбергу).
Note. Associations: 1 – Filipenduletum ulmariae alopecurosum; 2 – Phalaridetum arundinaceae filipendulosum; 3 – Alopecuretum pratensis anthriscosum; 4 – Arrhenatheretum elatioris geraniosum; 5 – Calamagrostietum epigeji.
Cv – coefficient of variation, %; F – moisture; N – soil nitrogen; R – acidity; L – illumination (according to G. Ellenberg).
В среднем на одно геоботаническое описание приходилось 20 ± 5 видов травянистых растений, тогда как на всем профиле отмечено в 3 раза больше видов (табл. 3). Метод Jackknife2 (bootstrapping: n = 1000) показал, что наша выборка выявленных видов растений достаточно репрезентативна и отличается от потенциально возможной в среднем на 2–3 вида, что, скорее всего, связано с сезонной изменчивостью выявляемого видового состава луговых фитоценозов (табл. 3).
Таблица 3.
α-разнообразие луговых сообществ Table 3. α-diversity of meadow communities
| Месяц, год, Мonth, year |
Параметры Parameters | Ассоциация/Association | spp. | spp.** Mean ± SD |
||||
|---|---|---|---|---|---|---|---|---|
| 1 | 2 | 3 | 4 | 5 | ||||
| IX 2018, 2 декада IX 2018, 2nd ten days |
S | 23 | 26 | 19 | 20 | 20 | 47 | 50.8 ± 10.5 |
| H′ | 0.43 | 0.35 | 0.50 | 0.32 | 0.29 | |||
| dМg | 4.61 | 5.17 | 4.05 | 4.08 | 4.20 | |||
| d | 0.33 | 0.46 | 0.34 | 0.55 | 0.63 | |||
| VII 2019, 1 декада VII 2019, 1st ten days |
S | 25 | 26 | 29 | 26 | 21 | 46 | 49.2 ± 8.7 |
| H′ | 0.26 | 0.31 | 0.42 | 0.39 | 0.25 | |||
| dМg | 4.92 | 4.99 | 5.84 | 5.20 | 4.22 | |||
| d | 0.51 | 0.32 | 0.32 | 0.31 | 0.63 | |||
| VI 2020, 3 декада VI 2020, 3rd ten days |
S | 19 | 20 | 23 | 17 | 15 | 33 | 35.4 ± 5.6 |
| H′ | 0.38 | 0.43 | 0.39 | 0.33 | 0.34 | |||
| dМg | 3.72 | 3.60 | 4.48 | 3.17 | 2.89 | |||
| d | 0.46 | 0.25 | 0.36 | 0.41 | 0.58 | |||
| VI 2021, 1 декада VI 2021, 1st ten days |
S | 17 | 18 | 23 | 20 | 25 | 39 | 41.7 ± 7.4 |
| H′ | 0.37 | 0.55 | 0.43 | 0.34 | 0.54 | |||
| dМg | 3.29 | 3.22 | 4.26 | 3.64 | 4.76 | |||
| d | 0.46 | 0.20 | 0.28 | 0.35 | 0.22 | |||
| VII 2021, 1 декада VII 2021, 1st ten days |
S | 22 | 23 | 26 | 18 | 25 | 44 | 47.0 ± 9.1 |
| H′ | 0.37 | 0.28 | 0.50 | 0.29 | 0.42 | |||
| dМg | 4.63 | 4.64 | 4.91 | 3.58 | 5.08 | |||
| d | 0.41 | 0.41 | 0.24 | 0.50 | 0.43 | |||
Примечание. 1–5 – номера ассоциаций (см. Примечание к табл. 2).
S – количество видов в ассоциации; H′ – индекс выравненности; dМg – индекс Маргалефа; d – индекс Бергера–Паркера.
Курсивом отмечены значения индексов, которые достоверно (p < 0.05) отличаются от максимального значения на профиле; spp – общее количество видов в изученных ассоциациях; Mean±SD – среднее арифметическое±квадратичное отклонение; ** метод оценки Jackknife2.
Note. 1–5 –numbers (see Note to Table 2).
S – number of species in association; H′ – evenness index; dМg – Margalef index; d – Berger–Parker index.
Italicized are the index values significantly (p < 0.05) differing from the maximum value on the profile; spp – the total number of species in the studied associations; Mean±SD – arithmetic mean± quadratic deviation; ** Jackknife2 estimation method.
Всего за 4 года выявлено 68 видов травянистых растений. Из видов деревьев отмечены у периметра пробных площадей: Pinus sylvestris L. (1 ед., h = = 5 м), из кустарников – Salix phylicifolia L. (h = = 2.5 м; проективное покрытие – 12%) в асс. Arrhenatheretum elatioris geraniosum и Salix myrsinifolia Salisb. (h = 4 м, 15%) в асс. Phalaridetum arundinaceae filipendulosum. Всходы березы, дуба, ольхи серой и рябины отмечались в отдельные годы, с последующей естественной элиминацией. Выявлено 9 видов мохообразных, количество и обилие которых зависит от сезона описания. Наибольшее количество видов мохообразных и максимальное их обилие выявлено в сентябре, что связано со снижением высоты и обилия травостоя во второй половине вегетационного сезона и, соответственно, снижением конкуренции за свет (табл. 3). Отмечена прямая зависимость суммарного обилия мхов от сезона (с июня по сентябрь): коэффициент корреляции Пирсона r = 0.68; p < 0.05. Чаще всего встречается Brachythecium salebrosum (F. Weber et D. Mohr) Bruch et al.
Относительно высокая выравненность видового состава сообществ устойчиво фиксировалась четыре года в середине профиля (Phalaridetum arundinaceae filipendulosum и Alopecuretum pratensis anthriscosum), что, по мнению В.И. Василевича (Vasilevich, 2015, с. 373), свидетельствует о преобладании в данных сообществах видов со схожей конкурентоспособностью. По видовому богатству (индекс Маргалефа) изученные сообщества, в целом, во многом синхронны с выравненностью (но максимальные значения, как правило, не совпадают). Индекс Маргалефа достаточно тесно связан с видовым богатством (количеством видов, шт./100 м2): r = 0.97 (p < 0.05). Между уровнем выравненности и индексом доминирования (Бергера–Паркера) выявлена обратная корреляционная связь: –0.79 (p < 0.05). Это подтверждает установленный некоторыми исследователями факт снижения биоразнообразия в травяных сообществах доминирующими видами (McCain et al., 2010). В июне 2020 г. все изученные сообщества, как по выравненности, так и по видовому богатству были сходными (табл. 3). Аналогичная ситуация была в сентябре 2018 г. по индексу Маргалефа. Это свидетельствует, что растительные сообщества, относящиеся к различным ассоциациям (и формациям), могут быть схожими по видовому богатству и разнообразию в зависимости от особенностей вегетационного сезона.
По индексу доминирования после стабильно высоких значений в сообществах в верхней части склона (Calamagrostietum epigeji; 2018–2020 гг.), в 2021 году произошло резкое усиление данного показателя в начале июня в асс. Filipenduletum ulmariae alopecurosum с последующим возвратом высоких значений индекса Бергера–Паркера в июле к сообществам верхней части профиля.
Все вышеприведенные результаты по α-разнообразию лугов подтверждают классические положения луговедения о значительной сезонной и разногодичной изменчивости, как значимости доминирующих видов, так и видового богатства в луговых сообществах (Shennikov, 1941).
Сравнение полученных нами данных с результатами В.И. Василевича (Vasilevich, 2015) по влажным лугам Европейской России показало достаточно высокое сходство по видовому богатству, суммарному проективному покрытию и выравненности асс. Filipenduletum и Alopecuretum (табл. 3, рис. 1В). Сравнение с данными по видовому разнообразию пойменных лугов р. Вятки (Shchukina, 2017) демонстрирует некоторые отличия. Для пойменных сообществ асс. Phalaridetum arundinaceae характерны сходные значения выравненности при меньшем видовом богатстве и суммарном проективном покрытии. Для асс. Filipenduletum и Alopecuretum при достаточно близких значениях видового богатства и суммарного проективного покрытия, выравненность в пойменных сообществах выше.
Рис. 1.
Морфо-ценотические параметры травостоя сообществ.
1–5 – номера ассоциаций (см. Примечание к табл. 2).
A – H, cm – высота травостоя; B – ∑ПП, % – суммарное проективное покрытие травостоя.
Fig. 1. Morpho-coenotic parameters of herb layer of communities.
1–5 – association numbers (see Note to Table 2).
A – H, cm – height of herb layer; B – ∑ПП, % – total projective cover of herb layer.
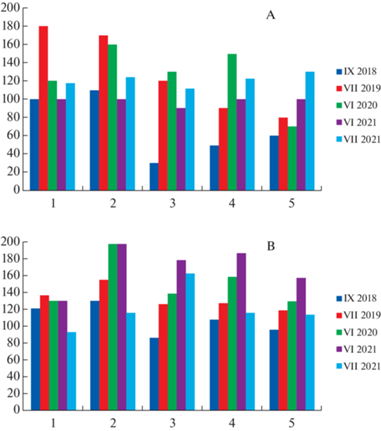
Морфо-ценотические характеристики растительных сообществ также являются важными индикаторами особенностей экологических режимов биотопов и конкурентных взаимоотношений между видами растений. В большинстве описаний луговых фитоценозов суммарное проективное покрытие стабильно выше 100% (рис. 1В), т.е. в сообществах имеется значительное перекрывание надземных частей растений, что ведет к усилению конкуренции за свет и снижению видового разнообразия (Vasilevich, 2015). Согласованная изменчивость объема травостоя (произведение суммарного проективного покрытия на высоту вегетирующей части) с видовым богатством и выравненностью недостоверна, аналогичные данные получены В.И. Василевичем (Vasilevich, 2015) для влажных лугов Северо-Запада. Нами установлена обратная средняя корреляционная связь между суммарным проективным покрытием и значением индекса Маргалефа (видового богатства): r = –0.63, p < 0.05, а также – обратная значимая корреляция согласованной изменчивости объема травостоя с индексом Маргалефа: r = –0.47 (p < 0.05), что, опять-таки, подтверждает мнение В.И. Василевича (Vasilevich, 2015).
Отмечено достоверное отличие высот вегетативной части побегов доминирующих растений асс. Filipenduletum ulmariae alopecurethosum (у основания склона) от высот вегетативных частей растений всех остальных сообществ. По высоте генеративных побегов значимо выделяется асс. Alopecuretum pratensis anthriscosum (табл. 4).
Таблица 4.
Различия и сходства по высотам генеративных побегов и их вегетативной части травостоя луговых сообществ (p-level по критерию наименьшего существенного различия (LSD-test); 2021 г.) Table 4. Differences and similarities in the heights of generative shoots and their vegetative part of the herb layer of meadow communities (p-level by the least significant difference (LSD-test); 2021)
| Параметр Parameter | Длина генеративных побегов/Length of generative shoots | |||||
|---|---|---|---|---|---|---|
| 5 | 4 | 3 | 2 | 1 | ||
| Длина вегетативной части побегов Length of vegetative part of shoots |
5 | 0.1 | 0.00003 | 0.41 | 0.07 | |
| 4 | 0.9 | 0.016 | 0.76 | 0.51 | ||
| 3 | 0.79 | 0.74 | 0.043 | 0.31 | ||
| 2 | 0.73 | 0.71 | 0.88 | 0.43 | ||
| 1 | 0.001 | 0.0022 | 0.0006 | 0.0081 | ||
Сравнение степени изменчивости высоты травостоя и суммарного проективного покрытия показало высокую сезонную и разногодичную (2018–2021 гг.) вариабельность данных параметров: Cv >20% (рис. 1). Высота вегетативной части травостоя изменялась от 30 до 180 см (Cv = = 32.6%), высота генеративных побегов – 110–200 см (Cv = 20.2%), суммарное проективное покрытие – 87–198% (Cv = 22.1%).
Выявлен общий сезонный тренд (июнь–сентябрь) изменчивости суммарного проективного покрытия травостоя: r = –0.64 (p < 0.05), что говорит о наиболее высокой степени конкуренции в начале лета, с последующим снижением до минимума осенью. Сезонное изменение высоты травостоя (июнь, июль, сентябрь) имеет колоколообразную (арочную) нелинейную тенденцию с максимумом: по генеративной высоте в июле и вегетативной части травостоя – во второй половине июня (рис. 1А), что считается закономерным фенологическим явлением.
Приведенные факты свидетельствуют о том, что луга являются довольно лабильными сообществами по видовому богатству и разнообразию, а также по морфо-ценотическим параметрам на экологических градиентах, что дает возможность формировать относительно устойчивые долговременные фитоценозы при высокой изменчивости надземной структуры.
Сравнительный анализ геоботанических описаний показал, что ведущую роль в дифференциации растительных сообществ играет комплексный фактор рельефа, а не погодные условия разных лет. Достоверное различие (по ANOSIM) между сериями описаний разных лет выявлено только для максимально удаленных по времени геоботанических данных: первая декада июня – вторая декада сентября (R = 0.44; p = 0.0066). Не отмечено различий между июньскими и июльскими описаниями одного года (2021 г., p = 0.44). В то же время между всеми ассоциациями по ANOSIM и PERMANOVA получены высокозначимые различия (p ≤ 0.01 с поправкой Bonferroni; табл. 5).
Таблица 5.
β-разнообразие луговых сообществ (p-level) Table 5. β-diversity of meadow communities (p-level)
| Test | ANOSIM | |||||
|---|---|---|---|---|---|---|
| 1 | 2 | 3 | 4 | 5 | ||
| PERMANOVA | 1 | 0.0073 | 0.008 | 0.0075 | 0.0087 | |
| 2 | 0.0074 | 0.0095 | 0.0099 | 0.009 | ||
| 3 | 0.0097 | 0.008 | 0.0079 | 0.0078 | ||
| 4 | 0.0083 | 0.0081 | 0.0089 | 0.01 | ||
| 5 | 0.0075 | 0.0097 | 0.0085 | 0.0088 | ||
Количество диагностических видов в ассоциациях с достоверным вкладом в их различие закономерно уменьшается вверх по склону, т.е. происходит снижение числа индикаторных видов растений, связанных с определенными условиями местообитаний. Количество видов со значением IndVal >50% снижается по мере уменьшения степени увлажнения и возрастания pH (см. табл. 2), а также уменьшается до одного количество индикаторных видов злаков. На более сухих участках катены (асс. Calamagrostietum epigeji) весомое индикаторное значение имеют только три вида (табл. 6).
Таблица 6.
Диагностические виды ассоциаций (Indicator Value; p < 0.05) Table 6. Diagnostic species of associations (Indicator Value; p < 0.05)
| Ассоциация/Association | ||||
|---|---|---|---|---|
| 1 | 2 | 3 | 4 | 5 |
| Equisetum sylvaticum L.* | Phalaris arundinacea L.* | Anthriscus sylvestris (L.) Hoffm.* | Cirsium hetero- phyllum (L.) Hill* | Calamagrostis epigeios (L.) Roth* |
| Scirpus sylvaticus L.* | Galium boreale L.* | Angelica sylvestris L.* | Arrhenatherum elatius (L.) J. Presl et C. Presl* | Trifolium medium L.* |
| Filipendula ulmaria (L.) Maxim.* | Lupinus polyphyllus Lindl.* | Alchemilla vulgaris L. s.l. | Geranium pratense L. | Hypericum maculatum Crantz |
| Rumex crispus L.* | Lysimachia vulgaris L.* | Equisetum arvense L. | Alchemilla vulgaris L. s.l. | – |
| Elytrigia repens (L.) Nevski | Deschampsia cespitosa (L.) P. Beauv. | Geum rivale L. | Angelica sylvestris L. | – |
| Veronica chamaedrys L. | Scirpus sylvaticus L. | Alopecurus pratensis L. | – | – |
| Geum rivale L. | Equisetum pratense Ehrh. | Lathyrus pratensis L. | – | – |
| Poa nemoralis L. | – | Aegopodium podagraria L. | – | – |
Примечание. 1–5 – номера ассоциаций (см. Примечание к табл. 2). Полужирным шрифтом выделены злаки. Виды расположены в столбцах сверху вниз по мере уменьшения значения IndVal; * – значения IndVal >50%.
Note. 1–5 – association numbers (see Note to Table 2). Grasses are highlighted in bold. The species are arranged in the columns from top to bottom in decreasing order of the IndVal value; * – IndVal values >50%.
Изменчивость спектра эколого-ценотических групп индикаторных видов растений по профилю адекватно отражает экологические особенности луговой части катены (рис. 2). Луговые виды становятся доминирующей группой среди индикаторных растений в середине профиля и выше. Это подтверждает тезис о лугах, как о достаточно стабильных экосистемах, которые сохраняют свои ценотические характеристики в течение длительного времени (Cherednichenko, Borodulina; 2018). В асс. Filipenduletum ulmariae alopecurethosum и Phalaridetum arundinaceae filipendulethosum явно прослеживается влияние расположенного рядом черноольшаника – среди индикаторных наблюдается значимое участие водно-болотных (Phalaris arundinacea, Scirpus sylvaticus, Lysimachia vulgaris) и нитрофильных (Filipendula ulmaria, Geum rivale, Elytrigia repens) видов. Наличие нитрофильных индикаторных видов в большинстве ассоциаций профиля отражает элювиально-аккумулятивный (делювиальный) тип увлажнения. В асс. Calamagrostietum epigeji ключевым индикаторным видом становится вейник наземный – боровой вид (влияние близлежащего сосняка). Соответственно, в луговых сообществах есть проявление экотонного эффекта в спектрах эколого-ценотических групп индикаторных видов растений в зависимости от близлежащего лесного фитоценоза. Значимая доля среди диагностических видов большинства ассоциаций катены – видов широколиственных лесов (неморальные виды) и черноольшаников (нитрофильные виды) свидетельствует о потенциальных перспективах данных луговых ассоциаций к сукцессионным изменениям в сторону формирования лесных фитоценозов.
Рис. 2.
Спектры эколого-ценотических групп индикаторных видов растений луговых ассоциаций.
Эколого-ценотические группы: Pn – боровая (виды сосновых лесов), Nm – неморальная (виды широколиственных лесов), Md – лугово-степная (виды лугов, луговых степей и лесных опушек), Wt – водно-болотная (виды низинных болот, прибрежно-водные, внутриводные и виды свежего аллювия), Nt – нитрофильная (виды черноольшаников).
По оси абсцисс – номера ассоциаций (см. табл. 2), по оси ординат – доля индикаторных видов эколого-ценотических групп (%).
Fig. 2. Spectra of ecological and coenotic groups of indicator plant species of meadow associations.
Ecological and coenotic groups: Pn – pinery (species of pine forests), Nm – nemoral (species of broad-leaved forests), Md – meadow-steppe (species of meadows, steppes and forest edges), Wt – wetland (species of lowland swamps, coastal-aquatic, aquatic and growing on fresh alluvium), Nt – nitrophilic (species of black alder forests).
X-axis – the numbers of associations (see Table 2), Y-axis – the share of indicator species of ecological and coenotic groups (%).
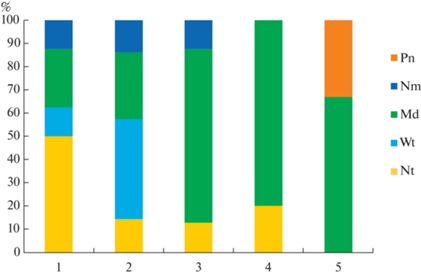
Полные спектры эколого-ценотических групп каждой ассоциации на протяжении 4-х лет достаточно устойчивы – колебания в каждой группе ±1–2 вида. Различия по эколого-ценотическим группам между ассоциациями более значительны: количество видов каждой эколого-ценотической группы отличалось до ±8–9 видов. В целом, спектр эколого-ценотических групп всего видового списка свидетельствует, что открытые сообщества катены относятся к влажным лугам на плодородных почвах: почти 60% травянистых видов относятся к луговой (мезофитной) группе, 18% – водно-болотные виды, 13% – нитрофилы.
Ординация (nMDS) растительных сообществ подтвердила результаты анализа ANOSIM и PERMANOVA: разногодичные отличия между растительными сообществами минимальны, а основные дифференцирующие факторы ассоциаций – локальные факторы катены (рис. 2). Это подтверждает предположение А.П. Шенникова и В.Н. Макаревич (Shennikov, Makarevich, 1963), что со временем выравненность сообществ исследуемых лугов должна уменьшаться, эдафические различия увеличиваться; вместе с тем должны становиться более четкими и различия в растительности по склону. Уменьшение видового богатства и разнообразия (по индексу Маргалефа и выравненности) в ходе сукцессии показано, например, в 21-летнем опыте восстановления растительности влажных лугов низменности Саньцзян (Сунгарийская равнина) на северо-востоке Китая (An et al., 2019).
Ординация, как в случае анализа луговой растительности катены за каждый год, так и в случае анализа описаний одной ассоциации, выявляет два примерно равных по значимости градиента (рис. 3). Первая ось объясняет 31.4% общей изменчивости, вторая – 41.2% общей изменчивости (рис. 3). Первая ось значимо коррелирует с параметрами кислотности почв (табл. 6). Таким образом, главным градиентом в данной совокупности, несомненно, оказывается фактор плодородия почв (кислотность), носящий, возможно, во многом антропогенный характер (кошение, поверхностное осушение, выпас скота) и естественные причины (сток воды по суглинкам склона). Вторая ось значимо коррелирует со шкалами почвенного увлажнения, содержания азота и освещенности (по Элленбергу). Набор двух ведущих факторов среды (плодородие и увлажнение почв) типичен для лугов и является определяющим для луговой растительности (Znamenskiy, 2003; Havlovà et al., 2004; Borodulina et al., 2019). Корреляции ординационных осей с погодными условиями низкие и недостоверные (p ≥ 0.6). Ординация показала ведущий вклад в дифференциацию луговых сообществ почвенно-грунтовых условий (r2 = 0.8722; p = 0.0009), тогда как погодные условия сезонов вносят минимальный вклад (r2 = 0.0306; p = 0.98).
Рис. 3.
Ординационная диаграмма nMDS сообществ луговой части катены.
А – Положение ассоциаций в системе экологических координат (различия ассоциаций); В – Положение луговых сообществ катены 2018–2021 гг. в системе экологических координат (разногодичные различия растительности катены).
Векторы факторов соответствуют экологическим шкалам (факторы фитоиндикации см. табл. 2); pH – кислотность почвенного раствора; нумерация точек: год_выполнения описания_номер ассоциации (см. табл. 2); ┼ – центроид; stress = 0.148.
Fig. 3. Ordination diagram of nMDS communities of the meadow part of the catena.
А – position of associations in the system of ecological coordinates (differences of associations); В – position of meadow communities of the catena 2018–2021 in the system of ecological coordinates (differences of vegetation of the catena in different years).
The factor vectors correspond to ecological scales (for factors of phytoindication see Table. 2); pH – acidity of soil solution. Numbering of points: year of relevé (last two digits)_number of association (see Table 2); ┼ – centroid; stress = 0.148.
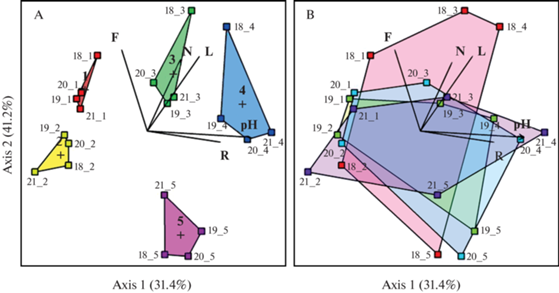
Отмечено, что вдоль градиента содержания азота, увлажнения и освещенности в наибольшей степени к осени изменяются асс. Filipenduletum ulmariae alopecurethosum (1), Alopecuretum pratensis anthriscosum (3) и Arrhenatheretum elatioris geraniethosum (4) (рис. 3В), тогда как асс. Phalaridetum arundinaceae filipendulethosum (2) и Calamagrostietum epigeji (5) достаточно стабильны в течение вегетационного сезона.
Таким образом, анализ геоботанических данных растительных сообществ луговой части катены береговой террасы озера Отрадное (Ленинградская обл.) за четыре вегетационных сезона показал, что:
– луговая растительность сформирована сообществами пяти ассоциаций, экологически закономерно сменяющих друг друга по склону террасы озера;
– видовое богатство фитоценозов составляет 20 ± 5 видов/100 м2, тогда как общее количество видов травянистых растений катены – 68; по видовому богатству и разнообразию луговые сообщества катены сходны с сообществами соответствующих ассоциаций европейской части России;
– относительно высокая выравненность и видовое богатство формируются в сообществах середины катены; увеличение роли доминирующих видов в сложении фитоценоза ведет к снижению выравненности (r = –0.79); увеличение суммарного проективного покрытия лугового сообщества ведет к снижению видового богатства сообществ (r = –0.63);
– отмечена высокая сезонная и разногодичная изменчивость параметров надземной части травостоя (суммарное проективное покрытие, высота травостоя): Сv >20%;
– количество диагностических видов ассоциаций уменьшается вверх по склону с 8 до 3; спектр эколого-ценотических групп во многом зависит от положения сообществ на склоне, и соответственно, от почвенно-грунтовых условий, а также от соседства с сообществами определенного типа леса (черноольшаником или сосняком), но в целом 60% видов, слагающих растительность открытой части катены, составляют виды лугового экоценотипа;
– достоверно установлено, с использованием разнообразных статистических методик, наличие на исследуемых лугах катенного эффекта; основными факторами среды, дифференцирующими луговые сообщества, являются эдафические условия, изменяющиеся на орографическом профиле.
Список литературы
An Y., Gao Y., Liu X.H., Tong S.Z. 2019. Interactions of soil moisture and plant community properties in meadows restored from abandoned farmlands on the Sanjiang Plain, China. – Community Ecology. 20 (1): 20–27: 1585–8553. https://doi.org/10.1556/168.2019.20.1
[Baranovskaya] Барановская А.В. 1963. Почвенный покров научно-опытной станции “Отрадное”. – Комплексные экспериментальные геоботанические исследования. М., Л. С. 12–24.
[Borodulina et al.] Бородулина В.П., Комарова А.Ф., Чередниченко О.В. 2019. Наземновейниковые луга в охранной зоне Полистовского заповедника (Псковская область). – Разнообразие растительного мира. 1 (1): 44–61. https://doi.org/10.22281/2686-9713-2019-1-44-61
Cherednichenko O., Borodulina V. 2018. Biodiversity of herbaceous vegetation in abandoned and managed sites under protection regime: a case study in the Central Forest Reserve, NW Russia. – Hacquetia. 17 (1): 35–59. https://doi.org/10.1515/hacq-2017-0015
[Didukh] Дідух Я.П. 2008. Етюди фітоекології. Киiв. 286 с.
Dufrêne M., Legendre P. 1997. Species Assemblages and Indicator Species: The Need for a Flexible Asymmetrical Approach. – Ecological monographs. 67: 345–366. https://doi.org/10.2307/2963459
Ellenberg H. 1996. Vegetation Mitteleuropas mit den Alpen in okologischer, dynamischer und historischer Sicht. 5. Aufl. Ulmer, Stuttgart. 1096 s.
[Geobotanicheskoe…] Геоботаническое районирование Нечерноземья европейской части РСФСР. 1989. Л. 64 с.
[Guzova, Tikhodeeva] Гузова Т.А., Тиходеева М.Ю. 2016. Преобразования биогеоценозов суходольных лугов в процессе зарастания ольхой серой (Alnus inсаnа (L.) Moench). – Вестник Санкт-Петербургского университета. Сер. 3. Биология. 1: 4–20.
Hammer, Øyvind, Harper, David A.T., Paul D. Ryan. 2001. Past: Paleontological Statistics Software Package for Education and Data Analysis. – Palaeontologia Electronica. 4 (1): art. 4: 9 p. 178 kb. http://palaeo-electronica.org/2001_1/past/issue1_01.htm.
Havlovà M., Chytrý M., Tichý L. 2004. Diversity of hay meadows in the Czech Republic: major types and environmental gradients. – Phytocoenologia. 34 (4): 551–567. https://doi.org/10.1127/0340-269X/2004/0034-0551
[Kholod] Холод С.С. 1991. Фитокатены в растительном покрове горных территорий (на примере тундровой зоны северо-востока СССР). – Бот. журн. 76 (1): 42–51.
[Kaprus] Капрусь І.Я. 2011. Порівняльний аналіз фаун Collembola на території України. – Біологічні Студії. 5 (3): 135–154.
[Katenin] Катенин А.Е. 1988. Классификация неоднородных территориальных единиц растительного покрова на примере растительности тундровой зоны. – Бот. журн. 73 (2): 186–197.
[Latmanizova] Латманизова Т.М. 2017. Научно-опытной станции “Отрадное” Ботанического института им. В.Л. Комарова РАН – 70 лет. – Hortus bot. 12: 4–43. https://doi.org/10.15393/j4.art.2017.4105
[Lopatin] Лопатин В.Д. 1971. Краткий очерк луговой растительности Северного Приладожья. – Очерки по растительному покрову Карельской АССР. Петрозаводск. С. 20–59.
McCain K.N.S., Baer S.G., Blair J.M., Wilson J.W.T. 2010. Dominant Grasses Suppress Local Diversity in Restored Tallgrass Prairie. – Restoration Ecology. 18 (1): 40–49. https://doi.org/10.1111/j.1526-100X.2010.00669.x
Magurran A.E. 2004. Measuring biological diversity. Oxford, UK. 256 p.
[Makarevich] Макаревич В.Н. 1963. Влияние на луговой травостой способов его использования. – Комплексные экспериментальные геоботанические исследования. М., Л. С. 39–103.
[Makarevich] Макаревич В.Н. 1970. Растительный покров. – Луговой фитоценоз и его динамика в зависимости от различных мер воздействия. Л. С. 24–28.
[Matveeva] Матвеева Е.П. 1967. Луга Советской Прибалтики. 355 с.
[Mordkovich] Мордкович В.Г. 1985. Степные катены. Новосибирск. 118 с.
[Mordkovich] Мордкович В.Г. 2017. Сопряженность экологических сукцессий с катенной организацией пространства. – Журнал общей биологии. 78 (2): 32–46.
[Nitsenko] Ниценко А.А. 1961. Изменение естественной растительности Ленинградской области под воздействием человека. Л. 51 с.
[Panasenko] Панасенко Н.Н. 2019. Arrhenatherum elatius (L.) J. Presl et C. Presl в Брянской области. – Разнообразие растительного мира. 3 (3): 26–38. https://doi.org/10.22281/2686-9713-2019-3-26-38
R Core Team. 2021. R: A language and environment for statistical computing. R Foundation for Statistics. Computing, Vienna, Austria. http://www.r-project.org/index.html
[Rabotnov] Работнов Т.А. 1984. Луговедение. М. 320 с.
[Rabotnov] Работнов Т.А. 1987. Экспериментальная фитоценология: Учеб.-метод. пособие. М. 160 с.
[Ramenskaya] Раменская М.Л. 1958. Луговая растительность Карелии. Петрозаводск. 400 с.
[Ramenskiy] Раменский Л.Г. 1971 Избранные работы. Проблемы и методы изучения растительного покрова. Л. 334 с.
[Safronova, Yurkovskaya] Сафронова И.Н., Юрковская Т.К. 2015. Зональные закономерности растительного покрова равнин Европейской России и их отображение на карте. – Бот. журн. 100 (11): 1121–1141.
[Samsonova et al.] Самсонова В.П., Кондрашкина М.И., Кротов Д.Г. 2016. Вейник наземный (Calamagrostis epigeios Roth.) как индикатор зарастающих территорий в нечерноземной зоне России. – Проблемы агрохимии и экологии. 2: 34–39.
[Selyaninov] Селянинов Г.Т. 1930. К методике сельскохозяйственной климатографии. – Труды по сельскохозяйственной метеорологии. XXII (2): 45–91.
[Serafimovich] Серафимович Н.Б. 1960. Изучение биологической разнородности популяции райграса высокого – Arrhenatherum elatius (L.) М. et К. – Тр. БИН АН СССР. Сер. III. Геоботаника. Вып. 12. С. 196–210.
[Shchukina] Щукина К.В. 2017. Видовое разнообразие лугов поймы р. Вятки. – Бот. журн. 102 (10): 1420–1436. https://doi.org/10.1134/S0006813617100040
[Shennikov] Шенников А.П. 1941. Луговедение. Л. 512 с.
[Shennikov] Шенников А.П. 1963. Задачи лаборатории экспериментальной геоботаники Ботанического института Академии наук СССР. – Комплексные экспериментальные геоботанические исследования. М., Л. С. 6–11.
[Shennikov, Makarevich] Шенников А.П., Макаревич В.Н. 1963. Краткий очерк природной флоры и растительности территории научно-опытной станции “Отрадное”. – Комплексные экспериментальные геоботанические исследования. М., Л. С. 33–38.
[Shennikov, Serafimovich] Шенников А.П., Серафимович Н.Б. 1963. О влиянии площади питания на райграс высокий (Arrhenatherum elatius (L.) М. et К.). – Комплексные экспериментальные геоботанические исследования. М., Л. С. 208–226.
[Smirnova et al.] Смирнова О.В., Ханина Л.Г., Смирнов В.Э. 2004. Эколого-ценотические группы в растительном покрове лесного пояса Восточной Европы. – Восточноевропейские леса: история в голоцене и современность. Кн. 1. М. С. 165–175.
Somodi I., Viragh K., Janos P. 2008. The effect of expansion of the clonal grass Calamagrostis epigejos on the species turnover of a semi-arid grassland. – Appl. Veg. Sci. 11: 187–192.
[Stepanovich et al.] Степанович И.М., Степанович Е.Ф., Зеленкевич Н.А. 2019. Мониторинг луговой и лугово-болотной растительности. – Мониторинг растительного мира в Республике Беларусь: результаты и перспективы. Минск. С. 23–70.
[Sushko] Сушко Г.Г. 2021. Программное обеспечение PAST – как инструмент анализа данных в фаунистических исследованиях. – Изв. Гомельского гос. ун-та им. Ф. Скорины. Сер. Естественные науки. 3 (126): 55–61.
[Vasilevich] Василевич В.И. 2015. Видовое разнообразие влажных лугов Европейской России. – Бот. журн. 100 (4): 372–381.
[Vasilevich, Belyaev] Василевич В.И., Беляев Е.А. 2005. Таволговые луга Северо-Запада Европейской России. – Бот. журн. 90 (12): 1801–1813.
[Vasilevich, Bibikova] Василевич В.И., Бибикова Т.В. 2007. Щучковые и лисохвостные луга Европейской России. – Бот. журн. 92 (1): 29–41.
[Vasilevich, Yurkovskaya] Василевич В.И., Юрковская Т.К. 2007. Александр Петрович Шенников. 1888–1962. М. 114 с.
[Vinogradova et al.] Виноградова Ю.К., Абрамова Л.М., Акатова Т.В., Аненхонов О.А., Анкипович Е.С., Антипова Е.М., Антонова Л.А., Афанасьев В.Е., Багрикова Н.А., Баранова О.Г., Борисова Е.А., Борисова М.А., Бочкин В.Д., Буланый Ю.И., Верхозина А.В., Владимиров Д.Р., Григорьевская А.Я., Ефремов А.Н., Майоров С.Р., Зыкова Е.Ю., Кравченко А.В., Крылов А.В., Куприянов А.Н., Лавриненко Ю.В., Лактионов А.П., Лысенко Д.С., Меньшакова М.Ю., Мещерякова Н.О., Мининзон И.Л., Михайлова С.И., Морозова О.В., Нотов А.А., Панасенко Н.Н., Пликина Н.В., Пузырев А.Н., Раков Н.С., Решетникова Н.М., Антипова С.В., Сагалаев В.А., Силаева Т.Б., Силантье-ва М.М., Стародубцева Е.А., Степанов Н.В., Стрельникова Т.О., Терехина Т.А., Тремасова Н.А., Третьякова А.С., Хорун Л.В., Чернова О.Д., Шауло Д.Н., Эбель А.Л. 2015. “Черная сотня” инвазионных растений России. – Совет ботанических садов стран СНГ при Международной ассоциации академий наук. 4 (27): 85–89.
[Walter] Вальтер Г. 1982. Общая геоботаника. М. 264 с.
[Zaugol’nova] Заугольнова Л.Б. 2001. Характеристика лесных фитокатен в подзоне хвойно-широколиственных лесов. – Бюл. МОИП. Отд. биол. 106 (5): 42–50.
[Zhukov et al.] Жуков А.В., Кунах О.Н., Новикова В.А., Ганжа Д.С. 2016. Фитоиндикационное оценивание катены сообществ почвенной мезофауны и их экоморфическая организация. – Бiологiчний вiсник МДПУ ім. Богдана Хмельницького. 6 (3): 91–117. https://doi.org/10.15421/201676
[Znamenskiy] Знаменский С.Р. 2003. Экологическая структура мезофитных луговых сообществ Заонежья (Карелия): Автореф. дис. … канд. биол. наук. М. 25 с.
[Znamenskiy] Знаменский С.Р. 2015. Растительность ксеромезофитных и мезофитных лугов среднетаежной Карелии: эколого-топологический подход. – Труды Карельского научного центра РАН. 2: 3–15. https://doi.org/10.17076/eco40
Дополнительные материалы отсутствуют.
Инструменты
Ботанический журнал