

Ботанический журнал, 2022, T. 107, № 2, стр. 180-197
МОРФОЛОГИЧЕСКАЯ ИЗМЕНЧИВОСТЬ IRIS PUMILA (IRIDACEAE) В НИЖНЕМ ПОВОЛЖЬЕ И НА ЮЖНОМ УРАЛЕ
А. С. Кашин 1, *, А. С. Пархоменко 1, Л. М. Абрамова 2, А. О. Кондратьева 1, А. В. Богослов 1, И. В. Шилова 1, А. В. Крюкова 2
1 Саратовский национальный исследовательский государственный университет имени Н.Г. Чернышевского
410012 г. Саратов, ул. Астраханская, 83, Россия
2 Южно-Уральский ботанический сад-институт УФИЦ РАН
450080 г. Уфа, ул. Менделеева, 195, корпус 3, Россия
* E-mail: kashinas2@yandex.ru
Поступила в редакцию 19.08.2021
После доработки 15.11.2021
Принята к публикации 16.11.2021
- EDN: LWBNXN
- DOI: 10.31857/S0006813622020065
Аннотация
Изучены 22 популяции ириса карликового (Iris pumila L.) из двух регионов – Южный Урал (Республика Башкортостан) и Нижнее Поволжье (Саратовская область). Установлено, что растения популяций I. pumila из двух географических локалитетов заметно отличаются друг от друга. Особи из популяций Южного Урала были крупнее по высоте генеративного побега, структурным элементам цветка и, в особенности, по длине и ширине листа. В свою очередь, растения популяций Нижнего Поволжья были крупнее по показателям диаметра цветка и диаметра клона. Ординация методами главных компонент и неметрического многомерного шкалирования также убедительно демонстрирует разделение популяционных выборок из двух географических локалитетов. При этом больший разброс точек, соответствующих совокупности средних значений морфологических признаков растений в популяциях Нижнего Поволжья, указывает на большую неоднородность условий произрастания вида в этой географической области по сравнению с Южным Уралом. На распределении изменчивости морфологических признаков по географическому градиенту в Нижнем Поволжье сказывается, прежде всего, большая амплитуда температур. Этому способствует и высота над уровнем моря мест произрастания популяций в данном регионе. На Южном Урале большее сходство морфологических параметров растений вида, независимо от пространственного положения популяций в данной географической области, обусловлено, прежде всего, меньшей амплитудой температур.
Изучение внутривидовой изменчивости редких растений имеет большое значение в популяционной биологии и экологии, поскольку позволяет оценить уровень фенотипической изменчивости, установить границы популяций, охарактеризовать микроэволюционные процессы и выявить экологические факторы, влияющие на формирование структуры популяций, что в конечном итоге способствует разработке мер по сохранению генофонда редких видов (Yoccoz, 2012; Kryukova, Abramova, 2018; Kryukova et al., 2018).
Гетерогенность окружающей среды в пространстве и времени обычно хорошо ощущается популяциями растений, которые по-разному приспосабливаются к этой неоднородности. Приспособление растений популяций конкретного вида к тем или иным условиям – как одного, так и разных в географическом отношении местообитаний – в естественных условиях реализуется посредством фенотипической пластичности. Фенотипической пластичностью называют способность одного и того же генотипа производить разные фенотипы в зависимости от условий среды. Диапазон фенотипов для данного признака, который может возникать в различных условиях среды, известен как “норма реакции”. Фенотипической пластичности в той или иной мере подвержены практически все признаки. В этом отношении фенотип является результатом взаимодействия между генами и окружающей средой, в которой происходило конкретное развитие организма. При этом, краткосрочные эволюционные реакции на неоднородность экологических факторов могут привести к генетической специализации, которая, в свою очередь, приводит к дифференциации популяций (Pemac, Tucić, 1998; Tucić et al., 2003; Tarasjev et al., 2009; Markov, Ivnitskiy, 2016; Avramov et al., 2017).
В соответствии с типами пластических реакций, проявляемых организмами, пластичность можно разделить на два основных вида: предвосхищающие модификации развития, опережающие те или иные изменения окружающей среды, и неизбежные изменения в результате непосредственного воздействия окружающей среды на фенотип. Предвосхищающая пластичность возникает в ответ на определенные экологические сигналы, характеризующие различные изменения в окружающей среде в ближайшем будущем. Считается, что пластичные фенотипические ответы на сигналы окружающей среды усиливают функции организма и, следовательно, приспособленность к среде, в которой этот организм обитает. В свою очередь, неизбежная (пассивная) пластичность характеризуется влиянием окружающей среды, вызванным неоптимальным уровнем ресурсов, на фенотип. В данном случае фенотипические изменения пропорциональны степени экологических воздействий. Таким образом, пластичность у растений может быть результатом прямой реакции признака на сигналы окружающей среды во время развития или пассивным результатом изменения скорости роста растений из-за различных внешних факторов (Tucić et al., 2003; Barišić Klisarić et al., 2012).
В настоящем исследовании предпринята попытка выявления морфологической дифференциации популяций Iris pumila L. на основании количественных признаков, измеряемых в период массового цветения растений. Исследование особенно актуально в связи с тем, что изучаемый вид относится к категории редких. I. pumila включен в Красную книгу РФ (Krasnaya…, 2008) с категорией и статусом редкости: 3 – редкий вид; а также Красные книги всех регионах РФ, где он произрастает, – в частности и на территориях исследованных регионов – Саратовской обл. (Krasnaya…, 2021) и Республики Башкортостан (Krasnaya…, 2011). Полученные результаты могут быть использованы для более объективной оценки состояния популяций вида на территории изученных регионов.
Ирис карликовый, I. pumila, представитель семейства Iridaceae, – травянистый короткокорневищный гемиэфемероидный поликарпик, 10–15 см высотой с желтыми или лиловыми цветками (при этом растения в популяциях часто сильно различаются по окраске цветка). В естественных местообитаниях вид образует клоны округлой формы разного размера в зависимости от возраста. Клоны I. pumila являются результатом развития горизонтально растущих и плотно упакованных сегментов (звеньев) корневища, которые распространяются радиально от центра каждого клона к периферии. Вид является ксерофитом и степным геофитом, произрастает в каменистых и луговых степях, на остепененных лугах; по щебнистым и каменистым склонам степных холмов западной, восточной и, в большей степени, южной экспозиции, на обнажениях. Предпочитает участки с разреженной растительностью. Имеет обширный ареал, охватывающий Центральную и Юго-восточную Европу. В России встречается в Астраханской, Белгородской, Волгоградской, Воронежской, Оренбургской, Ростовской, Самарской, Саратовской, Тамбовской, Ульяновской, Челябинской областях, в Краснодарском и Ставропольском краях, в Республиках Башкортостан, Чеченская, Кабардино-Балкарская, Карачаево-Черкесская, Северная Осетия–Алания, Ингушетия, Дагестан (Tucić et al., 2003; Alekseyeva, 2008; Yefimov et al., 2012; Yuritsyna, Vasyukov, 2014; Kryukova et al., 2014, 2018; Kryukova, Abra-mova, 2018; Abramova et al., 2019; Boltenkov, 2019).
В ряде исследований отмечается, что I. pumila является подходящим объектом для изучения фенотипической пластичности, а также пространственной и временной межпопуляционной дифференциации (Tarasjev et al., 2009, 2012; Barišić Klisarić et al., 2012; Avramov et al., 2017).
Данным исследованием охвачены два района произрастания I. pumila – Южный Урал (в границах Р. Башкортостан) и север Нижнего Поволжья (в границах Саратовской обл.). Первый район исследования находится на северо-восточной границе ареала и характеризуется менее засушливыми условиями произрастания вида, а второй – в центральной его части с более аридным климатом. Целью исследования было выявление особенностей морфологической изменчивости в центральной части ареала вида и на его границе в разных климатических условиях произрастания. Ожидалось, что на границе ареала в силу большей однородности условий произрастания полиморфизм должен быть у́же, а, учитывая более влажный и менее континентальный климат степного Предуралья, значения морфологических параметров выше, чем в центральной, более аридной части ареала.
МАТЕРИАЛЫ И МЕТОДЫ
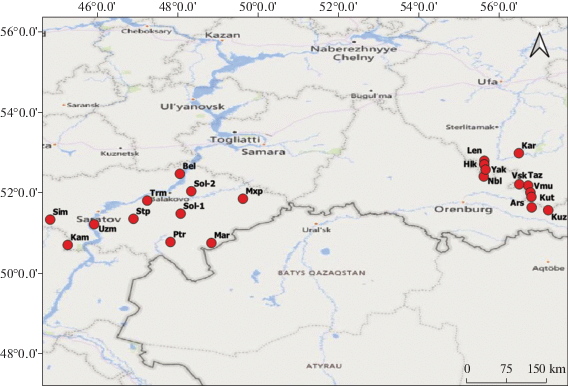
Исследовано 22 популяции I. pumila при одинаковом их числе для каждого из выбранных районов, т.е. 11 (275 растений) популяций исследовано в Нижнем Поволжье и 11 (1100 растений) – на Южном Урале (табл. 1, рис. 1). Так как ареал вида на севере Нижнего Поволжья охватывает фактически всю территорию Саратовской обл., для исследования отобрали популяции, более или менее равномерно распределенные по всей территории области. На Южном Урале ареал вида ограничен только юго-западными районами Республики Башкортостан (степное Предуралье). Соответственно исследованные популяции территориально произрастают гораздо ближе друг к другу, чем отобранные в Саратовской обл. На территории Предуралья исследование проводили в течение четырех лет, в то время как на территории севера Нижнего Поволжья – в течение одного года. Отчасти это было связано с тем, чтобы нивелировать разницу в размерах исследуемых территорий в сравниваемых частях ареала. Исходили также из того, что если проверяемая гипотеза верна, то даже при многолетнем мониторинге популяций с более однородными условиями произрастания диапазон изменчивости будет меньшим, чем при однолетнем изучении популяций с большей неоднородностью условий произрастания вида.
Таблица 1.
Местонахождение исследованных популяций Iris pumila Table 1. Location of the studied populations of Iris pumila
| Условное обозна-
чение популяции Conventional symbol for population |
Местонахождение популяции Population locality |
Координаты/Coordinates | |
|---|---|---|---|
| N | E | ||
| Республика Башкортостан/Republic of Bashkortostan | |||
| Taz | Зианчуринский район, окр. с. Тазларово Zianchurinsky district, vicinity of Tazlarovo village |
52.185807 | 56.699793 |
| Vmu | Зианчуринский район, окр. д. Верхний Муйнак Zianchurinsky district, vicinity of Verkhniy Muynak village |
52.017090 | 56.754400 |
| Kuz | Зианчуринский район, окр. д. Кужанак Zianchurinsky district, vicinity of Kuzhanak village |
51.574653 | 57.197240 |
| Ars | Зианчуринский район, окр. с. Арсенево Zianchurinsky district, vicinity of Arsenevo village |
51.643555 | 56.787757 |
| Kut | Зианчуринский район, ур. Куйтапкан Zianchurinsky district, Kuytapkan tract |
51.905430 | 56.780569 |
| Vsk | Кугарчинский район, гора Высокая Kugarchinsky district, Vysokaya Mt. | 52.217500 | 56.479074 |
| Len | Куюргазинский район, окр. д. Лена Kuyurgazinsky district, vicinity of Lena village |
52.801797 | 55.611802 |
| Nbl | Куюргазинский район, окр. с. Нижнее Бабаларово Kuyurgazinsky district, vicinity of Nizhneye Babalarovo village |
52.411747 | 55.598153 |
| Hlk | Куюргазинский район, окр. д. Холодный ключ Kuyurgazinsky district, vicinity of Kholodny Klyuch village |
52.712637 | 55.609113 |
| Yak | Куюргазинский район, окр. с. Якшимбетово Kuyurgazinsky district, vicinity of Yaksimbetovo village |
52.582541 | 55.643009 |
| Kar | Мелеузовский район, гора Караултау Meleuzovsky district, Karaultau Mt. |
52.989546 | 56.468915 |
| Саратовская область/Saratov Region | |||
| Ptr | Питерский район, граница с Новоузенским районом Pitersky district, border with Novouzensky district |
50.785110 | 47.824395 |
| Stp | Советский район, окр п. Степное Sovetsky district, vicinity of Stepnoye village |
51.36015 | 46.90591 |
| Bel | Хвалынский район, восточнее г. Хвалынска, гора Беленькая Khvalynsky district, east of Khvalynsk, Belenkaya Mt. |
52.47863 | 48.05778 |
| Mar | Дергачевский район, окр. х. Песчаный Мар Dergachevsky district, vicinity of Peschany Mar farm |
50.761518 | 48.840204 |
| Sol-1 | Ершовский район, окр. с. Новоряженка Ershovsky district, vicinity of Novoryazhenka village |
51.488845 | 48.078042 |
| Kam | Красноармейский район, окр. с. Каменка Krasnoarmeisky district, vicinity of Kamenka village |
50.71152 | 45.27483 |
| Sim | Лысогорский район, окр. с. Симоновка Lysogorsky district, vicinity of Simonovka village |
51.34137 | 44.83986 |
| Mxp | Пугачевский район, окр. с. Максютово Pugachevsky district, vicinity of Maksyutovo village |
51.85929 | 49.62207 |
| Sol-2 | Пугачевский район, окр. с. Солянка Pugachevsky district, vicinity of Solyanka village |
52.04402 | 48.34098 |
| Uzm | Энгельсский район, окр. с. Узморье Engels district, vicinity of Uzmorye village |
51.22534 | 45.93035 |
| Trm | Марксовский район, урочище Три Мара Marksovsky district, Tri Mara tract |
51.813144 | 47.248446 |
Морфологические характеристики с целью поддержания единообразия в их оценке учитывались только у растений генеративного состояния (Sharma, Pandit, 2011). В каждой популяции в один год наблюдения признаки измерялись у 25 особей. В качестве особи для морфометрических измерений считали клон. Всего было измерено 12 морфологических показателей (рис. 2): диаметр клона, число вегетативных побегов (лопаток), число листьев на вегетативный побег, длина второго листа, ширина второго листа, число генеративных побегов, длина генеративного побега, длина внешней доли околоцветника, ширина внешней доли околоцветника, длина внутренней доли околоцветника, ширина внутренней доли околоцветника, диаметр цветка.
Рис. 2.
Морфологические показатели Iris pumila, использованные в анализе: 1 – диаметр клона, 2 – число вегетативных побегов (лопаток), 3 – число листьев на вегетативный побег, 4 – длина второго листа вегетативного побега, 5 – ширина второго листа вегетативного побега, 6 – число генеративных побегов, 7 – длина генеративного побега, 8 – длина внешней доли околоцветника, 9 – ширина внешней доли околоцветника, 10 – длина внутренней доли околоцветника, 11 – ширина внутренней доли околоцветника, 12 – диаметр цветка.
Fig. 2. Morphological parameters of Iris pumila used in the analysis: 1 – clone diameter, 2 – number of vegetative shoots (blades), 3 – number of leaves per vegetative shoot, 4 – length of the second leaf on the vegetative shoot, 5 – width of the second leaf on the vegetative shoot, 6 – number of generative shoots, 7 – length of the generative shoot, 8 – length of the outer perianth lobe, 9 – width of the outer perianth lobe, 10 – length of the inner perianth lobe, 11 – width of the inner perianth lobe, 12 – flower diameter.

Анализ изменчивости морфологических показателей проводили с использованием описательной статистики (минимум и максимум показателя и его 95% доверительный интервал, коэффициент вариации), диаграмм размаха (среднее значение ± ошибка средней и стандартное отклонение) (Metody…, 2015), а также непараметрического анализа Манна–Уитни для сравнения средних двух независимых выборок (Khar′kova, Grzhibovskiy, 2014). На основе матрицы корреляций был проведен факторный анализ методом главных компонент (PCA) (Jolliffe, 2002; Zuur et al., 2009). Кроме того, были рассчитаны непараметрические коэффициенты корреляции Спирмена (Zar, 2010): в случае, если коэффициенты корреляции для коррелированных пар переменных превышали 0.90, один признак из пары исключался из факторного анализа (Nobis et al., 2016; Ocampo, d’Eeckenbrugge, 2017; Spaniel et al., 2017; Maia, Goldenberg, 2019).
Дополнительно выявлена и визуализирована взаимосвязь наблюдаемого паттерна морфологической изменчивости с некоторыми факторами окружающей среды (климатическими, географическими). Для этого был применен метод неметрического многомерного шкалирования (NMDS) с последующим наложением на плоскость ординации векторов, указывающих направление климатических или географических градиентов, а также силу их связи с наблюдаемым паттерном морфологической изменчивости.
Входные данные для NMDS представляли собой средние значения морфометрических показателей для каждой популяции из Нижнего Поволжья и Южного Урала, предварительно прошедшие процедуру стандартизации. Так как исследовались только количественные морфологические признаки, в качестве меры различия использовалось евклидово расстояние. Для каждого из 22 местоположений популяций ириса из открытой базы WorldClim ver. 2.0 (Fick, Hijmans, 2017) были взяты данные с пространственным разрешением 2.5 минуты (5 км), содержащие значения 19 биоклиматических характеристик и альтитуды (табл. 2). Кроме того, учитывались значения географической широты и долготы мест произрастания популяций. Полученные значения использовались в качестве переменных окружающей среды и были спроецированы на плоскость ординации в виде векторов, указывающих направление наиболее быстрого изменения значений соответствующей переменной (направление градиента), и имеющих длину, пропорциональную корреляции между переменной окружающей среды и осями ординации (сила градиента). Процедура расчета уровня значимости выявленных связей основывалась на применении метода случайных перестановок с числом итераций равным 999. Данный анализ был проведен в среде статистического программирования R ver. 4.1.0 (R Core Team, 2021).
Таблица 2.
Параметры векторов переменных окружающей среды, проецируемых на плоскость ординации Table 2. Parameters of vectors of environmental variables projected onto the ordination plane
| Переменная окружающей среды Environmental variable | Условное обозначе-ние переменной окружающей среды Environment variable symbol | Направляющие косинусы векторов Direction cosines of the vectors | Квадрат коэффициента корреляции (r2) Squared correlation coefficient (r2) | p-значение (Pr(>r)) p-value (Pr(>r)) |
|
|---|---|---|---|---|---|
| NMDS1 | NMDS2 | ||||
| Среднегодовая температура, ᵒС Average annual temperature, °С | bio1 | 0.99747 | 0.07115 | 0.6830 | 0.001*** |
| Среднемесячная суточная амплитуда температуры, °С Monthly average daily temperature range, °С |
bio2 | –0.99908 | –0.04295 | 0.6822 | 0.001*** |
| Изотермальность Isothermal | bio3 | –0.99981 | –0.01943 | 0.6713 | 0.001*** |
| Сезонность температуры (коэффициент вариации) Seasonality of temperature (coefficient of variation) |
bio4 | –0.99661 | –0.08230 | 0.5637 | 0.001*** |
| Максимальная температура наиболее теплого месяца, °С Maximum temperature of the warmest month, °С |
bio5 | 0.99896 | 0.04569 | 0.1817 | 0.140 |
| Минимальная температура наиболее холодного месяца, °С Minimum temperature of the coldest month, °С |
bio6 | 0.99875 | 0.05004 | 0.6955 | 0.001*** |
| Среднегодовая амплитуда колебания температуры, °С Annual average amplitude of temperature fluctuations, °С |
bio7 | –0.99871 | –0.05082 | 0.6256 | 0.001*** |
| Средняя температура наиболее влажного квартала, °С Average temperature of the wettest quarter, °С |
bio8 | –0.99730 | –0.07341 | 0.0784 | 0.454 |
| Средняя температура наиболее сухого квартала, °С Average temperature of the driest quarter, °С |
bio9 | 0.94424 | 0.32926 | 0.5596 | 0.001*** |
| Средняя температура наиболее теплого квартала, °С Average temperature of the warmest quarter, °С |
bio10 | 0.99870 | 0.05101 | 0.5756 | 0.002** |
| Средняя температура самого холодного квартала, °С Average temperature of the coldest quarter, °С |
bio11 | 0.99789 | 0.06487 | 0.6965 | 0.001*** |
| Среднегодовое количество осадков, мм Average annual rainfall, mm |
bio12 | 0.99884 | 0.04808 | 0.0952 | 0.401 |
| Количество осадков самого влажного месяца, мм Rainfall of the wettest month, mm |
bio13 | 0.99048 | –0.13765 | 0.0274 | 0.763 |
| Количество осадков самого сухого месяца, мм Rainfall of the driest month, mm |
bio14 | 0.99657 | –0.08279 | 0.1119 | 0.307 |
| Сезонность выпадения осадков (коэффициент вариации) Seasonality of precipitation (coefficient of variation) |
bio15 | –0.99725 | 0.07412 | 0.0947 | 0.362 |
| Количество осадков самого влажного квартала, мм Rainfall of the wettest quarter, mm |
bio16 | 0.99393 | –0.10998 | 0.0157 | 0.842 |
| Количество осадков самого сухого квартала, мм Rainfall of the driest quarter, mm |
bio17 | 0.99759 | –0.06935 | 0.1241 | 0.283 |
| Количество осадков самого теплого квартала, мм Rainfall of the warmest quarter, mm |
bio18 | 0.95800 | –0.28678 | 0.0035 | 0.956 |
| Количество осадков самого холодного квартала, мм Rainfall of the coldest quarter, mm |
bio19 | 0.99696 | 0.07795 | 0.2759 | 0.047* |
| Высота над уровнем моря, м Altitude above sea level, m |
Altitude | –0.98369 | –0.17988 | 0.5520 | 0.002** |
| Географическая широта Latitude | Latitude | 0.99998 | –0.00566 | 0.4921 | 0.002** |
| Географическая долгота Longitude | Longitude | 0.99234 | –0.12357 | 0.6731 | 0.001*** |
Морфометрические данные перед проведением факторного анализа, а также неметрического многомерного шкалирования подвергались процедуре стандартизации (Torrecilla et al., 2013; Biye et al., 2016; Finot et al., 2018; Cruz-Lustre et al., 2020).
Для расчетов и визуализации результатов использовались программы “Statistica 6.0” (STATISTICA, 2001) и “Past 3.26” (Hammer et al., 2001; Hammer, Harper, 2005), а также пакеты vegan (Oksanen et al., 2020) и ggpolot 2 (Wickham et al., 2016), реализованные в статистической среде R.
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
Сравнение выборок популяций Iris pumila из Нижнего Поволжья и Южного Урала методом описательной статистики показало, что у преобладающего большинства признаков средние значения, включая ошибку среднего, не перекрываются (табл. 3). По большинству морфологических признаков минимальные и максимальные значения в обеих группах также различались. Исключение составил показатель числа генеративных побегов, у которого перекрывались минимальные и средние значения. Средние значения большинства исследованных морфологических параметров были выше в выборке популяций с Южного Урала. Максимальное различие имело место между средними значениями длины и ширины второго листа: популяционная выборка с Южного Урала в два с лишним раза превосходила таковую из Нижнего Поволжья. У трех признаков – диаметр клона, число вегетативных побегов, и диаметр цветка – средние, а также минимальные и максимальные значения в популяциях Нижнего Поволжья были выше, чем в популяциях Южного Урала.
Таблица 3.
Диапазон изменчивости морфологических параметров I. pumila в Нижнем Поволжье и на Южном Урале Table 3. The range of variability of the morphological parameters of I. pumila in the Lower Volga region and in the South Urals
| Морфологический признак Morphological trait |
Диапазон изменчивости морфологического признака* The range of variability of morphological trait* |
Коэффициент вариации, % Coefficient of variation, % |
||
|---|---|---|---|---|
| Нижнее Поволжье Lower Volga region |
Южный Урал South Urals |
Нижнее Поволжье Lower Volga region |
Южный Урал South Urals |
|
| Диаметр клона, см Clone diameter, cm |
4.00–(27.41 ± 0.86)–81.00 | 2.00–(21.55 ± 0.36)–78.00 | 51.86 | 55.20 |
| Число вегетативных побегов, шт. Number of vegetative shoots |
1.00–(23.59 ± 1.61)–262.00 | 2.00–(18.78 ± 0.42)–102.00 | 112.98 | 73.60 |
| Число листьев на побег, шт. Number of leaves per shoot |
3.00–(4.69 ± 0.04)–7.00 | 3.00–(5.03 ± 0.02)–7.00 | 14.03 | 15.98 |
| Длина второго листа, см Length of the second leaf, cm |
2.60–(5.38 ± 0.08)–10.50 | 4.00–(14.25 ± 0.10)–26.00 | 26.12 | 23.22 |
| Ширина второго листа, см Width of the second leaf, cm |
0.30–(0.66 ± 0.01)–1.20 | 0.60–(1.36 ± 0.01)–2.60 | 29.25 | 21.62 |
| Число генеративных побегов, шт. Number of generative shoots |
1.00–(6.00 ± 0.42)–48.00 | 1.00–(5.43 ± 0.16)–74.00 | 115.13 | 94.99 |
| Длина генеративного побега, см Length of generative shoot, cm |
7.00–(12.69 ± 0.17)–21.00 | 4.50–(15.78 ± 0.08)–25.00 | 22.09 | 15.81 |
| Длина внешней доли околоцветника, см Length of the outer perianth lobe, cm |
2.49–(4.26 ± 0.04)–6.48 | 3.00–(4.83 ± 0.02)–8.00 | 15.47 | 12.67 |
| Ширина внешней доли околоцветника, см Width of the outer perianth lobe, cm |
0.51–(1.28 ± 0.02)–2.14 | 0.80–(1.42 ± 0.01)–2.80 | 19.66 | 17.35 |
| Длина внутренней доли околоцветника, см Length of the inner perianth lobe, cm |
1.93–(3.92 ± 0.04)–5.53 | 2.10–(4.67 ± 0.02)–7.00 | 17.74 | 14.14 |
| Ширина внутренней доли околоцветника, см Width of the inner lobe of the perianth, cm |
0.46–(1.27 ± 0.02)–4.26 | 0.70–(1.47 ± 0.01)–2.60 | 26.71 | 17.53 |
| Диаметр цветка, см Flower diameter, cm |
3.00–(6.36 ± 0.07)–10.00 | 2.60–(5.26 ± 0.03)–9.30 | 17.41 | 17.88 |
Коэффициент вариации был заметно выше в выборке популяций из Нижнего Поволжья по пяти признакам: числу вегетативных побегов, ширине второго листа, числу и длине генеративных побегов, ширине внутренней доли околоцветника. По остальным признакам (диаметру клона, числу листьев на вегетативный побег, длине второго листа, длине и ширине внешней доли околоцветника, длине внутренней доли околоцветника и диаметру цветка) был близок по обеим выборкам популяций (табл. 3). Максимальные значения коэффициентов вариации в обеих выборках популяций отмечены для трех морфологических признаков: числа генеративных и вегетативных побегов и диаметра клона. В целом популяции Нижнего Поволжья имели более широкий диапазон изменчивости основной части исследованных морфологических признаков.
Диаграммы размаха (рис. 3) также наглядно демонстрируют, что средние значения и ошибки средних по большинству признаков в двух сравниваемых географических выборках заметно отличаются между собой. Лишь по числу генеративных и вегетативных побегов сравниваемые выборки достоверно, согласно тесту Манна–Уитни, не различаются (табл. 4). При этом величины дисперсии по большинству показателей для каждого отдельного признака между выборками популяций практически одинаковы. Исключение составили следующие показатели: число вегетативных побегов, число генеративных побегов, длина и ширина второго листа, а также ширина внутренней доли околоцветника. По первому из перечисленных признаков различие в дисперсии между рассматриваемыми выборками наиболее заметно.
Рис. 3.
Диаграммы размаха морфологических показателей Iris pumila: SU – популяционная выборка из Республики Башкортостан; LV – популяционная выборка из Саратовской области.
Fig. 3. Diagrams of the range of morphological parameters of Iris pumila: SU – population sample from the Republic of Bashkortostan; LV – population sample from the Saratov Region.
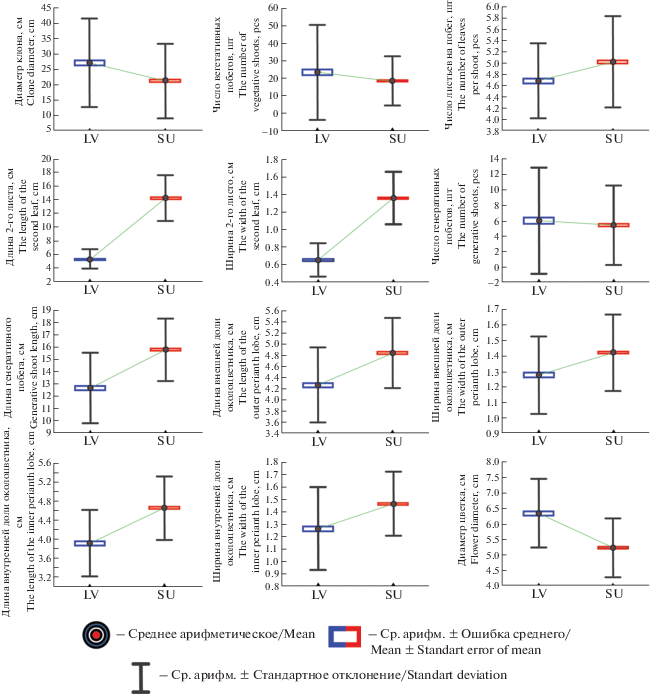
Таблица 4.
Тест Манна–Уитни для двух независимых выборок популяций I. pumila Южного Урала и Нижнего Поволжья Table 4. Mann–Whitney test for two independent samples of I. pumila populations of the South Urals and the Lower Volga region
| Морфологический признак / Morphological trait | p-level |
|---|---|
| Диаметр клона, см / Clone diameter, cm | 0.000 |
| Число вегетативных побегов, шт. / Number of vegetative shoots | 0.938 |
| Число листьев на побег, шт. / Number of leaves per shoot | 0.000 |
| Длина второго листа, см / Length of the second leaf, cm | 0.000 |
| Ширина второго листа, см / Width of the second leaf, cm | 0.000 |
| Число генеративных побегов, шт. / Number of generative shoots | 0.271 |
| Длина генеративного побега, см / Length of generative shoot, cm | 0.000 |
| Длина внешней доли околоцветника, см / Length of the outer perianth lobe, cm | 0.000 |
| Ширина внешней доли околоцветника, см / Width of the outer perianth lobe, cm | 0.000 |
| Длина внутренней доли околоцветника, см / Length of the inner perianth lobe, cm | 0.000 |
| Ширина внутренней доли околоцветника, см / Width of the inner lobe of the perianth, cm | 0.000 |
| Диаметр цветка, см / Flower diameter, cm | 0.000 |
По большинству признаков величины их средних значений выше в выборке растений из популяций Южного Урала. В частности, по показателям листьев (длине и ширине второго листа) и структурных частей цветка (длине и ширине внешней и внутренних долей околоцветника) растения популяций Южного Урала значительно превосходят популяции Нижнего Поволжья. По длине генеративного побега растения из популяций Южного Урала также значительно превосходили таковые из Нижнего Поволжья. Последние лишь по двум параметрам заметно превышали популяции Южного Урала: диаметр клона и диаметр цветка. По показателям числа генеративных и вегетативных побегов выборка из Нижнего Поволжья очень незначительно, но все же превышала выборку растений из популяций Южного Урала.
Таким образом, растения I. pumila из популяций Южного Урала, как по вегетативным, так и по генеративным признакам – цветки и листья, а также генеративные побеги – были крупнее, нежели растения популяций Нижнего Поволжья.
Как уже упоминалось, согласно непараметрическому анализу сравнения средних значений по тесту Манна–Уитни (табл. 4) исследуемые выборки также различались по большинству показателей, кроме числа генеративных побегов и числа вегетативных побегов.
Интересно, что по показателям, которые не выявили достоверного различия между группами, выборки имели наибольшие значения коэффициента вариации, что указывает на высокую внутригрупповую неоднородность особей именно по этим показателям. Неоднородность, в свою очередь, может быть вызвана различными, прежде всего экологическими причинами.
По результатам факторного анализа выделены две главные компоненты, имеющие наибольшие значения собственных чисел и объясняющие наибольший процент дисперсии, как по отдельности (относительно других компонент), так и по общей сумме объясненной дисперсии. Наибольшими нагрузками на первую главную компоненту характеризовались длина второго листа (0.66), ширина второго листа (0.70), длина генеративного побега (0.70), длина внешней доли околоцветника (0.66), длина внутренней доли околоцветника (0.68); по второй главной компоненте наибольшие значения нагрузок имели: диаметр клона (0.82), число генеративных побегов (0.74) (табл. 5). Таким образом, наибольшие нагрузки на выделенные компоненты в более или менее равной степени имели как генеративные, так и вегетативные показатели.
Таблица 5.
Факторный анализ морфологических показателей I. pumila Южного Урала и Нижнего Поволжья Table 5. Factor analysis of morphological parameters of I. pumila of the South Urals and the Lower Volga region
| № | Признак/Trait | Компоненты/Components | |
|---|---|---|---|
| PC 1 | PC 2 | ||
| 1 | Диаметр клона, см / Clone diameter, cm | –0.03 | 0.82 |
| 2 | Число вегетативных побегов, шт. / Number of vegetative shoots | –0.06 | 0.60 |
| 3 | Число листьев на побег, шт. / Number of leaves per shoot | 0.23 | 0.06 |
| 4 | Длина второго листа, см / Length of the second leaf, cm | 0.66 | –0.41 |
| 5 | Ширина второго листа, см / Width of the second leaf, cm | 0.70 | –0.09 |
| 6 | Число генеративных побегов, шт. / Number of generative shoots | 0.05 | 0.74 |
| 7 | Длина генеративного побега, см / Length of generative shoot, cm | 0.70 | 0.05 |
| 8 | Длина внешней доли околоцветника, см / Length of the outer perianth lobe, cm | 0.66 | 0.09 |
| 9 | Ширина внешней доли околоцветника, см / Width of the outer perianth lobe, cm | 0.56 | 0.17 |
| 10 | Длина внутренней доли околоцветника, см / Length of the inner perianth lobe, cm | 0.68 | 0.14 |
| 11 | Ширина внутренней доли околоцветника, см / Width of the inner lobe of the perianth, cm | 0.60 | 0.15 |
| 12 | Диаметр цветка, см / Flower diameter, cm | –0.07 | 0.60 |
| Свойства компонент / Component properties | |||
| 1 | Собственное число / Proper number | 3.05 | 2.22 |
| 2 | Дисперсия, % / Dispersion,% | 25.39 | 18.51 |
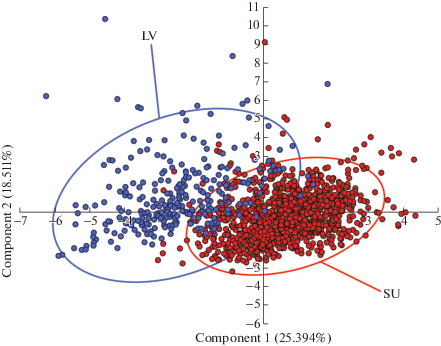
При ординации в пространстве первых двух главных компонент (PCA) (рис. 4) показано разделение исследуемых выборок растений I. pumila из Нижнего Поволжья, с одной стороны, и Южного Урала – с другой, при относительно незначительном перекрывании их облаков рассеяния. При этом обособление облаков рассеяния наиболее выражено относительно горизонтальной оси, характеризующей первую главную компоненту. По этой компоненте обособление точек и их распределение в пространстве в наибольшей степени, как уже было сказано, определяется длиной и шириной второго листа, длиной генеративного побега, а также длинами внешней и внутренней долей околоцветника. В целом, по этим же признакам наблюдалось наибольшее различие исследуемых выборок по диаграммам размаха и описательным статистикам. Именно эти признаки во многом обуславливают различие растений из двух географических регионов и являются ключевыми, т.е. характеризующими общее состояние растений относительно внешних условий. При этом по второй компоненте наблюдается менее заметное обособление точек, обусловленное в большей мере показателями диаметра клона и длины генеративного побега согласно их значениям факторных нагрузок. По диаметру клона и по результатам теста Манна–Уитни, и по диаграммам размаха наблюдается достоверное различие сравниваемых выборок. Что касается длины генеративного побега, то различие выборок по этому параметру не достоверно, как по тесту Манна–Уитни, так и по диаграммам размаха. Дисперсия точек в пространстве области ординации по второй компоненте в большей степени характеризует различие особей внутри анализируемых выборок. Последнее утверждение подтверждается и фактически максимальными значениями коэффициентов вариации описываемых показателей внутри исследуемых групп среди всех морфометрических показателей.
Рис. 4.
Ординация Iris pumila методом главных компонент (PCA): SU – популяционная выборка из Республики Башкортостан; LV – популяционная выборка из Саратовской области.
Fig. 4. Ordination of Iris pumila by the principal component analysis (PCA): SU – population sample from the Republic of Bashkortostan; LV – population sample from the Saratov Region.
На диаграмме рассеяния, полученной методом неметрического многомерного шкалирования, помимо точек, обозначающих местоположение исследованных популяций в пространстве двух главных осей, отмечены векторы, соответствующие наиболее скоррелированным с осями переменным окружающей среды, а именно среднегодовой температуре (bio1), среднемесячной суточной амплитуде температуры (bio2), изотермальности (bio3), cезонности температуры (коэффициент вариации) (bio4), минимальной температуре наиболее холодного месяца (bio6), среднегодовой амплитуде колебания температуры (bio7), средней температуре наиболее сухого квартала (bio9), средней температуре наиболее теплого квартала (bio10), средней температуре самого холодного квартала (bio11), а также высоте над уровнем моря (Altitude), широте (lat) и долготе (long) места произрастания популяций (табл. 2; рис. 5). При этом в анализе влияния биоклиматических характеристик на распределение точек в пространстве осей неметрического многомерного шкалирования учитывались только направление и относительная длина соответствующих векторов.
Рис. 5.
Ординация методом неметрического многомерного шкалирования (NMDS). Мера различий – евклидово расстояние; стресс-значение – 0.06. Условные обозначения популяций те же, что в табл. 1. Условные обозначения биоклиматических факторов те же, что в табл. 2. SU – популяции с Южного Урала, LV – популяции из Нижнего Поволжья.
Fig. 5. Ordination by non-metric multidimensional scaling (NMDS). The measure of the differences is Euclidean distance; stress value – 0.06. For the symbols of the populations see Table 1. For the symbols of bioclimatic factors see Table 2. SU – populations from the South Urals, LV – populations from the Lower Volga region.

На рис. 5 видно хорошее расхождение точек, соответствующих популяциям различных географических групп. При этом в правой части области ординации расположились точки, отвечающие популяциям Южного Урала, в левой – популяциям Нижнего Поволжья. Обособление этих двух агрегаций точек более ярко выражено вдоль горизонтальной оси. Характерно, что разброс точек, соответствующих популяциям из Нижнего Поволжья, велик, как вдоль горизонтальной, так и вдоль вертикальной осей неметрического многомерного шкалирования, в то время как точки, соответствующие популяциям Южного Урала, располагаются одной компактной группой в правой части плоскости ординации. Заметно отделилась от них только точка, соответствующая популяции с горы Караултау из Мелеузовского р-на (Kar), являющаяся самой северной как для Южного Урала, так и для всей выборки в целом (рис. 5). В целом это указывает на большую неоднородность условий произрастания популяций I. pumila в Нижнем Поволжье и относительную однородность таковых на Южном Урале. Нельзя не отметить и тот факт, что в пространстве координат разброс точек, соответствующих совокупности средних значений морфологических признаков в популяциях Нижнего Поволжья, подчиняется определенной закономерности. Вдоль горизонтальной оси точки, соответствующие популяциям юга Заволжья, расположились слева (Mar, Sol-1, Ptr, Stp), а точки, соответствующие популяциям Правобережья и севера Заволжья, – справа (ближе к точкам, соответствующим популяциям Южного Урала), причем среди последних вдоль вертикальной оси выше нулевой отметки расположились точки, соответствующие популяциям Правобережья (Bel, Kam, Sim) и самой западной из левобережных популяций, находящейся на излучине Волги (Uzm), а ниже – популяциям севера Заволжья (Mxp, Trm, Sol-2). При этом к последним ближе всего расположены точки, соответствующие южным популяциям Правобережья (Sim, Kam). Выше расположилась точка, соответствующая самой западной из левобережных популяций, произрастающей в излучине Волги (Uzm), а еще выше – точка, соответствующая популяции севера Правобережья (Bel). То есть, в целом расположение точек в пространстве осей неметрического многомерного шкалирования соответствует географическому градиенту распределения популяций, которым эти точки соответствуют.
Из графика хорошо видно, что переменные окружающей среды достаточно сильно скоррелированы с горизонтальной осью – о чем свидетельствует их длина и соответствующее направление. Напрашивается вывод о том, что за обособление вдоль горизонтальной оси популяций Нижнего Поволжья ответственны: среднемесячная суточная амплитуда температуры (bio2), изотермальность (bio3), сезонность температуры (коэффициент вариации) (bio4), среднегодовая амплитуда колебания температуры (bio7) и высота над уровнем моря (Altitude), а за обособление популяций Южного Урала – среднегодовая температура (bio1), минимальная температура наиболее холодного месяца (bio6), средняя температура наиболее сухого квартала (bio9), средняя температура наиболее теплого квартала (bio10), средняя температура самого холодного квартала (bio11), количество осадков самого холодного квартала (bio19), географическая широта (Latitude) и географическая долгота (Longitude). При этом первые пять переменных, ответственных за обособление популяций Нижнего Поволжья, скоррелированы с горизонтальной осью отрицательно, а из оставшихся восьми переменных, связанных с обособлением популяций Южного Урала, шесть – положительно, а две – отрицательно. Нельзя не заметить, что все биоклиматические характеристики, ответственные за обособление популяций Нижнего Поволжья, по сути, отражают амплитуду колебаний температуры, т.е. диапазон изменчивости этого параметра, в то время как биоклиматические характеристики, ответственные за обособление популяций Южного Урала, отражают средние, и лишь по одному параметру минимальные температуры в регионе, а также – количество осадков в холодный квартал.
Разброс точек по вертикальной оси неметрического многомерного шкалирования определяет, в большей мере, средняя температура наиболее сухого квартала, географическая долгота и высота над уровнем моря. Это следует из того, что направления векторов именно этих биоклиматических и географических факторов отклоняются в пространстве неметрического многомерного шкалирования в сторону вертикальной оси.
Таким образом, больший разброс точек, соответствующих совокупности средних значений морфологических признаков растений в популяциях Нижнего Поволжья, подтверждает большую неоднородность условий произрастания вида в этой географической области по сравнению с Южным Уралом. Наиболее различными по параметрам средней температуры самого сухого квартала (bio9) и высоты над уровнем моря мест произрастания (Altitude) являются места произрастаний популяций Bel (bio9 = –5.08°C, Altitude = 229 м) и Trm (bio9 = –12.78°C, Altitude = = 311 м) (табл. 6). Следовательно, из всех климатических и географических характеристик, участвующих в анализе, именно данные параметры оказывают наибольшее влияние на распределение изменчивости морфологических признаков популяций Нижнего Поволжья. Соответственно, в местообитаниях на Южном Урале, прежде всего, меньшая амплитуда температур и не столь значительный, как в Поволжье, разброс по географической долготе и высоте над уровнем моря мест произрастания приводят к близости морфологических параметров растений изучаемого вида.
Таблица 6.
Значения переменных окружающей среды, высоты над уровнем моря, широты и долготы, векторы которых спроецированы на оси ординации NMDS, согласно местам нахождения популяций в Нижнем Поволжье и Южном Урале Table 6. Values of environmental variables, altitude, latitude, and longitude, the vectors of which are projected onto NMDS ordination axes, in accordance with the locations of the populations in the Lower Volga region and in the South Urals
| Reg | ID | bio1 | bio2 | bio3 | bio4 | bio6 | bio7 | bio9 | bio10 | bio11 | bio19 | Altitude | Latitude | Longitude |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| SU | Taz | 5.43 | 9.70 | 21.85 | 1312.60 | –16.20 | 44.40 | 5.97 | 20.92 | –10.87 | 102 | 49 | 52.185807 | 56.699793 |
| SU | Vmu | 5.59 | 9.38 | 21.60 | 1286.35 | –15.40 | 43.40 | –3.00 | 20.85 | –10.32 | 116 | 66 | 52.017090 | 56.754400 |
| SU | Kuz | 5.83 | 9.65 | 21.98 | 1291.87 | –15.40 | 43.90 | –2.85 | 21.15 | –10.13 | 112 | 84 | 51.574653 | 57.197240 |
| SU | Ars | 6.36 | 9.95 | 22.46 | 1293.96 | –15.00 | 44.30 | –2.37 | 21.75 | –9.62 | 76 | 58 | 51.643555 | 56.787757 |
| SU | Kut | 5.79 | 10.28 | 22.48 | 1322.83 | –16.40 | 45.70 | –3.17 | 21.47 | –10.58 | 90 | 87 | 51.905430 | 56.780569 |
| SU | Vsk | 6.79 | 9.32 | 22.08 | 1241.56 | –13.40 | 42.20 | –1.45 | 21.63 | –8.48 | 98 | 42 | 52.217500 | 56.479074 |
| SU | Len | 5.62 | 9.48 | 22.53 | 1231.84 | –14.50 | 42.10 | –2.65 | 20.32 | –9.55 | 94 | 267 | 52.801797 | 55.611802 |
| SU | Nbl | 5.69 | 9.37 | 22.46 | 1220.07 | –14.50 | 41.70 | –2.50 | 20.15 | –9.40 | 92 | 137 | 52.411747 | 55.598153 |
| SU | Hlk | 6.51 | 9.43 | 21.99 | 1263.73 | –14.00 | 42.90 | –1.92 | 21.60 | –9.05 | 114 | 62 | 52.712637 | 55.609113 |
| SU | Yak | 6.07 | 9.33 | 21.79 | 1266.41 | –14.50 | 42.80 | 6.40 | 21.17 | –9.55 | 148 | 73 | 52.582541 | 55.643009 |
| SU | Kar | 4.89 | 9.18 | 21.41 | 1270.86 | –15.90 | 42.90 | –3.55 | 19.98 | –10.83 | 125 | 181 | 52.989546 | 56.468915 |
| LV | Ptr | 3.39 | 11.25 | 23.78 | 1358.69 | –19.70 | 47.30 | –12.40 | 19.28 | –13.67 | 84 | 266 | 50.785110 | 47.824395 |
| LV | Stp | 3.47 | 10.83 | 23.33 | 1335.56 | –19.30 | 46.40 | –5.18 | 19.08 | –13.33 | 97 | 255 | 51.36015 | 46.90591 |
| LV | Bel | 3.66 | 10.88 | 23.39 | 1342.18 | –19.10 | 46.50 | –5.08 | 19.35 | –13.22 | 94 | 229 | 52.47863 | 48.05778 |
| LV | Mar | 3.22 | 11.35 | 23.89 | 1363.78 | –20.00 | 47.50 | –12.65 | 19.13 | –13.93 | 82 | 291 | 50.761518 | 48.840204 |
| LV | Sol-1 | 3.30 | 11.44 | 23.99 | 1367.68 | –20.00 | 47.70 | –12.62 | 19.30 | –13.87 | 80 | 286 | 51.488845 | 48.078042 |
| LV | Kam | 3.76 | 11.50 | 24.01 | 1374.05 | –19.60 | 47.90 | –12.27 | 19.85 | –13.48 | 76 | 234 | 50.71152 | 45.27483 |
| LV | Sim | 3.44 | 11.70 | 24.22 | 1381.33 | –20.10 | 48.30 | –12.67 | 19.60 | –13.88 | 74 | 274 | 51.34137 | 44.83986 |
| LV | Mxp | 3.18 | 10.83 | 23.35 | 1335.92 | –19.60 | 46.40 | –5.48 | 18.78 | –13.63 | 100 | 301 | 51.85929 | 49.62207 |
| LV | Sol-2 | 4.13 | 10.86 | 23.35 | 1343.48 | –18.60 | 46.50 | –4.63 | 19.83 | –12.77 | 89 | 158 | 52.04402 | 48.34098 |
| LV | Uzm | 3.25 | 11.12 | 23.80 | 1340.90 | –19.70 | 46.70 | –5.42 | 18.88 | –13.65 | 91 | 227 | 51.22534 | 45.93035 |
| LV | Trm | 3.21 | 11.48 | 24.01 | 1372.04 | –20.10 | 47.80 | –12.78 | 19.27 | –14.00 | 79 | 311 | 51.813144 | 47.248446 |
ЗАКЛЮЧЕНИЕ
Растения популяций Iris pumila Южного Урала по большинству исследованных морфологических признаков достоверно отличаются от растений этого вида, произрастающих в Нижнем Поволжье. Растения Южного Урала крупнее по высоте генеративного побега, структурным элементам цветка и, в особенности, по параметрам листа. В свою очередь, растения из популяций Нижнего Поволжья превосходят по показателям диаметра цветка и диаметра клона. Последний признак характеризуется высокой вариативностью. Лишь по двум показателям, – количеству вегетативных и генеративных побегов, – различия между двумя географическими выборками популяций не были установлены. Эти два параметра, также как и диаме тр клона, обладали высоким, по сравнению с остальными показателями, уровнем варьирования.
Ординация методом главных компонент, выделенных в ходе факторного анализа, также убедительно демонстрирует разделение популяционных выборок из двух географических регионов. При этом, наиболее показательно обособление по первой компоненте, которое, согласно значениям факторных нагрузок, определяется длиной и шириной второго листа, длиной генеративного побега, а также длинами внешней и внутренней долей околоцветника. По этим признакам отмечено наибольшее различие исследуемых выборок согласно диаграммам размаха и описательным статистикам. По второй компоненте наблюдается менее заметное обособление точек, обусловленное в большей мере показателями диаметра клона и длины генеративного побега.
Анализ непараметрического многомерного шкалирования также показал сильное разделение точек, отвечающих той или иной географической группе популяций в пространстве ординации. Больший разброс точек, соответствующих совокупности средних значений морфологических признаков растений в популяциях Нижнего Поволжья, указывает на большую неоднородность условий произрастания вида в этой географической области по сравнению с Южным Уралом. В Нижнем Поволжье на распределении изменчивости морфологических признаков, прежде всего, сказывается большая амплитуда температур (в крайних границах, вероятно, приводящая в летний сезон к засухе и в зимний – к глубокому промерзанию почвы). Этому способствует и высота над уровнем моря мест произрастания популяций в данном географическом регионе. На Южном Урале сходство морфологических параметров растений вида обусловлено, прежде всего, небольшой амплитудой температур, как в отдельные месяцы, так и между сезонами, независимо от пространственного положения популяций в данной географической области.
Список литературы
[Abramova et al.] Абрамова Л.М., Широких П.С., Голованов Я.М., Мустафина А.Н., Крюкова А.В. 2019. К экологии редких степных видов рода Iris на Южном Урале. – Вестник Томского государственного университета. Биология. 48: 56–72. https://doi.org/10.17223/19988591/48/3
[Alekseyeva] Алексеева Н.Б. 2008. Род Iris (Iridaceae) в России. – Turczaninowia. 11 (2): 5–68.
Avramov S., Miljković D., Barišić Klisarić N., Živković U., Tarasjev A. 2017. Ontogenetic plasticity of anatomical and ecophysiological traits and their correlations in Iris pumila plants grown in contrasting light conditions. – Plant Species Biol. 32 (4): 392–402. https://doi.org/10.1111/1442-1984.12171
Barišić Klisarić N., Avramov S., Miljković D., Živković U., Tarasjev A. 2012. Ontogeny of flower parts on naturally growing Iris pumila clones: implications for population differentiation and phenotypic plasticity studies. – Russian Journal of Genetics. 48 (4): 556–560.
Biye E.H., Cron G.V., Balkwill K. 2016. Morphometric delimitation of Gnetum species in Africa. – Plant Syst. Evol. 302: 1067–1082. https://doi.org/10.1007/s00606-016-1317-3
Boltenkov E.V. 2019. Typification of the Linnaean name Iris pumila (Iridaceae). –Willdenowia. 49 (2): 147–150. https://doi.org/10.3372/wi.49.49202
Cruz-Lustre G., Batista J.A.N., Radins J.A., Gonzalez A., Borba E.L. 2020. Morphometric analysis of the Habenaria parviflora complex (Orchidaceae). – Plant Syst. Evol. 306. Published online. https://doi.org/10.1007/s00606-020-01634-2
Fick S.E., Hijmans R.J. 2017. WorldClim 2: new 1km spatial resolution climate surfaces for global land areas. – International Journal of Climatology. 37 (12): 4302–4315.
Finot V.L., Soreng R.J., Giussani L.M., Munoz R.G. 2018. A multivariate morphometric delimitation of species boundaries in the South American genus Nicoraepoa (Poaceae: Pooideae: Poeae). – Plant Syst. Evol. 304: 679–697. https://doi.org/10.1007/s00606-018-1499-y
Jolliffe I.T. 2002. Principal Component Analysis, 2nd edn. New York. 487.
[Khar’kova, Grzhibovskiy] Харькова О.А., Гржибовский А.М. 2014. Сравнение двух несвязанных выборок c использованием пакета статистических программ Stata: непараметрические критерии. – Экология человека. 4: 60–64.
[Krasnaya…] Красная книга Республики Башкортостан: в 2 т. Т. 1. Растения и грибы. 2011. Уфа. 384 с.
[Krasnaya…] Красная книга Российской Федерации (растения и грибы). 2008. М. 855 с.
[Krasnaya…] Красная книга Саратовской области: Грибы. Лишайники. Растения. Животные. 2021. Саратов. 496 с.
[Kryukova et al.] Крюкова А.В., Абрамова Л.М., Мустафина А.Н. 2018. К биологии и экологии редких видов ирисов в степях Южного Урала. – Самарская Лука: проблемы региональной и глобальной экологии. 27 (4–1): 271–275. https://doi.org/10.24411/2073-1035-2018-10126
[Kryukova et al.] Крюкова А.В., Мулдашев А.А., Голованов Я.М., Абрамова Л.М. 2014. Распространение и фитоценотическая приуроченность редких видов рода Iris L. на Южном Урале. – Научные ведомости Белгородского государственного университета. Сер. Естественные науки. 23 (29): 5–11.
[Kryukova, Abramova] Крюкова А.В., Абрамова Л.М. 2018. Влияние экологических факторов на изменчивость морфометрических параметров редкого вида Iris pumila L. – Изв. Сарат. ун-та. Нов. сер. Сер. Химия. Биология. Экология. 18 (2): 232–236. https://doi.org/10.18500/1816-9775-2018-18-2-232-236
Maia F.R., Goldenberg R. 2019. Morphometric analysis and the distinction between Tibouchina hatschbachii and T. marumbiensis: morphological differentiation driven from the past. – Plant Syst. Evol. 305: 169–180. https://doi.org/10.1007/s00606-018-1560-x
[Markov, Ivnitskiy] Марков А.В., Ивницкий С.Б. 2016. Эволюционная роль фенотипической пластичности. – Вест. Моск. ун-та. Сер. 16. Биология. 4: 3–11.
[Metody…] Методы изучения ценопопуляций цветковых растений. 2015. Саратов. 127 с.
Nobis M., Klichowska E., Nowak A., Gudkova P.D., Rola K. 2016. Multivariate morphometric analysis of the Stipa turkestanica group (Poaceae: Stipa sect. Stipa). – Plant Syst. Evol. 302: 137–153. https://doi.org/10.1007/s00606-015-1243-9
Ocampo J., d’Eeckenbrugge G. 2017. Morphological characterization in the genus Passiflora L.: an approach to understanding its complex variability. – Plant Syst. Evol. 303: 531–558. https://doi.org/1007/s00606-017-1390-2
Oksanen J., Blanchet F.G., Friendly M., Kindt R., Legendre P., McGlinn D., Minchin P.R., O’Hara R.B., Simpson G.L., Solymos P., Stevens M.H.H., Szoecs E., Wagner H. 2020. vegan: Community Ecology Package. R package version 2.5–7. https://CRAN.R-project.org/package=vegan (дата обращения 15.03.2021).
Pemac D., Tucić B. 1998. Reaction norms of juvenile traits to light intensity in Iris pumila (Iridaceae): A comparison of populations from exposed and shaded habitats. – Plant Syst. Evol. 209: 159–176.
R Core Team. R: A language and environment for statistical computing. R Foundation for Statistical Computing. Vienna, Austria. 2020. Available at: http://www.R-project.org (дата обращения 15.03.2021).
Sharma S.K., Pandit M.K. 2011. A morphometric analysis and taxonomic study of Panax bipinnatifidus Seem. (Araliaceae) species complex from Sikkim Himalaya, India. – Plant Syst. Evol. 297: 87–98. https://doi.org/10.1007/s00606-011-0501-8
Spaniel S., Zozomova-Lihova J., Marhold K. 2017. Revised taxonomic treatment of the Alyssum montanum-A. repens complex in the Balkans: a multivariate morphometric analysis. – Plant Syst Evol. 303: 1413–1442. https://doi.org/10.1007/s00606-017-1468-x
STATISTICA (data analysis software system), version 13 // StatSoft, Inc., 2001. http://www.statsoft.com (дата обращения 15.03.2021).
Tarasjev A., Avramov S. & Miljković D. 2012. Evolutionary biology studies on the Iris pumila clonal plant: advantages of a good model system, main findings and directions for further research. – Archives of Biological Sciences. 64: 159–174.
Tarasjev A., Barisić Klisarić N., Stojković B., Avramov S. 2009. Phenotypic plasticity and between population differentiation in Iris pumila transplants between native open and anthropogenic shade habitats. – Гeнeтикa. 45 (8): 1078–1086.
Torrecilla P., Acedo C., Marques I., Diaz-Pérez A. J., López-Rodriguez J. A., Mirones V., Sus A., Llamas F., Alonso A., Pérez-Collazos E., Viruel J., Sahuquillo Sancho M. D., Komac B., Manso J. A., Segarra-Moragues J. G., Draper D., Villar L., Catalán P. 2013. Morphometric and molecular variation in concert: taxonomy and genetics of the reticulate Pyrenean and Iberian alpine spiny fescues (Festuca eskia complex Poaceae). – Bot. J. Linn. Soc. 173: 676–706. https://doi.org/10.1111/boj.12103
Tucić B., Pemac D., Avramov S. 2003. Plasticity to daylength of Iris pumila leaf phenological traits. – Popul. Ecol. 45: 31–39. https://doi.org/10.1007/s10144-003-0137-9
Wickham H. 2016. ggplot2: Elegant Graphics for Data Analysis. New York. 268 p.
[Yefimov et al.] Ефимов С.В., Чернышенко О.В., Кирпичева Л.Ф., Дацюк Е.И. 2012. Крымские популяции ириса карликового (Iris pumila L.): распространение и морфологические особенности. – Лесной вестник. 4: 7–12.
Yoccoz N.G. 2012. Ecological monitoring. – In: Encyclopedia of Life Sciences. Published online. https://doi.org/10.1002/9780470015902.a0023571
[Yuritsyna, Vasyukov] Юрицына Н.А., Васюков В.М. 2014. Новое местонахождение Iris pumila L. (Iridaceae) на юге Приволжской возвышенности. – Известия Самарского научного центра Российской академии наук. 16 (5): 143–144.
Zar J.H. 2010. Biostatistical analysis. New Jersey. 944 p.
Zuur A.F., Ieno E.N., Elphick C.S. 2009. A protocol for data exploration to avoid common statistical problems. – Methods in Ecology and Evolution. 1: 3–14. https://doi.org/10.1111/j.2041-210x.2009.00001.x
Дополнительные материалы отсутствуют.
Инструменты
Ботанический журнал