

Ботанический журнал, 2022, T. 107, № 11, стр. 1055-1066
МОРФОЛОГИЧЕСКАЯ ИЗМЕНЧИВОСТЬ PALIMBIA REDIVIVA s.l. И СТАТУС P. TURGAICA (APIACEAE)
Т. А. Остроумова *
Московский государственный университет им. М.В. Ломоносова, биологический факультет, ботанический сад
119991 Москва, Ленинские Горы, 1, стр. 12, Россия
* E-mail: ostro_t_a@mail.ru
Поступила в редакцию 17.02.2022
После доработки 13.09.2022
Принята к публикации 20.09.2022
- EDN: DFZDBV
- DOI: 10.31857/S0006813622110059
Аннотация
Проведен критический анализ применимости предложенных ранее разными авторами морфологических диагностических признаков для разграничения наиболее распространенных видов Palimbia, P. rediviva и P. turgaica (число лучей терминального зонтика, угол между главным стеблем и ветвями, характер соцветия, порядок ветвления, длина плодоножек, ширина сегментов листа, число междоузлий на стебле). Изучены типовые образцы Palimbia rediviva и P. turgaica и коллекции обоих видов из гербариев LE, MHA и MW (всего 934 особи) методами одномерной и многомерной статистики. Показано, что указанные признаки имеют одновершинное распределение и не годятся для разграничивания видов, следовательно, название P. turgaica следует считать синонимом P. rediviva. Вместе с тем отмечена некоторая географическая дифференциация Palimbia. В северной части ареала распространены растения, в соцветии которых имеются только ветви первого и второго порядка (синфлоресценцию в целом можно охарактеризовать как щитковидную), а соцветия с ветвями третьего порядка (в ключах для определения из называют раскидистыми метелками) практически отсутствуют. Западнее Волги почти не встречаются растения с ветвями только первого порядка. В Казахстане и Оренбургской области распространены все три варианта. Показаны корреляции на уровне 0.4–0.5 между признаками, которые предлагались в качестве диагностических.
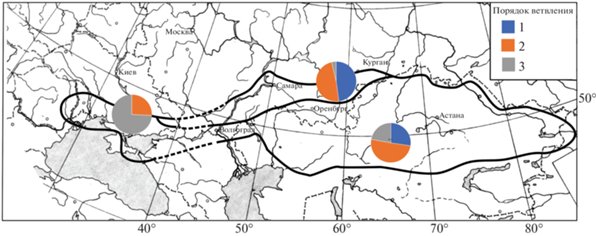
Palimbia Besser ex DC. – олиготипный или даже монотипный род Восточной Европы и Западной Азии; его виды (1–3) растут в зоне сухих засоленных степей и т.н. полупустынь от западного Китая до восточной Румынии. Для территории России и некоторых сопредельных стран долгое время указывался один вид рода Palimbia Besser ex DC. – P. rediviva (Pall.) Thell. (P. salsa (L.f.) Besser), распространенный в западном Китае (Синьцзян – Pimenov, 2017), восточной и северной части Казахстана (Korovin, 1963, Pimenov, 2020), Западной Сибири (юг Курганской обл.), Южном Урале, Поволжье, Северном Кавказе (Таманский полуостров) (Pimenov, Ostroumova, 2012), в Крыму (Yena, 2012), причерноморской части Украины (Kotov, 1955, Mosyakin, Fedoronchuk, 1999), Молдове (Negru, 2007), восточной Румынии (Яссы – Todor, 1958) в ковыльных и типчаковых степях, на сухих щебнистых склонах и на солонцах (рис. 1). В Ростовской области, Калмыкии и Ставропольском крае палимбия не была надена, в этой области дизъюнкции граница ареала показана прерывистой линией.
Рис. 1.
Карта ареала Palimbia rediviva s.l. и соотношение морфотипов с первым, вторым и третьим порядком ветвления в северной, западной и юго-восточной частях ареала.
Fig. 1. Map of Palimbia rediviva s.l. range and the ratio of morphotypes with the first, second, and third branching order in the northern, western, and southeastern parts of the range.
В 1931 году был описан новый вид P. turgaica Lipsky ex Woronov (Woronov, 1931). Лектотип – LE 01121243 из Башкортостана, Кучеровская № 281. В качестве признаков отличия от P. rediviva (Pall.) Thell. были указаны направленные вверх веточки, 6–20 лучей зонтика, более широкие листочки обертки, тогда как у P. rediviva (P. salsa) оттопыренные веточки и 5–7 лучей зонтика. На рисунке 535 (стр. 819) P. turgaica изображена с направленными вверх ветвями, терминальный зонтик с 4 лучами, зонтики на ветвях первого порядка с 4–8 лучами, что не вполне соответствует диагнозу (6–20 лучей). Воронов отметил, что “некоторые образцы затруднительно с уверенностью отнести к тому или иному виду” (стр. 818).
Другие ботаники предлагали для разграничения двух видов некоторые дополнительные признаки: характер общего соцветия – щитковидная или раскидистая метелка (Kotov, 1955; Korovin, 1963), порядок ветвления (Korovin, 1963), длина плодоножек, число междоузлий от розетки до первой ветви, ширина сегментов листа (Ryabinina, Knyazev, 2009)
Среди авторов нет единой точки зрения об ареалах видов. Рябинина, Князев (Ryabinina, Knyazev, 2009) считали, что P. rediviva распространена только к западу от Волги. Коровин (Korovin, 1963), Пименов (Pimenov, 1983, 2020), Пименов и Остроумова (Pimenov, Ostroumova, 2012) полагали, что P. rediviva и P. turgaica встречаются как в западной, так и в восточной частях ареала Palimbia.
Б.К. Шишкин (Shishkin, 1951) и его последователи (Vinogradova, 2004; Yelenevskii et al., 2009) не соглашались с самостоятельностью P. turgaica и включали это название в синонимику P. rediviva (P. salsa).
Учитывая сохраняющиеся в литературе различия в таксономический трактовке P. redivivа и P. turgaica, мы решили переисследовать этот вопрос, с использованием, впервые для этого таксона, методов молекулярной таксономии (Degtyareva et al., 2021), и одномерной и многомерной статистики (данное сообщение).
В первой работе отношения трех видов (P. rediviva, P. turgaica и P. defoliata (Ledeb.) Korovin) изучались методами ядерной (ITS яд-р ДНК) и хлоропластной (psbA-trnH хр ДНК) филогеномики. Все три вида по молекулярным данным относятся к одной кладе, причем образцы, определенные как P. turgaica (4 образца из Казахстана), нашли свое место среди более многочисленных образцов P. rediviva, т.е. с молекулярной точки зрения виды не отличаются. Одновременно была показана довольно значительная молекулярная вариабельность P. rediviva.
Задача настоящей статьи – использовать одномерный и многомерный анализ морфологических диагностических признаков для оценки возможности различения двух видов по этим признакам; оценить статус P. turgaica.
МАТЕРИАЛ И МЕТОДЫ
Были измерены все образцы Palimbia из гербариев LE, MHA и MW (Seregin, 2022), типовые образцы P. rediviva (B-W05860-01, B-W05860-02, B-W05860-03, LINN-HL356-11, M0173089 (Global plants…, 2000–2022) и P. turgaica (LE 01121242-01121249), всего 934 особи. Растения имели разную степень сохранности, были собраны в разные фенофазы, так что далеко не на каждой особи удавалось выявить все признаки.
Порядок ветвления побегов определяли с учетом мелких ветвей, зонтики на которых иногда засыхают, не приступая к цветению. Подсчитывали число лучей терминального зонтика, число междоузлий на стебле от розетки листьев до первой ветви и в области соцветия до терминального зонтика. Измеряли длину 5 цветоножек в терминальном зонтике, определяли среднее значение. Было замечено, что после цветения цветоножки удлиняются, пока плоды не достигнут длины около 2 мм. Для сравнения длин цветоножек взяты особи со зрелыми плодами и молодыми плодами длиной 2 и более миллиметров. На ранних этапах развития соцветия боковые ветви плотно прижаты к главному стеблю; ветви занимают свое окончательное положение и приобретают жесткость ко времени распускания цветков на ветвях первого порядка. Углы отхождения ветвей первого порядка измеряли на полностью развитых растениях без явных механических повреждений, по 1–3 ветви на каждом растении, вычисляли среднее для особи. Измеряли длину стебля от розетки листьев до первой ветви и от первой ветви до терминального зонтика, длину ветвей. В области общего соцветия стебель извилистый, мы определяли его длину как сумму междоузлий. Прикорневые листья у палимбий трижды перисторассеченные на линейные или нитевидные сегменты, ко времени цветения засыхают, а в период плодоношения растения обычно вообще не имеют пластинок прикорневых листьев; стеблевые листья редуцированы до чешуевидных влагалищ. Ширину конечных сегментов листьев розетки удалось измерить у вегетативных особей и у немногих генеративных. Листочки оберточки ко времени цветения начинают засыхать и теряют форму, впоследствии часто опадают; ширину листочков удалось измерить только у 22 особей, материала для оценки этого признака недостаточно.
Число лучей терминального зонтика, порядок ветвления, число междоузлий в гербариях LE и MHA подсчитывали непосредственно на гербарных листах, а из MW использовали сканированные изображения на сайте гербария (Seregin, 2022).
Для измерений линейных размеров и углов использовали фотографии гербарных образцов и программу ImageScope S в режиме ручных измерений с помощью “мыши”.
Эти данные использованы для построения диаграмм распределения значений признаков и для вычисления корреляций между признаками с помощью программы Microsoft Excel 2016.
Число лучей терминального зонтика определено для 695 особей, углы между ветвями и главным стеблем на 778 особях, длину плодоножек на 354, число междоузлий между розеткой и первой ветвью на 587, длину этого участка стебля на 665, ширину сегментов листа на 131 особи, порядок ветвления на 834, число междоузлий и длину главного стебля в соцветии на 649, длину ветвей на 499 особях.
Для изучения изменчивости объектов по многим признакам широко используются (отнюдь не только в биологии) многомерные методы ординации, такие как метод главных компонент. Цель этих методов – визуализация взаимоотношений объектов (таксонов) в пространстве меньшей размерности с сохранением максимума информации об их сходстве и различии. На три первые главные компоненты приходится большая часть изменчивости. Проекции объектов на плоскости, образуемые этими компонентами, позволяют убедительно судить об отношениях объектов (в данном случае о близости двух видов) по многим признакам.
Анализ главных компонент основан на материалах 186 особей, у которых удалось измерить все 6 признаков: общую длину стебля, число лучей терминального зонтика, число междоузлий главного стебля от розетки до первой ветви и в области инфлоресценции, углы между ветвями и главным стеблем, длину плодоножек11. Ширину сегментов листа не использовали, так как в выборке почти не было плодоносящих растений с сохранившимися листьями. Обработка данных проведена с помощью пакета программ PaSt (Hammer et al., 2001) по нормализованным значениям переменных. Точки разбиты по регионам: 1 – северный (Курганская и Челябинская области, республика Башкортостан, Саратовская, Самарская и Ульяновская области); 2 – Крым и Таманский полуостров; 3 – Волгоградская область; 4 – Казахстан и Оренбургская область. Анализ крымско-таманских и волгоградских растений проведен отдельно, потому что в Воронежской и Ростовской областях, Калмыкии и Ставропольском крае палимбия до сих пор не найдена, наблюдается значительная дизъюнкция ареала.
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
Морфометрические данные изученных типовых образцов представлены в табл. 1.
Таблица 1.
Морфометрические признаки типовых образцов Palimbia rediviva и P. turgaica Table 1. Morphometric characters of type specimens of Palimbia rediviva and P. turgaica
| Образец/Specimen | Тип/Type | Признаки/Characters | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 01 | 02 | 03 | 04 | 05 | 06 | 07 | 08 | 09 | 10 | 11 | ||
| Palimbia rediviva | ||||||||||||
| M0173089b | isolecto | 34.9 | 3 | 3 | 65 | 3.1 | 4 | 9 | 14.6 | 20.3 | 7.3 | |
| LINN-HL356-11a | isolecto | 2 | 4 | 68 | 4.1 | 7 | 19.4 | 5.4 | ||||
| LINN-HL356-11b | 0.37 | |||||||||||
| B-W05860-01b | isolecto | 32.8 | 2 | 3 | 75 | 3.7 | 7 | 7 | 18.7 | 14.1 | 5.4 | |
| B-W05860-02a | lecto | 3 | 7.0 | |||||||||
| B-W05860-02b | 0.22 | |||||||||||
| B-W05860-03a | isolecto | 3 | 57 | 2.1 | 8.7 | |||||||
| B-W05860-03b | 50.4 | 3 | 6 | 77 | 3.9 | 14 | 8 | 33.5 | 16.9 | 5.1 | ||
| B-W05860-03c | 0.37 | |||||||||||
| Palimbia turgaica | ||||||||||||
| LE 01121242a | isolecto | 51.2 | 2 | 8 | 43 | 3.0 | 1 | 6 | 7.3 | 44.0 | 15.5 | 0.51 |
| LE 01121242b | isolecto | 43.0 | 1 | 4 | 3 | 31.3 | 11.7 | 0.35 | ||||
| LE 01121243 | lecto | 55.6 | 2 | 11 | 4.2 | 5 | 30.7 | 24.8 | 7.8 | 0.46 | ||
| LE 01121244a | isolecto | 2 | 36 | 2 | 11.2 | 0.43 | ||||||
| LE 01121244b | isolecto | 64.4 | 2 | 9 | 44 | 3.9 | 4 | 7 | 30.7 | 33.6 | 8.4 | |
| LE 01121245a | syn | 28.3 | 2 | 6 | 40 | 3.1 | 2 | 4 | 11.9 | 16.3 | 3.9 | |
| LE 01121245b | syn | 26.3 | 2 | 6 | 45 | 4.2 | 3 | 5 | 12.2 | 14.1 | 4.9 | |
| LE 01121246 | syn | 37.0 | 0.48 | |||||||||
| LE 01121247a | syn | 3 | 5.5 | 2 | 8.6 | |||||||
| LE 01121247b | syn | 30.7 | 2 | 5 | 64 | 3.8 | 5 | 15.8 | 14.9 | |||
| LE 01121248a | syn | 2 | 3 | 76 | 3.0 | 5 | 10.0 | 3.7 | ||||
| LE 01121248b | syn | 2 | 3 | 84 | 2.0 | 5 | 11.3 | 2.2 | ||||
| LE 01121248c | syn | 2 | 3 | 51 | 2.1 | 5 | 6.6 | 4.4 | ||||
| LE 01121248d | syn | 2 | 4 | 71 | 2.2 | 6 | 12.5 | 3.5 | ||||
| LE 01121248e | syn | 2 | 3 | 80 | 1.8 | 4 | 12.4 | 1.7 | ||||
| LE 01121249a | syn | 30.4 | 2 | 4 | 38 | 2.8 | 6 | 8 | 11.4 | 19.0 | 7.0 | |
| LE 01121249b | syn | 22.0 | 3 | 59 | 3.7 | 4 | 10.1 | 11.9 | 6.7 | |||
| LE 01121249c | syn | 25.6 | 3 | 38 | 3.2 | 4 | 9.4 | 16.2 | ||||
Примечание. Признаки: 01 – общая длина главного стебля, см; 02 – порядок ветвления; 03 – число лучей терминального зонтика; 04 – углы между главным стеблем и ветвями, градусы; 05 – плодоножки, мм; 06 – стебель ниже синфлоресценции, число междоузлий; 07 – синфлоресценция, число междоузлий; 08 – стебель ниже синфлоресценции, длина, см; 09 – синфлоресценция, длина главного стебля, см; 10 – крупные ветви, длина, см; 11 – ширина сегментов листа, мм.
Note. Characters: 01 – main stem, total length, cm; 02 – branching order; 03 – terminal umbel, ray number; 04 – angles between main stem and branches, degrees; 05 – pedicel length, mm; 06 – stem below synflorescence, number of internodes; 07 – synflorescence, number of internodes; 08 – stem below synflorescence, length, cm; 09 – synflorescence, main stem length, cm; 10 – larger branches length, cm; 11 – width of leaf segments, mm.
Типовой материал Palimbia rediviva. В работе П.С. Палласа (Pallas, 1783), где описан Peucedanum redivivum, на таблице VIII изображено растение с ветвями 1, 2 и 3-го порядка, терминальный зонтик с 2 лучами. Образцы этого вида, собранные Палласом, имеются в гербариях B-Willd, BM, K, LINN, M, в качестве лектотипа был выбран образец B-W05860 02 (Pimenov, 2020: 202). На сайте plants.JSTOR.org (Global plants … 2000–2022) имеются высококачественные изображения 5 гербарных листов: M 0173089 (2 особи – справа P. rediviva, слева растение с клубнем – Elaeosticta lutea); LINN-HL 356-11 (2 особи P. rediviva – вегетативная и фрагмент генеративной); B-W 05860-01 (2 особи – справа P. rediviva, а слева, видимо, фрагмент E. lutea), B-W 0586 0-02 (2 особи P. rediviva – прикорневой лист и фрагмент крупного генеративного растения без терминального зонтика); B-W 05860-03 (3 особи – фрагменты двух генеративных растений и одна вегетативная). Растения ветвятся до второго или третьего порядка, терминальные зонтики с 3–6 лучами, зонтики первого порядка с 3–8 лучами. Число междоузлий от основания стебля до первой ветви 4–14, длина этой части стебля 14–37 см. Число междоузлий от первой ветви до терминального зонтика 5–9, длина этой части стебля 11–20 см. Средние для особи углы отхождения ветвей от 45 до 77°. Ширина сегментов листа от 0.22 до 0.37 мм.
Типовой материал P. turgaica. Все сборы, процитированные во “Флоре Юго-Востока” (Wo-ronow, 1931), хранятся в гербарии Ботанического ин-та РАН (LE). В качестве лектотипа В.М. Виноградова (Vinogradova, 2003) выделила лист LE 01121243: одно растение с розеткой листьев, терминальным зонтиком с 11 лучами и зонтиками на ветвях 1-го порядка с 8–9 лучами, что соответствует диагнозу вида. Угол отхождения ветвей определить невозможно, так как растения находятся в самом начале цветения, общее соцветие еще не приобрело окончательной структуры. Изолектотипы LE 01121242 и LE 01121244 относятся к тому же сбору (Башкортостан, Кучеровская 283) и имеют 6–9 лучей зонтиков. LE 01121245 из Саратовской области соответствует диагнозу: терминальные зонтики с 6 лучами, углы отхождения ветвей 40–45°. LE 01121246 из Саратовской области – крупное растение с еще не сформировавшимся соцветием. Растения из Оренбургской области, процитированные во “Флоре Юго-Востока”, не соответствуют диагнозу P. turgaica. LE 01121247 – одно растение с корнем, поврежденным главным стеблем и двумя крупными боковыми ветвями, фрагмент другого растения (возможно, главный стебель с терминальным зонтиком и ветвями) расположен между этими ветвями. Число лучей зонтиков 4–7, углы 64°. На листе LE 01121248 расположены 5 небольших растений без корней, у которых зонтики с 3–5 лучами, углы отхождения ветвей 71–84°, что никак не ассоциируется с “направленными вверх веточками”, только у одной особи (средней) углы 51°, но зонтики с 3 лучами. LE 01121249 – три растения с корнями, веточки направлены вверх (37–58°), зонтики с 3–4, у правой особи с 3–6 лучами. Таким образом, даже автор вида P. turgaica процитировал ряд экземпляров, не соответствующих диагнозу этого вида.
Другие образцы из гербариев LE, MHA и MW. В попытках различить P. turgaica и P. rediviva большое значение придавали облику синфлоресценции (щитковидное – раскидистая метелка); объективным показателем облика соцветия можно считать порядок ветвления, по этому признаку мы выделили три группы (морфотипа) A, B и C (рис. 2).
Рис. 2.
Схемы расположения побегов и сложных зонтиков у морфотипов Palimbia rediviva s.l.: A – синфлоресценция с ветвями только первого порядка; B – с ветвями первого и второго порядка; C – с ветвями первого, второго и третьего порядка.
Fig. 2. Arrangements of shoots and compound umbels in the morphotypes of Palimbia rediviva s. l.: A – synflorescence with branches of the first order only; B – with branches of the first and second order; C – with branches of the first, second and third order.
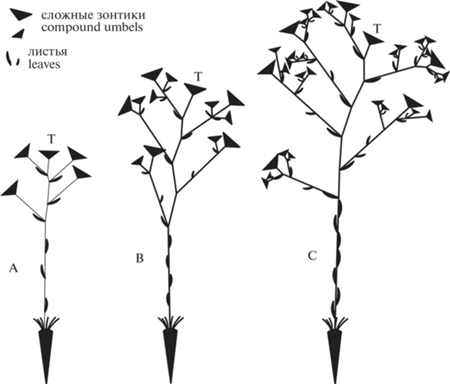
Морфотип А – растения с ветвями только первого порядка, 203 особи.
Морфотип B – растения с ветвями первого и второго порядка, 349 особей.
Морфотип C – растения с ветвями первого, второго и третьего (редко четвертого) порядка, 217 особей.
Цветки терминального зонтика всегда распускаются первыми. Верхние 1–2 ветви в большинстве случаев перевершинивают терминальный зонтик, они длиннее расстояния от соответствующего узла до терминального зонтика, независимо от порядка ветвления. Самые длинные ветви первого порядка обычно располагаются в нижней трети синфлоресценции, независимо от порядка ветвления. У достаточно мощных растений морфотипа C веточки второго порядка также перерастают концы материнских ветвей. В общем соцветии у морфотипов А и В наиболее заметны терминальный зонтик и боковые зонтики первого порядка. Крупные экземпляры морфотипа С имеют сильно разветвленное соцветие, терминальный зонтик нередко мельче боковых зонтиков первого порядка, находится в глубине общего соцветия и не доминирует в растении. Облик соцветия зависит от фенологической фазы.
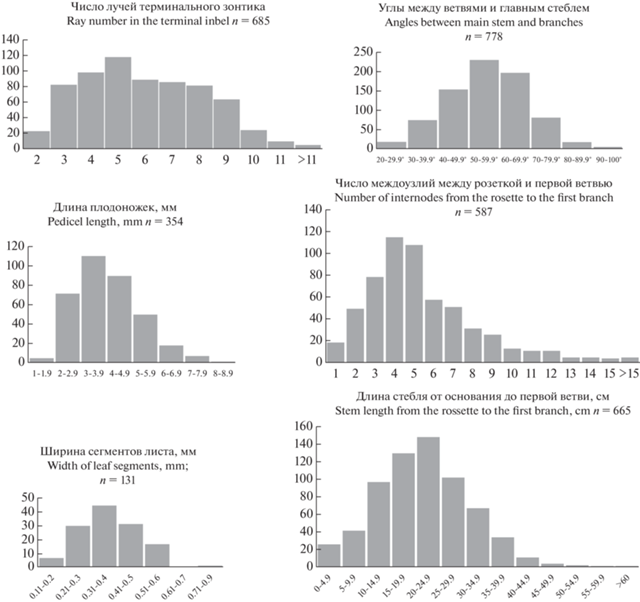
На рис. 3 представлены распределения значений признаков, предлагавшихся для диагностики видов: число лучей зонтика, угол отхождения боковых ветвей, длина плодоножек, число междоузлий между розеткой и первой ветвью и длина этого участка, ширина сегментов листа. Все распределения одновершинные, хотя не все могут быть названы нормальными (гауссовскими). Уже эта одновершинность распределений позволяет усомниться в наличии в выборке особей разных таксонов, особенно видов. Ни один признак не имеет хиатусов и не годится для диагностики.
Рис. 3.
Распределения значений количественных признаков Palimbia rediviva s.l., предлагавшихся для выделения P. turgaica. По оси Y – число особей.
Fig. 3. Distributions of quantitative trait values of Palimbia rediviva s.l. proposed for the identification of P. turgaica. Y-axis – number of individuals.
В то же время наблюдается некоторая географическая дифференциация (рис. 1). В северной части ареала (Курганская и Челябинская области, республика Башкортостан, Саратовская, Самарская и Ульяновская области и, вероятно, донецкая лесостепь, где растения были определены как P. turgaica (Kotov, 1955), встречаются преимущественно растения с ветвями первого и второго порядка (морфотипы A и B), а сильно разветвленные растения практически отсутствуют. Западнее Волги очень мало растений с ветвями только первого порядка, встречаются преимущественно морфотипы B и C, а в Казахстане и Оренбургской области встречаются все три морфотипа. Самый многочисленный морфотип В встречается по всему ареалу. Растения с очень большим числом междоузлий на стебле между розеткой листьев и первой ветвью (15–18) встречаются в Крыму, наряду с 8–14 междоузлиями; самые узкие сегменты листьев (0.15–0.29 мм) отмечены в Волгоградской области, наряду с 0.30–0.37 мм; самые широкие сегменты отмечены у одного растения в Саратовской области (0.66 мм) и одного в Самарской (0.88 мм), у других растений из северной части ареала ширина сегментов 0.34–0.53 мм.
Были определены корреляции между количественными признаками (табл. 2). Значения более 0.5 показаны для показателей общей мощности растений (общая длина стебля, длина стебля ниже первой ветви и в соцветии, длина ветвей, число междоузлий), а также между признаками синфлоресценции (порядок ветвления, число междоузлий и длина главного стебля в области соцветия, длина ветвей).
Таблица 2.
Корреляции между признаками Palimbia rediviva s. l. Жирным шрифтом выделены значения >0.5, серой заливкой от 0.4 до 0.5 Table 2. Correlations between characters of Palimbia rediviva s. l. Values >0.5 are highlighted in bold, values from 0.4 to 0.5 are filled gray
| 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | ||
|---|---|---|---|---|---|---|---|---|---|---|
| 1 | Общая длина главного стебля Length of main stem |
0.39945 | –0.00959 | 0.13598 | 0.47759 | 0.40629 | 0.48453 | 0.73557 | 0.70605 | 0.60978 |
| 2 | Порядок ветвления Branching order |
–0.4759 | 0.30321 | 0.20024 | 0.49303 | 0.66309 | 0.092148 | 0.50621 | 0.48281 | |
| 3 | Число лучей терминального зонтика Ray number of the terminal umbel |
–0.35167 | –0.05195 | –0.47306 | –0.41057 | 0.083556 | –0.07788 | –0.01901 | ||
| 4 | Углы отхождения ветвей Angles between main stem and branches |
0.13084 | 0.49073 | 0.2896 | 0.14704 | 0.021622 | –0.07782 | |||
| 5 | Длина плодоножек Pedicel length |
0.18032 | 0.3268 | –0.00737 | 0.45833 | 0.2549 | ||||
| 6 | Число междоузлий от розетки до первой ветви Number of internodes from rosette to the first branch |
0.35813 | 0.57156 | 0.069452 | 0.040561 | |||||
| 7 | Число междоузлий в соцветии Number of internodes in inflorescence |
–0.01232 | 0.72284 | 0.50029 | ||||||
| 8 | Длина стебля от розетки до первой ветви Length of stem from rosette to the first branch |
0.039624 | 0.0971 | |||||||
| 9 | Длина главного стебля в синфлоресценции Length of the main stem in synflorescence |
0.80371 | ||||||||
| 10 | Длина самой крупной ветви Length of the largest branch |
Порядок ветвления, число лучей терминального зонтика, углы отхождения ветвей, число междоузлий от розетки до первой ветви скоррелированы слабее, значения от 0.4 до 0.5 выделены в таблице серой заливкой. Связь этих признаков уже была замечена составителями ключей для определения растений (Woronow, 1931; Kotov, 1955; Korovin, 1963; Pimenov, 1983; Ryabinina, Knyazev, 2009 и др.), но на деле корреляция не очень высока.
Для порядка ветвления и числа лучей терминального зонтика показана отрицательная корреляция -0.4759. Тенденция к редукции терминального зонтика известна и у других зонтичных. У некоторых видов с многократно разветвленными соцветиями терминальный зонтик отсутствует, например, Dorema aitchisonii, Seseli tortuosum, Ferula tatarica, F. tuberifera. Известны случаи, когда терминальный зонтик содержит меньше лучей, чем боковые. У Anthriscus sylvestris и Falcaria vulgaris встречаются варианты как со слабо развитым терминальным зонтиком, так и совсем без него (Reuther, Classen-Bockhoff, 2010; Kusnetzova, Timonin, 2017).
Число междоузлий стебля от розетки до первой ветви скоррелировано не только с длиной этого участка, но и с порядком ветвления, числом лучей зонтика (отрицательно) и с углом отхождения боковых ветвей. Интересно, что порядок ветвления практически не коррелирует (0.092148) с длиной стебля до первой ветви, но имеет корреляцию 0.49303 с числом междоузлий этого участка. Длина плодоножек связана только с длиной стебля и длиной соцветия, а с известными диагностическими признаками скоррелирована слабо (0.007–0.326).
При анализе главных компонент (рис. 4, табл. 3 и 4) первая компонента включает 46.98% дисперсии, вторая – 20.77%, в сумме 67.7% (табл. 3); таким образом, проекция точек на плоскость первой и второй компонент довольно точно отражает положение точек в многомерном пространстве. Коэффициенты проекций точек на первую компоненту для разных признаков от 0.36 до 0.48 по абсолютной величине, ни одна из переменных не преобладает (табл. 4). Точки на диаграмме образуют сплошное облако, изученные особи не разделяются на изолированные группы, хотя области типовых образцов в многомерном пространстве не перекрываются. Области точек Волгоградской области и крымско-таманских в значительной степени совпадают, несмотря на дизъюнкцию. Области западного региона (Крым, Тамань и Волгоградская обл.) не перекрываются с растениями из северной части ареала. Точки растений из Казахстана и Оренбургский области показывают наибольший разброс и перекрываются со всеми остальными регионами (рис. 4).
Рис. 4.
Результаты анализа главных компонент по 6 морфологическим признакам. Регионы: 1 ⚪ – северная часть ареала – Курганская и Челябинская области, республика Башкортостан, Саратовская, Самарская и Ульяновская области; 2 + – Крым и Таманский полуостров; 3 ◇ – Волгоградская обл.; 4 ◻ – Казахстан и Оренбургская обл.
Fig. 4. Results of principal component analysis for 7 morphological characters. Regions: 1 ⚪ – northern part of the range: Kurgan and Chelyabinsk regions, Republic of Bashkortostan, Saratov, Samara and Ulyanovsk regions; 2 + – Crimea and Taman peninsulas; 3 ◇ – Volgograd Region; 4 ◻ – Kazakhstan and Orenburg Region.
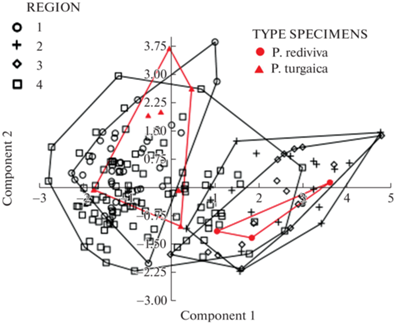
Таблица 3.
Характеристики главных компонент Table 3. Summary of the principal components (PC)
| Главные компоненты PC | Собственные значения Eigenvalue |
Доля объясненной дисперсии, % Proportion of the explained variance, % |
|---|---|---|
| 1 | 2.81861 | 46.977 |
| 2 | 1.24602 | 20.767 |
| 3 | 0.67055 | 11.176 |
| 4 | 0.601419 | 10.024 |
| 5 | 0.413789 | 6.8965 |
| 6 | 0.249617 | 4.1603 |
Таблица 4.
Нагрузки на главные компоненты (коэффициенты) Table 4. Loadings of variables into principal components (coefficients)
| Признаки/Characters | PC 1 | PC 2 | PC 3 | PC 4 | PC 5 | PC 6 |
|---|---|---|---|---|---|---|
| Общая длина главного стебля Length of main stem |
0.40817 | 0.52731 | 0.070169 | 0.31366 | –0.22948 | –0.63196 |
| Число лучей терминального зонтика Ray number of the terminal umbel |
–0.35877 | 0.50251 | 0.45461 | 0.46593 | 0.1697 | 0.40768 |
| Число междоузлий от розетки до первой ветви Number of internodes from rosette to first branch |
0.48453 | –0.17751 | 0.34049 | 0.05889 | –0.63327 | 0.46183 |
| Число междоузлий в соцветии Number of internodes in inflorescence |
0.43958 | 0.15112 | –0.64696 | 0.34719 | 0.27728 | 0.40981 |
| Углы отхождения ветвей Angles between main stem and branches |
0.37408 | –0.47322 | 0.44501 | 0.29888 | 0.56798 | –0.16174 |
| Длина плодоножек Pedicel length |
0.36978 | 0.4372 | 0.23639 | –0.68643 | 0.34355 | 0.16442 |
ЗАКЛЮЧЕНИЕ
Вид Palimbia turgaica невозможно надежно отличить по морфологическим признакам от Palimbia rediviva. Первый был описан как крайний морфологический вариант из северной части ареала, но он связан непрерывными переходами с остальными особями. Среди процитированных автором образцов (Woronow, 1931) далеко не все соответствуют предложенному диагнозу. Все авторы ключей предлагали в качестве важного диагностического признака угол отхождения ветвей от главного стелбя, но этот признак зависит от фенологической фазы, на ранних стадиях развития соцветия углы всегда острые. Palimbia rediviva s.l. – это полиморфная группа с широким ареалом, которая нуждается в дальнейшем изучении.
Palimbia rediviva (Pall.) Thell., 1926, in Hegi, Ill. Fl. Mittel-Eur. 5, 2: 1364.
≡ Peucedanum redivivum Pall., 1779, in Acta Acad. Sci. Imp. Petrop. 2: 252, tab. VIII.
≡ Sison salsum L.f., 1781, in Suppl. Pl.: 181.
Тип: Россия “In salsis limosis aridissimis circa Wolgam [In aridis deserti caspiae]. P.S. Pallas” Лектотип: B-W05860-02!, выделен М.Г. Пименовым (Pimenov, 2020: 202); изолектотипы: B-W05860-01!, B-W05860-03!, LINN-HL356-11!, M0173089!.
= Palimbia turgaica Lipsky ex Woronow, 1931, Trudy Bot. Sada Akad. Nauk SSSR 43, 2 (Fl. Ross. Austro-Or. 5): 818, fig. 535 .
Тип: Россия “Bashkiria, kanton Sterlitamak, Chekchenevskoe, pag. Jakschimbetova. 03.07.1928. S.E. Kucherovskaya 281” LE 01121243! – лектотип, выделен В.М. Виноградовой (Vinogradova, 2003: 109); изолектотипы: LE 01121242!, LE 01121244!; синтипы LE 01121245!, LE 01121246!, LE 01121247!, LE 01121248!, LE 01121249!
Список литературы
[Degtjareva et al.] Дегтярева Г.В., Остроумова Т.А., Самигуллин Т.Х., Пименов М.Г. 2021. Положение рода Palimbia в системе семейства Umbelliferae. – Проблемы ботаники Южной Сибири и Монголии 20, 1: 156–159. https://doi.org/10.14258/pbssm.2021029
Global Plants on JSTOR. 2000–2022. Available from: https://plants.jstor.org/ (last accessed: 30.06.2022).
Hammer Ø., Harper D.A.T., Ryan P.D. 2001. PaSt: Paleontological statistics software package for education and data analysis. – Paleontologia Electronica. 4 (1): 9. Free software available at https://www.nhm.uio.no/english/research/resources/past/index.html
[Korovin] Коровин Е.П. 1963. Зонтичные – Umbelliferae Moris. – В кн.: Флора Казахстана. Т. 6. Алма-Ата. С. 258–428.
Kotov M.I. 1966. Umbelliferae Moris. – В кн.: Флора УРСР. Т. VII. Київ. С. 460–618.
[Kusnetzova, Timonin] Кузнецова Т.А., Тимонин А.К. 2017. Соцветие: морфология, эволюция, таксономическое значение (применение комплементарных подходов). М. 183 с.
Mosyakin S.L., Fedoronchuk M.M. 1999. Vascular plants of Ukraine. A nomenclatural checklist. Kiev. 345 p.
Negru A. 2007. Determinatór de plante din flora Republicii Moldova. Chişinău. 391 p.
Pallas P.S. 1779. Descriptiones plantarum sibiriae peculiarium. – Acta Acad. Sci. Imp. Petrop. 1779 (2): 247–272. Available from: https://www.biodiversitylibrary.org/page/10127809#page/285/mode/1up
[Pimenov] Пименов М.Г. 1983. Umbelliferae – Зонтичные. – В кн.: Определитель растений Средней Азии. Критический конспект флоры. Т. 7. Ташкент. С. 167–322.
[Pimenov, Ostroumova] Пименов М.Г., Остроумова Т.А. 2012. Зонтичные (Umbelliferae) России. М. 477 с.
Pimenov M.G. 2017. Updated checklist of Chinese Umbelliferae: nomenclature, synonymy, typification, distribution. – Turczaninowia 20 (2): 106–239. https://doi.org/10.14258/turczaninowia.20.2.9
Pimenov M.G. 2020 Updated checklist of the Umbelliferae of Middle Asia and Kazakhstan: nomenclature, synonymy, typification, distribution. – Turczaninowia. 23 (4): 127–257. https://doi.org/10.14258/turczaninowia.23.4.12
Reuther K., Classen-Bockhoff R. 2009. Diversity behind uniformoty – inflorescence architecture and flowering sequence in Apiacee – Apioideae. – Plant Div. Evol. 128: 181–220.
[Ryabinina, Knyazev] Рябинина З.Н., Князев М.С. 2009. Определитель сосудистых растений Оренбургской области. М. 760 с.
[Schischkin] Шишкин Б.К. 1950. Umbelliferae. – В кн.: Флора СССР. Т. 16. М.; Л. С. 36–604.
Seregin A.P. (Ed.). 2022. Moscow Digital Herbarium: Electronic resource. – Moscow State University, Moscow. – Available at: https://plant.depo.msu.ru/ (accessed 21.06.2022).
Todor I. 1958. Umbelliferae. – In: Flora Republicii Populare Române. Vol. VI. București. P. 316–660.
[Vinogradova] Виноградова В.М. 2003. Типовые образцы таксонов семейства Apiaceae Восточной Европы, хранящиеся в гербарии Ботанического института им. В.Л. Комарова РАН (LE). Anthriscus – Trinia. – Бот. журн. 88 (8): 105–111.
[Vinogradova] Виноградова В.М. 2004. Apiaceae Lindl. (Umbelliferae Juss.) – Сельдереевые (Зонтичные). – В кн.: Флора Восточной Европы. Т. 11. М.–СПб. С. 331–437.
[Woronov] Воронов Ю.М. 1931. Umbelliferae (Moris.) Juss. – В кн.: Флора Юго-Востока европейской части СССР. Вып. 5. Л. С. 759–838.
[Yelenevskii et al.] Еленевский А.Г., Буланый Ю.И., Радыгина В.И. 2009. Определитель сосудистых растений Саратовской области. Саратов. 248 с.
[Yena] Ена А.Е. 2012. Природная флора Крымского полуострова. Симферополь. 231 с.
Дополнительные материалы
- скачать ESM.doc
- Приложение. Материал, использованый для анализа главных компонент.
Инструменты
Ботанический журнал