

Ботанический журнал, 2022, T. 107, № 4, стр. 360-384
ГОДИЧНЫЙ ПРИРОСТ РАННЕЙ И ПОЗДНЕЙ ДРЕВЕСИНЫ В ЛОКАЛЬНЫХ УЧАСТКАХ КАМБИАЛЬНОЙ ЗОНЫ СТВОЛА КАК ПОКАЗАТЕЛЬ ЭТАПОВ РОСТА И РАЗВИТИЯ ДЕРЕВА QUERCUS ROBUR (FAGACEAE)
А. В. Тихомиров *
* E-mail: dendrodoctor@yandex.ru
Поступила в редакцию 25.05.2018
После доработки 19.01.2022
Принята к публикации 25.01.2022
- EDN: MGJUXL
- DOI: 10.31857/S000681362204007X
Аннотация
В стволе Quercus robur L. в течение жизненного цикла локального участка камбиальной зоны доля поздней древесины относительно ранней древесины в годичном кольце уменьшается, стремясь к нулю. С учетом этого фактора введены понятия контрольного и нормированного возраста, характеризующие возрастные этапы жизнедеятельности участка камбиальной зоны. Описан способ их определения. Продолжительность жизнедеятельности участка камбиальной зоны может быть в пределах от нескольких десятков до сотен лет. Для сравнительной характеристики соотношения поздней и ранней древесины в однозначные нормированные периоды развития камбиальных зон выделено пять возможных вариантов, которые могут быть выражены в балльной системе. После начального периода закономерного увеличения годичного прироста ранней древесины по радиусу и по площади характер изменения их трендов может быть различен. В пределах ствола рост и развитие локальных участков камбиальных зон находится под общим контролем целостной системы дерева. Показатели контрольного и нормированного возраста камбиальных зон позволяют оценить особенности состояния дерева в различные периоды его жизни.
Задача оценки состояния деревьев является одной из важнейших в лесоводстве. Знание о возможной продолжительности жизни конкретного дерева и признаков, характеризующих различные временные периоды жизни, позволяет более точно оценивать его текущее состояние. В процессе своего развития дерево проходит различные этапы: молодость, зрелость, старение, которые отражаются на всех его структурных и функциональных уровнях (Chailakhyan, 1980).
Годичный радиальный прирост древесины ствола представляет одну из наиболее информативных структур, фиксирующих состояние дерева. Как отмечает К. Эзау (Ezau, 1980), анализ серийных срезов годичных колец древесины позволяет получить представление о состоянии и особенностях поведения камбия в прошлом.
В течение жизни дерева в структуре древесины происходят направленные изменения. В первые 10–20 лет формируется молодая древесина, затем зрелая (Zobel, Sprague, 1998), изменения продолжаются и в дальнейшем (Leprovost, Stokes, 2001).
Вопросы оценки этапов онтогенеза и продолжительности жизни дерева тесно взаимосвязаны. Одним из признаков скорого отмирания дерева является заметное уменьшение годичного прироста диаметра ствола. Его значение, равное 0.1 мм, было предложено в качестве тестового признака, свидетельствующего о вероятном усыхании дерева (Botkin et al., 1972). Различия в годичном приросте древесины у отмерших и оставшихся в живых деревьев могут наблюдаться за несколько десятилетий до момента усыхания (Bigler, Bugmann, 2004). Вместе с тем, диагностика природного потенциала, устойчивости и продолжительности жизни дерева остается малоизученной областью лесоведения (Dobbertin, 2005; Rohner , 2012).
Объектом наших исследований являлся анализ годичного прироста ранней и поздней древесины в стволах дуба черешчатого (Quercus robur L.).
В годичном приросте ствола дуба черешчатого ранняя древесина состоит из крупных сосудов, расположенных в один или несколько слоев, аксиальной паренхимы, вазицентрических трахеид, могут присутствовать древесные волокна. Поздняя древесина включает в свой состав древесные волокна, волокнистые трахеиды, клетки паренхимы, мелкие сосуды (Vikhrov, 1953; Yatsenko-Khmelewski, Kobak, 1978).
Формирование ранней древесины происходит большей частью за счет питательных веществ, накопленных растением в прошлом сезоне. Формирование поздней древесины идет за счет ассимилянтов текущего года и продолжается до конца августа – начала сентября (Vikhrov, 1954; Grud-zinskaya, 1962; Elagin, 1962, и др.).
Радиальный годичный прирост поздней древесины дуба проявляет более высокую изменчивость, чем ранняя древесина (Yakovlev, 1949; Samtsov, 1966; Rusalenko, 1986, и др.). В.К. Ширнин (Shirnin, 1986) отнес ширину годичного слоя и долю (%) поздней древесины к признакам со слабой генетической обусловленностью. По мнению Н.Е. Косиченко (Kosichenko, 1999) изменчивость ширины ранней древесины в течение онтогенеза у кольце-сосудистых растений выражена в меньшей степени и находится под более жестким генетическим контролем, чем ранняя древесина у хвойных деревьев, и в эволюционном плане является более прогрессивной частью годичного кольца.
В процессе старения у дуба снижается доля поздней древесины в годичном кольце (Vikhrov, 1954; Mikhailov, 1959; Chevedaev, 1963; Kosichenko, 1999, и др.). При ухудшении его состояния наблюдается уменьшение радиального прироста и процента поздней древесины (Savina, 1941, 1956; Yakovlev, 1949; Vakin, 1954; Vikhrov, 1954 и др.).
С возрастом изменяются темп и особенности функционирования эмбриональных тканей в различных частях дерева, меняются соотношения анатомических элементов (Raskatov, 1950, 1956; Palandzhyan, 1989).
В течение жизнедеятельности камбиальной зоны характер трендов радиального годичного прироста ранней и поздней древесины изменяются соответственно “кривой биологического роста”: вначале они увеличиваются, а затем уменьшаются (Vikhrov, 1954; Shiyatov, 1973, 1986). При этом различные воздействия, связанные с циклами солнечной активности, климатическими аномалиями, поражением от вредителей вызывают продолжительные (до десятилетия и более) отклонения прироста от усредненного его значения (Lovelius, 1979, Bigler, 2003).
Согласно теории циклического старения и омоложения Н.П. Кренке (Krenke, 1940), растение и формирующиеся в процессе роста и развития новые его части изначально обладают определенным потенциалом жизнеспособности и периодом развития. Этот вывод справедлив как для метамеров, образованных в результате деятельности претерпевающей циклически прогрессивное старение верхушечной апикальной меристемы побега, так и для участков камбия, формируемых латеральной меристемой.
Цель работы – анализ общих закономерностей годичного прироста ранней и поздней древесины и их взаимосвязи в стволе дуба черешчатого (Quercus robur) в процессе его онтогенеза и разработка на их основе метода возрастной диагностики состояния дерева.
МАТЕРИАЛ И МЕТОДИКА
Структуру радиального годичного прироста дуба черешчатого изучали на спилах модельных деревьев, взятых на разной высоте ствола и кернах, взятых приростным буравом из ствола на высоте 1.3 м. Периметры стволов, из которых отбирали керны и выпилы, не имели ран и открытых дупел. На спилах и кернах последовательное измерение радиальных значений ранней и поздней древесины для каждого годичного кольца проводили за весь период роста камбиальной зоны, начиная от сердцевины до последнего года прироста. Измерение проводили под бинокулярной лупой с точностью до 0.05 мм. На спилах модельных деревьев общий для каждого годичного кольца размер радиальных приростов ранней и поздней древесины определяли как среднее из четырех их значений, полученных при анализе крестообразно расположенных от сердцевины направлений. Суммируя полученные средние радиальные размеры годичных приростов ранней и поздней древесины до конкретного годичного кольца, определяли его расстояние по радиусу от сердцевины. Используя эти данные, рассчитывали по формуле площади круга значения площадей поперечного сечения, как до текущего фрагмента анализируемого годичного кольца, так и включая прирост ранней или поздней древесины в этом годичном кольце. Рассчитав разницу между ними, получали значение площади годичного прироста ранней или поздней древесины в годичном кольце для всего периметра на определенной высоте ствола. Рассчитывая последовательно от первого годичного кольца до последнего, получали ежегодные значения прироста площадей ранней и поздней древесины и годичного кольца в целом за весь период роста камбиальной зоны.
Основной объем исходного полевого материала был собран в Теллермановском опытном лесничестве Воронежской области в период с 1986 по 1992 год. Кроме того, взяты 3 модельных дерева в Шацком лесничестве Рязанской области и керны у 4 дубов в Воронежском лесничестве Воронежской области. Всего было взято 19 модельных деревьев, 97 выпилов и 17 кернов.
При анализе данных были использованы как абсолютные значения годичных радиальных приростов ранней и поздней древесины, так и их относительные величины.
Для характеристики соотношения в годичном кольце радиальных размеров ранней и поздней древесины определялось соотношение ширины поздней и ранней древесины. Оно рассчитывалось как отношение величины радиального прироста поздней древесины к величине прироста ранней древесины.
Для сравнительного анализа показателей радиального прироста в локальных камбиальных зонах абсолютные значения признака переводили в относительные величины по отношению к норме, характерной для данного признака. В качестве базового значения для такой нормы конкретного ряда годичных радиальных приростов ранней древесины мы принимали среднюю из 2–3-х последовательных максимальных величин радиальных приростов, фиксирующих окончание периода начального закономерного ее увеличения, по отношению к которой рассчитывались остальные значения.
Так как в течение жизнедеятельности участка камбиальной зоны наблюдается устойчивая тенденция к понижению соотношения ширины поздней и ранней древесины, то единый тренд такого соотношения может быть использован для оценки динамики функционирования камбиальной зоны.
Для расчета единого тренда соотношения ширины поздней и ранней древесины в течение жизнедеятельности участка камбиальной зоны были использованы алгоритмы регрессионных уравнений логарифма, полинома и экспоненты из программы Statistica 7 (StatSoft Statistica 7.0.61.0 En, 2006). Как правило, только часть годичного тренда, рассчитанного по этим алгоритмам, адекватно описывает исходное соотношение ширины поздней и ранней древесины. Регрессионные уравнения логарифма или полинома применяли для описания начальных этапов роста, а уравнение экспоненты – для последующего заключительного периода деятельности камбиальной зоны, учитывая, что в состоянии, близком к отмиранию, поздняя древесина почти не образуется и значение соотношения ширины поздней и ранней древесины стремится к нулю. Для получения единого тренда за весь период жизнедеятельности камбиальной зоны часть годичных значений начального периода роста, полученных на основе уравнений логарифма или полинома, соединяли с годичными значениями на заключительном периоде роста, рассчитанными по уравнению экспоненты. При этом критерием объединения двух этих трендов конкретного года служат одинаковые для них годичные значения соотношения ширины поздней и ранней древесины.
Список введенных автором терминов и параметров
Соотношение ширины поздней и ранней древесины – отношение радиального прироста поздней древесины к приросту ранней древесины в годичном кольце.
Контрольный возраст (Аcon) – возраст участка камбиальной зоны, при котором среднее значение соотношения ширины поздней и ранней древесины равно 0.4. В процессе жизнедеятельности камбиальной зоны доля поздней древесины в годичном кольце уменьшается, приближаясь к нулю. При данном значении соотношения ширины поздней и ранней древесины наблюдается наибольшая вероятность отмирания дерева.
Нормированный возраст (Аstand.) – возраст годичного кольца в относительных величинах, определяемый как частное от деления возраста конкретного годичного кольца (Aгодичного кольца) на величину контрольного возраста (Аcon).
(1)
${{A}_{{{\text{stand}}{\text{.}}}}}{\text{}} = {\text{}}\frac{{{{А}_{{{\text{годичного кольца}}}}}}}{{{{A}_{{{\text{con}}}}}}}.$Вариант взаимосвязи соотношения ширины поздней и ранней древесины и нормированного возраста. – Вариант тренда взаимосвязи соотношения ширины поздней и ранней древесины, наблюдаемый в камбиальных зонах в течение их жизнедеятельности. В камбиальных зонах тренды могут существенно различаться по таким соотношениям. Для оценки в баллах подобных различий в качестве маркеров выделено 5 трендов возможных вариантов взаимосвязи соотношения ширины поздней и ранней древесины и нормированного возраста.
Реперная зона радиального годичного прироста ранней древесины (Zреперное) – зона, расположенная в верхней точке начального закономерного увеличения радиального годичного прироста ранней древесины. Она включает в себя 2–3 годичных кольца с максимальным приростом.
Реперная зона годичного прироста площади ранней древесины (Sреперное) – зона, которая фиксирует, согласно тренду, окончание начального, закономерного увеличения средней годичной площади ранней древесины, перед последующим снижением такого прироста.
Относительное значение радиального годичного прироста ранней древесины (Zотносительное) – частное от деления величины данного годичного прироста ранней древесины (Zn) на среднее из 2–3 максимальных значений годичного прироста ранней древесины, расположенных в реперной зоне (Zреперное).
(2)
${\text{}}{{Z}_{{{\text{относительное}}}}} = {\text{}}\frac{{{{{\text{Z}}}_{n}}}}{{{{Z}_{{{\text{реперное}}}}}}}.$Относительное значение годичного прироста площади ранней древесины (Sотносительное) – частное от деления величины годичного прироста площади ранней древесины (Sn) на ее значение в реперной зоне (Sреперное).
РЕЗУЛЬТАТЫ И ИХ ОБСУЖДЕНИЕ
I. Локальный участок камбиальной зоны
В течение онтогенеза дуба доля поздней древесины закономерно уменьшается, приближаясь к нулю в момент его отмирания. Годичные значения общего совмещенного тренда соотношения ширины поздней и ранней древесины, полученного по описанной выше методике, характеризуют особенности развития участка камбиальной зоны в течение всего периода его жизнедеятельности. Связь между фактическими годичными значениями соотношения ширины поздней и ранней древесины и общим их трендом оценивали на основе анализа 30 участков камбиальных зон, взятых у 10 моделей в нижней, средней и верхней части ствола. Среднее значение коэффициентов парной корреляции между ними составило: r = 0.83 ± 0.020, что указывает на наличие тесной связи. Среднее значение достоверности коэффициентов регрессии между ними равно: treg = 11.8 ± 1.06 для критерия Стьюдента tstd = = 3.66 при уровне значимости P = 0.001. Существует тесная связь годичных значений общего тренда соотношения ширины поздней и ранней древесины с фактическими годичными значениями за весь период жизнедеятельности участка камбиальной зоны. При этом анализ тренда соотношения ширины поздней и ранней древесины позволяет оценить различные этапы ее развития.
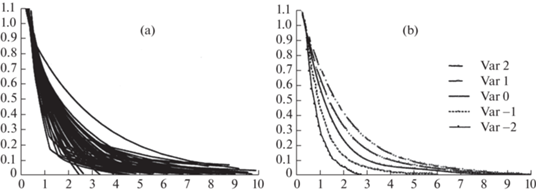
На рис. 1а приведен пример типичного изменения годичного радиального прироста ранней и поздней древесины, соотношения ширины поздней и ранней древесины и их трендов у усохшего дерева на высоте 1.3 м.
Рис. 1.
Радиальный годичный прирост ранней и поздней древесины, соотношение ширины поздней и ранней древесины и их тренды в течение жизнедеятельности участка камбиальной зоны ствола на примере усохшего дерева дуба.
(а) Значения годичного радиального прироста ранней древесины (1), поздней древесины (2), соотношения ширины поздней и ранней древесины (3) и их тренды, рассчитанные по принципу скользящей средней.
(b) Значения соотношения ширины поздней и ранней древесины (1) и их тренд (2), рассчитанный с помощью функций: логарифмической (начальный период роста) и экспоненциальной (последующий период роста).
По оси абсцисс – возраст участка камбиальной зоны, годы; по оси ординат – размер годичного радиального прироста ранней и поздней древесины в мм и значение соотношения ширины поздней и ранней древесины.
Fig. 1. Radial annual increment of earlywood and latewood, latewood/earlywood width ratio and their trends through the lifetime of a local portion of cambium in a selected dead oak tree.
(а) values of the annual radial earlywood increment (1), values of the annual radial latewood increment (2), latewood/earlywood width ratio (3) and their trends, calculated on the basis of moving average method.
(b) values of latewood/earlywood width ratio (1) and their trends (2), calculated using logarithmic (initial growth period) and exponential (subsequent growth period) functions.
X-axis – cambium portion age, years; Y-axis – value of the annual radial earlywood/latewood increment (mm) and the value of latewood/earlywood width ratio.

Линия тренда соотношения ширины поздней и ранней древесины (рис. 1b) характеризует потенциал метаболической активности участка камбиальной зоны на разных этапах жизнедеятельности.
Рассмотрим в год отмирания дерева выровненные по тренду значения общего годичного прироста и соотношение поздней и ранней древесины для 58 участков камбия, расположенных на разной высоте в стволах 12 усохших модельных деревьев дуба черешчатого. Для анализа камбиальных зон у каждой модели выпилы брали из нижней, средней и верхней части ствола. У некоторых модельных деревьев брали промежуточные выпилы. Общее количество участков камбиальных зон в пределах ствола у разных моделей было разным, от 3 до 13. Для определения расположения камбиальной зоны по высоте ствола рассчитывали относительную высоту, как отношение высоты камбиальной зоны к общей длине дерева. Среднее значение относительной высоты для всех камбиальных зон: M = 0.41 ± 0.040. Коэффициенты парной корреляции между относительной высотой дерева и годичным радиальным приростом древесины в год отмирания дерева: r = –0.17 при P = 0.3, между относительной высотой дерева и соотношением ширины поздней и ранней древесины в год отмирания дерева: r = –0.21 при P = 0.18. Следовательно, достоверной связи между данными показателями не обнаружено.
На рис. 2а представлено распределение участков камбиальных зон по размеру годичного прироста в год усыхания дерева. Его значение у большинства участков камбиальных зон (44 из 58) составляет 0.1–0.4 мм. Эти величины несколько выше значения радиального прироста равного 0.05 мм, при котором дерево полностью теряет способность к выживанию (Botkin et al., 1972). Вместе с тем, радиальный прирост менее 0.5 мм свидетельствует о значительном ослаблении устойчивости дерева и увеличении вероятности его отмирания.
Рис. 2.
Показатели радиального годичного прироста для 58 участков камбиальных зон в год отмирания, расположенных на разной высоте ствола у 12 модельных деревьев:
(а) Распределение участков камбиальных зон относительно размера радиального прироста.
По оси абсцисс – размер годичного радиального прироста в мм; по оси ординат – количество участков камбиальных зон в группе.
(b) Отношение значений радиального прироста и соотношения ширины поздней и ранней древесины.
По оси абсцисс – размер годичного радиального прироста в мм; по оси ординат – значение соотношения ширины поздней и ранней древесины.
Fig. 2. Radial annual increments for 58 portions of cambium zones in the year of a tree death, located at different trunk height of 12 model trees:
(а) Distribution of cambium portions with respect to radial increment.
X-axis – annual radial increment (mm); Y-axis – number of cambium portions in the group.
(b) Relationship of radial increments and latewood/earlywood width ratio.
X-axis – annual radial increment (mm); Y-axis – latewood/earlywood width ratio.

На рис. 2b приведено отношение годичного радиального прироста и соотношение ширины поздней и ранней древесины, определенные по характеру тренда в год усыхания дерева. Радиальный прирост в большинстве случаев менее 1.0 мм, а показатель соотношения ширины поздней и ранней древесины менее 0.7. Заметной связи между ними не наблюдается.
Отношение ширины поздней и ранней древесины на момент отмирания участка камбиальной зоны (рис. 3а) варьирует в пределах 0–1.2, средняя величина такого отношения равна 0.48 ± ± 0.027. Наиболее многочисленная группа значений (25% от общего числа) находится в пределах от 0.4 до 0.5. Количество зон прироста со значением менее 0.4 равно 36%.
Рис. 3.
Значения соотношения ширины поздней и ранней древесины для 58 участков камбиальных зон в год отмирания, расположенных на разной высоте ствола у 12 модельных деревьев:
(а) Диаграмма распределения участков камбиальных зон по величине соотношения ширины поздней и ранней древесины.
(b) Распределение участков камбиальных зон по возрасту и величине соотношения ширины поздней и ранней древесины.
По оси абсцисс – значение соотношения ширины поздней и ранней древесины; по оси ординат: (а) количество участков зон камбиального прироста в группе; (б) возраст участка камбиальной зоны, годы.
Fig. 3. Values of the latewood/earlywood width ratio for 58 cambium portions in the year of tree death, located at different trunk height of 12 model trees:
(а) Distribution of cambium portions in respect to the latewood/earlywood width ratio.
(b) Distribution of cambium portions in respect to the age and the latewood/earlywood width ratio.
X-axis – latewood/earlywood width ratio; Y-axis: (а) number of cambium portions in the group; (b) age of cambium portion, years.

На рис. 3b представлено распределение конечных значений величины соотношения ширины поздней и ранней древесины на момент усыхания в зависимости от возраста участков камбиальных зон в диапазоне от 14 до 180 лет. Связи между возрастом участка камбиальной зоны и конечным значением соотношения ширины поздней и ранней древесины на момент усыхания не наблюдается.
Так как тренд соотношения ширины поздней и ранней древесины в течение жизни дерева проявляет явную тенденцию к понижению, стремясь к нулю, то логично связать различные этапы формирования тренда с определенным периодом функционирования участка камбиальной зоны. Используя нормированное время, можно провести сравнительный анализ периодов развития различных участков камбиальных зон.
Для относительной оценки подобных возрастных изменений идеальным временем отсчета является период, в котором участок камбиальной зоны полностью израсходовал свой жизненный потенциал и находится на грани отмирания, а соотношение ширины поздней и ранней древесины по тренду равно нулю. На практике, однако, такие случаи довольно редки, так как часто усыхание происходит на более раннем этапе. В исследуемой группе участков камбиальных зон у отмерших деревьев наибольшее их количество приходится на градацию тренда соотношения ширины поздней и ранней древесины в интервале 0.4–0.5. По данным Н.М. Завады (Zavada, 1983) необратимый процесс усыхания дубов после объедания их листогрызущими насекомыми наступает, когда в течение 3 лет доля ранней древесины в годичном кольце была более двух третей, т.е. соотношение ширины поздней и ранней древесины составляло менее 0.5.
Поэтому, в качестве возрастной точки отсчета, приближенной к максимально ослабленному состоянию участка камбиальной зоны, рассчитанной по тренду, было принято значение соотношения ширины поздней и ранней древесины, равное величине 0.4.
Обозначим возраст локального участка камбиальной зоны в точке тренда соотношения ширины поздней и ранней древесины, равный величине 0.4, как контрольный – Аcon.
Для сравнительной оценки периодов в процессе жизнедеятельности различных участков камбиальных зон введем показатель нормированного возраста – Аstand. Он определяется как частное от деления возраста годичного кольца на величину контрольного возраста:
(4)
${{А}_{{{\text{stand}}{\text{.}}}}} = {\text{}}\frac{{{{А}_{{{\text{годичного кольца}}}}}}}{{{{А}_{{{\text{con}}}}}}}.$В год, когда значение по тренду соотношения ширины поздней и ранней древесины участка камбиальной зоны равно 0.4, нормированный возраст равен 1.0.
Для оценки связи тренда соотношения ширины поздней и ранней древесины с нормированным возрастом был проведен анализ данных участков камбиальных зон, имеющих величину тренда соотношения ширины поздней и ранней древесины равного 0.4.
В графическом виде связь между трендами соотношения ширины поздней и ранней древесины и нормированным возрастом для 49 камбиальных зон изображена на рис. 4а. Она имеет форму, где наибольшие расхождения между ними приходятся на первые периоды жизни участка камбиальной зоны. Все линии трендов стремятся к единой точке, определяемой нормированным возрастом, равным единице. Следует отметить, что существенных взаимных пересечений линий трендов, после прохождения ими зоны нормированного возраста, равного 0.2, относительно мало, (они найдены только в 3 случаях). При этом участки камбиальных зон, имеющие равные значения нормированного возраста, могут существенно различаться по величине соотношения ширины поздней и ранней древесины. В разных камбиальных зонах при нормированном возрасте 0.3 величина соотношения годичного прироста от поздней древесины к ранней может изменяться от 1 до 4.
Рис. 4.
Взаимосвязь значений соотношения ширины поздней и ранней древесины и нормированного возраста в участках камбиальных зон.
(а) Общая совокупность для 49 участков камбиальных зон.
(b) Выделенные 5 вариантов взаимосвязи соотношения ширины поздней и ранней древесины и нормированного возраста.
По оси абсцисс – значение соотношения ширины поздней и ранней древесины; по оси ординат – значение нормированного возраста.
Fig. 4. Relationship of latewood/earlywood ratio to normalized age of cambium portions.
(а) 49 curves for all examined cambium portions.
(b) Five regression lines calculated for different types of relationship between latewood/earlywood ratio and normalized age of cambium.
X-axis – the value of latewood/earlywood ratio; Y-axis – normalized age.
Биологический смысл данного факта указывает на способность участков камбиальных зон осуществлять весь цикл своего развития при различном начальном соотношении годичного радиального прироста поздней и ранней древесины. При этом надо учитывать, что одинаковые значения соотношения ширины поздней и ранней древесины могут характеризовать различные этапы развития камбиальных зон.
Камбиальные зоны в течение жизнедеятельности могут существенно различаться по соотношению поздней и ранней древесины относительно равным значениям нормированного возраста. Для сравнительной оценки значений взаимосвязей соотношения ширины поздней и ранней древесины и нормированного возраста конкретного участка камбиальной зоны в течение его жизнедеятельности было выделено 5 вариантов промежуточных трендов, равномерно распределенных в зоне возможного проявления соотношения поздней и ранней древесины (рис. 4b). Такое количество вариантов, с точки зрения автора, оптимально для последующих расчетов. Находящийся в центре тренд обозначен как “Var 0”, а тренды, расположенные выше и ниже, соответственно, как: “Var 1”, “Var 2” и “Var-1”, “Var-2”.
Ниже для данных вариантов приведены формулы связи нормированного возраста и соотношения ширины поздней и ранней древесины, обозначенного символом “SI” (формулы № 5–9).





В табл. 1 приведены конкретные значения соотношения ширины поздней и ранней древесины для выделенных вариантов при различных значениях нормированного возраста.
Таблица 1.
Варианты взаимосвязи соотношения ширины поздней и ранней древесины и нормированного возраста Table 1. Variants of relationship between of the normalized age value and the latewood/earlywood width ratio
| ВАР VAR |
Значения нормированного возраста/Normalized age value | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| 0.05 | 0.1 | 0.2 | 0.3 | 0.4 | 0.5 | 0.6 | 0.7 | 0.8 | 0.9 | 1.0 | |
| Соотношение ширины поздней и ранней древесины/Latewood/earlywood width ratio | |||||||||||
| Var -2 | 2.08 | 1.76 | 1.33 | 1.08 | 0.92 | 0.82 | 0.73 | 0.64 | 0.54 | 0.44 | 0.4 |
| Var -1 | 3.36 | 2.42 | 1.8 | 1.44 | 1.19 | 1.02 | 0.88 | 0.74 | 0.61 | 0.49 | 0.4 |
| Var 0 | 4.64 | 3.08 | 2.26 | 1.79 | 1.46 | 1.21 | 1.02 | 0.84 | 0.68 | 0.53 | 0.4 |
| Var 1 | 5.62 | 3.98 | 2.85 | 2.22 | 1.8 | 1.47 | 1.21 | 0.97 | 0.77 | 0.58 | 0.4 |
| Var 2 | 6.6 | 4.87 | 3.44 | 2.64 | 2.14 | 1.73 | 1.39 | 1.1 | 0.86 | 0.63 | 0.4 |
Оценить характер тренда показателя отношения приростов для конкретного участка камбиальной зоны относительно выделенных выше пяти вариантов можно следующим расчетно-графическим способом. Используя значения выровненного тренда показателя отношения приростов для каждого годичного кольца по формулам (5–9), следует рассчитать соответствующие предполагаемые значения нормированного возраста для всех пяти вариантов. Затем, преобразуя формулу (4), надо определить отдельно по вариантам для каждого года предполагаемый контрольный возраст, разделив возраст годичного кольца на вычисленный нормированный возраст. В системе координат по осям представлены реальный возраст участка камбиальной зоны и шкала контрольного возраста. Расчетные линии контрольных возрастов для пяти вариантов будут представлены в виде различных линий, сходящихся в одну точку. Для конкретной камбиальной зоны значения такой точки по шкале расчетного контрольного возраста и по шкале реального возраста будут одинаковыми. В случае, когда динамика приростов камбиальной зоны точно соответствуют одному из пяти вариантов, расчетные значения контрольного возраста для такого варианта будут однозначны и представлены в виде прямой линии. Но чаще всего такого совпадения нет. В этом случае искомый контрольный возраст будет соответствовать точке пересечения трендов всех вариантов, однозначной по обеим возрастным шкалам (рис. 5а).
Рис. 5.
(а) Пример результатов расчета контрольного возраста для зоны камбиального прироста по пяти вариантам формул (№ 5–9), описывающих связь нормированного возраста с соотношением ширины поздней и ранней древесины, и его реальное значение.
Цифрами со стрелками обозначено значение контрольного возраста: 1 – реальное значение; 2 – для варианта Var -1; 3 – для варианта Var -2.
По оси абсцисс – возраст камбиальной зоны, годы; по оси ординат – расчетные значения контрольного возраста, годы.
(b) Значение нормированного возраста для жизнеспособных участков камбиальных зон при величине соотношения ширины поздней и ранней древесины менее 0.4: 1 – среднее значение, 2 – отдельные участки камбиальных зон.
По оси абсцисс – значение соотношения ширины поздней и ранней древесины; по оси ординат – нормированный возраст.
Fig. 5. (а) Calculation data example of the cambium increment zone age by five different formulas (No. 5–9) specifying the relationship between the normalized age and the latewood/earlywood ratio as well as its measured value.
The numerals with arrows indicate the control ages: 1 – actual value; 2 – for the line Var -1; 3 – for the line Var -2.
X-axis – cambium zone age, years; Y-axis – calculated control age, years.
(b) Normalized age for the viable cambium portions with the latewood/earlywood width ratio less than 0.4: 1 – average value, 2 – selected cambium portions.
X-axis – latewood/earlywood width ratio; Y-axis – normalized age.

На рис. 5а приведен пример расчета контрольного возраста и номера варианта на основе 5 выделенных вариантов соотношения ширины поздней и ранней древесины. Тренды контрольных возрастов выделенных вариантов не совпадают с реальным значением контрольного возраста, представленного прямой линией на уровне 78 лет. Она расположена между линиями трендов вариантов “Var-1” и “Var-2”. Для более детальной характеристики соотношения ранней и поздней древесины в опытном образце можно использовать номера вариантов в качестве балльной оценки. Прямая линия реальных значений контрольного возраста несколько смещена от линии варианта “Var-1” к варианту “Var-2”, примерно на 0.1 расстояния между ними. Поэтому по балльной шкале распределения вариантов уровень взаимосвязи соотношения ширины поздней и ранней древесины и нормированного возраста для данного участка камбиальной зоны оценивается как равный – 1.1 балла. Следует отметить, что данный способ определения контрольного возраста и номера его варианта за первые 10–20 лет роста участка камбиальной зоны малоинформативен из-за высокой изменчивости его расчетных значений.
Часть деревьев дуба сохраняют жизнеспособность при значениях соотношения ширины поздней и ранней древесины менее 0.4 и нормированного возраста более 1.0. В этом случае взаимосвязь данных показателей приведена на примере анализа 15 участков зон камбиального прироста (рис. 5b). Тренд такой взаимосвязи описывается формулой № 10, где Аstand. – значение нормированного возраста, SI – соотношение ширины поздней и ранней древесины годичного кольца.
Конечное значение трендов нормированного возраста на момент отмирания данных участков камбиальных зон в основном не выше 1.4. При этом продолжительность жизни после достижения контрольного возраста, чаще всего не более 20% от его значения и при максимуме около 70%.
Особенности формирования ранней древесины в процессе жизнедеятельности камбиальной зоны анализировались на основе двух показателей: годичного прироста по радиусу и годичного прироста по площади. В динамике их формирования наблюдается существенное различие. Тренд радиального прироста после начального увеличения, достигнув максимума, затем постепенно понижается. Его форма соответствует типичной возрастной кривой развития, имеющей восходящие и нисходящие ветви (Krenke, 1940). Вершина кривой приходится на период максимального потенциала радиального прироста ранней древесины. Тренд площади поперечного сечения годичного прироста ранней древесины по всему периметру ствола вначале так же закономерно увеличивается до некоторого момента, после которого в различных случаях его величина или продолжает повышаться, как правило, с меньшей интенсивностью, или остается стабильной, или понижается.
Фрагменты линии тренда, в которых наблюдается закономерное изменение его динамики, свидетельствуют о завершении участком камбиальной зоной определенных этапов ее жизнедеятельности и могут быть использованы в качестве реперных зон.
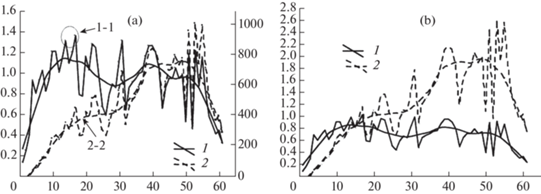
На рис. 6a приведены значения годичного прироста ранней древесины по радиусу и по площади поперечного сечения, взятые на срезе усыхающего дерева дуба на высоте 1.3 м. На графике выделены две реперные зоны. Реперная зона максимального значения радиального годичного прироста (1-1 на рис. 6a) размером в 1.39 мм, определяющая меру ростовых возможностей камбиальной зоны, и реперная зона, фиксирующая окончание закономерного начального увеличения годичного прироста площади ранней древесины (2-2 на рис. 6a) величиной в 390 мм2. Годичный прирост площади ранней древесины в дальнейшем может возрастать, но с меньшей интенсивностью.
Рис. 6.
Пример изменения во времени радиального прироста и площади ранней древесины на срезе усыхающего дерева дуба на высоте 1.3 м.
(а) Годичный прирост ранней древесины, выраженный в натуральных единицах. По оси абсцисс – возраст участка камбиальной зоны, годы. По левой оси ординат: размер годичного радиального прироста ранней древесины, мм; по правой оси ординат: размер площади годичного прироста ранней древесины, мм2; 1 – радиальный прирост ранней древесины, 2 – прирост ранней древесины по площади. 1-1 – реперная зона максимального радиального прироста ранней древесины, 2-2 – реперная зона окончания начального закономерного этапа увеличения прироста ранней древесины по площади.
(b) Годичный прирост ранней древесины, выраженный в относительных величинах. 1 – относительный радиальный прирост ранней древесины, 2 – площадь относительного прироса ранней древесины в квадратных единицах. По оси абсцисс – возраст участка камбиальной зоны, годы. По оси ординат – размер годичного прироста ранней древесины в относительных единицах.
Fig. 6. An example of the change in time of the radial growth and the area of early wood on a section of a drying oak tree at a height of 1.3 m.
(а) Annual growth of early wood, expressed in natural units. The abscissa shows the age of the cambial zone area, years. On the left Y-axis: the size of the annual radial growth of early wood, mm; on the right Y-axis: the size of the area of the annual growth of early wood, mm2; 1 – radial growth of early wood, 2 – growth of early wood by area. 1-1 – reference zone of maximum radial growth of early wood, 2-2 – reference zone of the end of the initial regular stage of increase in growth of early wood over the area.
(b) Annual growth of early wood, expressed in relative terms. 1 – relative radial growth of early wood, 2 – area of relative growth of early wood in square units. The abscissa shows the age of the cambial zone area, years. On the Y-axis – the size of the annual growth of early wood in relative units.
При сравнительной оценке этапов жизнедеятельности участков камбиальных зон в течение всего периода их деятельности абсолютные размеры годичного прироста не являются достаточно релевантными показателями такого процесса, так как в однотипные периоды развития они могут различаться по интенсивности роста. Корректнее использовать относительные значения приростов с учетом ростовых возможностей конкретного участка камбиальной зоны. По этой причине абсолютные значения годичного радиального прироста ранней древесины были переведены нами в относительные величины. Для радиального годичного прироста ранней древесины относительная величина рассчитывалась как частное от деления величины годичного прироста на среднюю из 2–3 самых больших его значений, расположенных в реперной зоне. Для годичного прироста ранней древесины по площади всего периметра относительные величины рассчитывались как частное от деления величины площади каждого годичного прироста на усредненное значение площади годичного прироста в реперной зоне 2-2 на рис. 6a. Пример подобных преобразований показан на рис. 6b, где в относительных величинах выражены значения годичного прироста ранней древесины того же участка камбиальной зоны, что и на рис. 6a.
Реперные зоны ранней древесины могут являться маркерами определенных этапов в жизнедеятельности локального участка камбиальной зоны. Связь реперных зон с соотношением ширины поздней и ранней древесины и нормированным возрастом была исследована на образцах 114 участков камбиальных зон.
На рис. 7a приведено распределение таких зон с соотношением ширины поздней и ранней древесины. Среднее значение показателя отношения приростов для реперной зоны радиального годичного прироста ранней древесины равно 2.44 ± ± 0.066, а для реперной зоны годичного прироста площади ранней древесины 1.87 ± 0.056. Средняя разница значений показателя отношения приростов между данными точками равна 0.6 ± 0.047 и достоверна при уровне значимости Р = 0.001.
Рис. 7.
Распределение 114 реперных зон годичного прироста ранней древесины по радиусу и по площади относительно соотношения ширины поздней и ранней древесины и нормированного возраста.
1 – годичный радиальный прирост ранней древесины, 2 – годичный прирост площади ранней древесины.
(а) Диаграмма распределения реперных зон относительно соотношения ширины поздней и ранней древесины.
(b) Диаграмма распределения реперных зон относительно нормированного возраста.
По оси абсцисс: (а) значение соотношения ширины поздней и ранней древесины в группе; (b) значение нормированного возраста в группе; по оси ординат – количество участков камбиальных зон в группе.
Fig. 7. Distribution of 114 reference zones in respect to the annual earlywood increment in terms of radius/area in relation to the of latewood/earlywood width ratio and normalized age.
1 – annual radial earlywood increment, 2 – annual earlywood area increment.
(а) Reference zones distribution in respect to latewood/earlywood width ratio.
(b) Reference zones distribution in respect to normalized age values.
X-axis: (a) latewood/earlywood width ratio in the group; (b) normalized age per group; Y-axis – number of cambium portions per group.

На рис. 7a приведено распределение таких зон с соотношением ширины поздней и ранней древесины. Среднее значение соотношения ширины поздней и ранней древесины для реперной зоны радиального годичного прироста ранней древесины равно 2.44 ± 0.066, а для реперной зоны годичного прироста площади ранней древесины 1.87 ± 0.056. Средняя разница значений соотношения ширины поздней и ранней древесины между данными точками равна 0.6 ± 0.047 и достоверна при уровне значимости Р = 0.001.
На рис. 7b представлено распределение реперных зон относительно значений нормированного возраста. Средняя величина нормированного возраста для реперной зоны радиального годичного прироста ранней древесины равна 0.24 ± 0.008, а для реперной зоны годичного прироста площади ранней древесины она составляет 0.35 ± 0.009. Средняя разница значений нормированного возраста между данными реперными зонами достоверна и равна 0.11 ± 0.007 и достоверна при Р < 0.001. Полученные данные свидетельствуют о том, что период начального развития и закономерного повышения размеров годичного прироста ранней древесины приходится на первую треть общего времени жизнедеятельности участка камбиальной зоны. При этом после достижения радиальными годичными приростами ранней древесины максимальных значений, площадь годичного кольца ранней древесины в дальнейшем некоторое время продолжает увеличиваться с прежней интенсивностью.
Нормированный возраст позволяет провести сравнительную оценку камбиальных зон по периодам их жизнедеятельности. Период, когда происходит закономерное увеличение прироста ранней древесины по радиусу и по площади, ограничивается значением нормированного возраста равного 0.3 (рис. 7b), затем наступает период постепенного уменьшения радиального прироста ранней древесины, а прирост по площади может быть различным, увеличиваться или уменьшаться (рис. 6). Заключительный период наступает, когда возрастает вероятность отмирания дерева при нормированном возрасте, близком к единице и значению соотношения ширины поздней и ранней древесины, равному 0.4 (рис. 3а).
Особенности взаимосвязи абсолютных размеров радиальных годичных приростов ранней и поздней древесины можно оценить, сравнивая максимальные значения радиального годичного прироста ранней древесины в реперной зоне с вариантами связи соотношения ширины поздней и ранней древесины с нормированным возрастом. На рис. 8а приведены результаты такого сравнения для 106 участков камбиальных зон.
Рис. 8.
Распределение максимальных значений годичного радиального прироста ранней древесины в реперных зонах относительно балльных оценок взаимосвязи соотношения ширины поздней и ранней древесины с нормированным возрастом для 106 участков камбиальных зон (а) и относительно контрольного возраста для 40 участков камбиальных зон (b).
По оси абсцисс – значение годичного радиального прироста ранней древесины в реперной зоне, мм; по оси ординат: (а) Вариант взаимосвязи соотношения ширины поздней и ранней древесины с нормированным возрастом, баллы; (b) значение контрольного возраста, годы.
Fig. 8. Distribution of maximal values of the annual radial earlywood increments in the reference zones in relation to numerical score of relation between the latewood/earlywood width ratio and normalized age for 106 cambium portions (a) and in relation to control age for 40 cambium portions (b).
X-axis – the value of the annual radial earlywood increment in the reference zone, mm; Y-axis: (a) type of relationship between the latewood/earlywood width ratio and normalized age, scores; (b) control age, years.

Достоверной связи между данными показателями не наблюдается. В пределах всего спектра возможных соотношений поздней и ранней древесины, от минимального (вариант “Var-2”) до близкого к максимальному (вариант “Var 2”), в реперной зоне наблюдается формирование различных значений годичного радиального прироста ранней древесины размерами от 0.2 до 1.2 мм. Это значит, что при одинаковой максимальной величине радиального годичного прироста ранней древесины в реперной зоне при нормированном возрасте около 0.25 соотношение поздней древесины к ранней может изменяться примерно в 2.5 раза (табл. 1). Эти результаты согласуются с выводами Косиченко (Kosichenko, 1999) о том, что формирование ранней и поздней древесины у кольцесосудистых растений не находится в тесной взаимосвязи друг с другом и контролируется разными метаболическими системами.
На рис. 8b приведена взаимосвязь размеров годичного прироста ранней древесины в реперной зоне и контрольного возраста для 40 участков камбиальных зон деревьев на высоте ствола 1.3 метра. Существенной связи между данными показателями нет. Коэффициент парной корреляции равен: r = 0.31 при P = 0.054. В пределах диапазона годичного радиального прироста ранней древесины в размере от 0.6 до 1.7 мм величина контрольного возраста изменяется от 50 до 180 лет и выше. Это значит, что потенциальная способность камбиальной зоны формировать низкие или высокие значения радиального годичного прироста ранней древесины не имеет тесной связи со временем ее жизнедеятельности.
Тренд площади годичного прироста ранней древесины может быть одним из признаков, характеризующих состояние камбиальной зоны. Его форма зависит не только от величины радиального прироста ранней древесины, но и от протяженности периметра ствола, который в свою очередь обусловлен как размерами общего годичного прироста в предшествующий период, так и возрастом дерева. Их комбинации могут быть разнообразными. Для понимания некоторых особенностей взаимосвязи динамики годичного прироста ранней древесины по радиусу и по площади в процессе жизнедеятельности камбиальной зоны ниже приведены их тренды, смоделированные в пределах конкретных временных и ростовых параметров.
В моделируемых трех вариантах в масштабе нормированного возраста были заданы значения радиального прироста ранней древесины, выраженные в относительных единицах. При этом увеличение годичного прироста ранней древесины до реперной зоны у них было идентичным, а после нее интенсивность уменьшения прироста в вариантах изменялась в разной степени (рис. 9а).
Рис. 9.
Тренды годичного прироста ранней древесины.
(а) Тренды радиального годичного прироста ранней древесины. 1, 2, 3 – варианты трендов с различной интенсивностью понижения своих значений после реперной зоны.
По оси абсцисс – значение нормированного возраста; по оси ординат – значение годичного радиального прироста ранней древесины в относительных единицах.
(b) Тренды площади годичного прироста ранней древесины, соответствующие ее радиальным значениям в вариантах на рис. 9, А и рассчитанные для двух групп с различным контрольным возрастом: 1, 2, 3 – варианты трендов с контрольным возрастом 128 лет; 4, 5, 6 – варианты трендов с контрольным возрастом 64 года.
По оси абсцисс – возраст годичного кольца, годы; по оси ординат – площадь годичного прироста ранней древесины в относительных квадратных единицах.
Fig. 9. Trends of the annual earlywood increments.
(а) Trends of the radial annual earlywood increment. 1, 2, 3 – different trends with various value decrease rate beyond the reference zone.
X-axis – normalized age; Y-axis – relative annual radial earlywood increment in terms of area.
(b) Trends of the annual earlywood area increments plotted relating to their radial increments specified in the versions shown in Fig. 9, A, calculated for two groups with different control ages: 1, 2, 3 – different trends with control age 128 years; 4, 5, 6 – different trends with control age 64 years.
X-axis – tree-ring age, years; Y-axis – relative annual earlywood increment in terms of area.

Для двух значений контрольного возраста, равных 64 и 128 годам, были рассчитаны шкалы нормированного возраста. На их основе, используя относительные значения радиального годичного прироста ранней древесины, с помощью регрессионного уравнения, характеризующего соотношения поздней и ранней древесины и нормированного времени в варианте “Var 0” (табл. 1), были определены для каждого года относительные значения приростов ранней и поздней древесины, общий годичный прирост и соответствующие значения годичного прироста площади ранней древесины (рис. 9b).
При равных относительных значениях годичного прироста ранней древесины и едином характере ее взаимосвязи с размером поздней древесины в процессе жизненного цикла камбиальной зоны форма тренда годичного прироста площади ранней древесины не зависит от продолжительности жизнедеятельности. В приведенном примере с контрольными возрастами в 64 и 128 лет тренды различаются по абсолютным значениям, но аналогичны по форме.
После реперной зоны динамика прироста ранней древесины по площади в пределах одного варианта соотношения поздней и ранней древесины зависит от интенсивности понижения ее радиального прироста. Стабильное уменьшение радиального прироста обусловлено понижением ростовой активности камбиальной зоны и является определенной характеристикой ее состояния. При этом увеличение или уменьшение годичного прироста по площади ранней древесины может отражать общее состояние проводящей структуры, обеспечивающей объем вертикального тока метаболитов. У дуба формирование зоны ранней древесины играет особо важную роль, так как ее проводящая способность полностью сохраняется только в течение года ее формирования (Zimmermann, 1983).
Процесс увеличения или уменьшения годичного прироста площади ранней древесины при стабильном уменьшении радиального прироста зависит как от динамики общего годичного прироста древесины, так и от размеров периметра ствола. У старых деревьев, при заметном уменьшении радиального прироста ранней древесины, но при большом диаметре ствола, площадь годичного прироста ранней древесины со временем может не только оставаться стабильной, но и увеличиваться. Можно предположить, что сохранение определенного функционального равновесия между приростами ранней древесины по радиусу и по площади есть необходимое условие для поддержания стабильной жизнеспособности деревьев дуба.
II. Ствол дерева
Для изучения особенностей развития участков камбиальных зон, расположенных на разной высоте ствола, были отобраны 12 модельных деревьев, имеющих величину соотношения ширины поздней и ранней древесины равную или близкую к 0.4, позволяющую определить контрольный возраст. Общая характеристика модельных деревьев приведена в табл. 2.
Таблица 2.
Размерные показатели 12 модельных деревьев Table 2. Size parameters of 12 model trees
| Показатели Data | Среднее арифметическое Mean |
Минимум Minimum |
Максимум Maximum |
|---|---|---|---|
| Диаметр ствола на высоте 1.3 метра без коры, см Trunk diameter at a height of 1.3 m without bark, cm |
38 ± 11.0 | 16 | 99 |
| Высота, м Height, m |
24 ± 6.9 | 18 | 33 |
| Возраст, лет Age, years |
111 ± 17.9 | 61 | 230 |
Высоты и диаметры стволов без коры на 1.3 м у модельных деревьев показаны на рис. 10a. Всего на разной высоте ствола модельных деревьев было взято 74 поперечных среза. Соотношение реального, на момент взятия модели, и контрольного возрастов для анализируемых участков камбиальных зон приведено на рис. 10b. Между ними наблюдается определенное различие. Вместе с тем, значения реального и контрольного возраста у отдельно взятых участков камбиальных зон близки между собой. Коэффициент корреляции составляет: ґ = 0.9. Можно предположить, что данные деревья находились на заключительных этапах своего онтогенеза.
Рис. 10.
(а) Диаметр без коры и высота 12 модельных деревьев.
По оси абсцисс – диаметр без коры на высоте 1.3 м, см; по оси ординат – высота дерева, м.
(b) Реальный возраст и контрольный возраст 74 участков камбиальных зон модельных деревьев.
По оси абсцисс – реальный возраст, годы; по оси ординат – контрольный возраст, годы.
Fig. 10. (а) Trunk diameter without bark and the height of 12 model trees.
X-axis – diameter without bark at a height of 1.3 m, cm; Y-axis – tree height (m).
(b) Actual/control age of 74 cambium portions of model trees.
X-axis – actual age, years; Y-axis – control age, years.

Для сравнительного анализа участков камбиальных зон, расположенных на разной высоте по стволу модельного дерева, значения по высоте перевели в относительные нормированные величины – “Нnor.”, рассчитанные как отношение конкретной высоты участка камбиальной зоны к общей высоте дерева.
На рис. 11а представлено распределение значений контрольного возраста участков камбиальных зон по нормированной высоте ствола модельных деревьев.
Рис. 11.
Распределение значений контрольного возраста для 74 участков камбиальных зон по высоте ствола 12 модельных деревьев.
(а) По оси абсцисс – высота дерева, относительные значения; по оси ординат – контрольный возраст, годы.
(b) По оси абсцисс – высота дерева, относительные значения; по оси ординат – контрольный возраст, относительные значения.
Fig. 11. Distribution of the control age values for 74 cambium portions in respect to the trunk height of 12 model trees.
(а) X-axis –relative tree height; Y-axis – control age, years.
(b) X-axis – relative tree height; Y-axis – relative control age.

По стволу дерева от комля к вершине наблюдается тенденция к уменьшению величины контрольного возраста участков камбиальных зон. Поскольку у разных деревьев на одной и той же относительной высоте абсолютные значения контрольного возраста не одинаковы, контрольный возраст участков камбия в пределах ствола дерева также был переведен в относительные нормированные единицы – “Аcon. stand.”, рассчитанные как отношение контрольного возраста конкретного участка камбиальной зоны к контрольному возрасту участка камбиальной зоны на высоте 1.3 м (рис. 11b). Относительная величина контрольного возраста участков камбиальных зон с увеличением высоты по стволу дерева достоверно уменьшается, коэффициент корреляции ґ = –0.85 при Р = 0.05.
Тренд данной зависимости описывается формулой полинома № 11.
(11)
${{А}_{{{\text{con}}{\text{.stand}}{\text{.}}}}} = 1.0221--0.3131{{Н}_{{{\text{nor}}{\text{.}}}}} + 0.5964Н_{{{\text{nor}}{\text{.}}}}^{2}.$Уменьшение величины контрольного возраста локальных участков камбиальных зон по направлению от комля к вершине ствола дерева свидетельствует о закономерном сокращении периода их развития. Эти данные подтверждают вывод, сделанный Кренке (Krenke, 1940) о том, что однотипные метамерные части одного растения обладают различной скоростью старения, так как, имея различный собственный возраст, они в процессе своего развития приходят в конечном итоге к равному функциональному состоянию. Л.А. Лебеденко (Lebedenco, 1955, 1959) указывает, что с увеличением возраста и высоты ствола дуба скорость старения и формирования признаков зрелой древесины в пределах отдельных участков камбиальных зон возрастает.
Согласно теории Кренке (Krenke, 1940) имеющийся на данный момент наличный потенциал жизнеспособности для любой части растения равен исходному общему потенциалу минус использованная его доля за истекший срок жизни. Это положение справедливо как для метамеров, формируемых апикальной первичной меристемой стебля, так и для производных камбия, то есть вторичной латеральной меристемы. Контрольный возраст участка камбиальной зоны в определенной степени является аналогом понятия потенциала жизнеспособности. Его значение по высоте ствола согласно данной теории должно закономерно изменяться. Поэтому следует ожидать, что для ствола в целом соотношение контрольных возрастов камбиальных зон представляет собой некую заданную величину, которая на любой его высоте равна сумме, состоящей из количества лет, прошедших до момента достижения апикальным побегом данной высоты, и значения контрольного возраста участка камбиальной зоны, сформированной на данной высоте. В этом случае контрольный возраст участка камбиальной зоны в нижней части ствола наиболее полно характеризует общий контрольный возраст дерева. Для сравнительного анализа динамики контрольного возраста по высоте у различных стволов их значения были преобразованы в относительные величины, которые были получены как частное от деления суммы значений контрольного возраста конкретного участка камбиальной зоны на определенной высоте и предшествующего ей возраста ствола на контрольный возраст участка камбиальной зоны из нижней части ствола (формула 12). В случае если проверяемая гипотеза справедлива, то частное от такого деления должно быть близко к единице. Обозначим данное значение как комплексный возраст:
где F – расчетный комплексный возраст соответствующей определенному (n) участку камбиальной зоны ствола; Аcon.n – контрольный возраст участка камбиальной зоны на данной (n) высоте ствола; Аn – возраст ствола до высоты данной (n) зоны прироста; Аcon.1.3 – контрольный возраст участка камбиальной зоны на высоте 1.3 м.На рис. 12a приведено распределение значений комплексного возраста участков камбиальных зон по относительной высоте ствола у 12 модельных деревьев.
Рис. 12.
(а) Значение расчетного комплексного возраста для участков камбиальных зон на различной высоте ствола у 12 модельных деревьев.
(b) Значения вариантов связи соотношения ширины поздней и ранней древесины, выраженные в баллах, с нормированным возрастом для участков камбиальных зон на различной высоте ствола 12 модельных деревьев.
По оси абсцисс – высота модельного дерева, относительные значения; по оси ординат: (а) значение расчетного комплексного возраста участка камбиальной зоны; (b) значение баллов для вариантов связи соотношения ширины поздней и ранней древесины и нормированного возраста.
Fig. 12. (а) Combined tree age estimation for cambium portions located at different trunk height of 12 model trees.
(b) Types of relationship between latewood/earlywood width ratio specified as scores and normalized age for cambium portions of 12 model trees.
X-axis – relative height of model tree; Y-axis: (a) estimated combined age of cambial portions; (b) score number for the types of relationship between the latewood/earlywood width ratio and the normalized age.

Количество участков камбиальных зон, расположенных выше 1.3 м, равно 56. Среднее для них значение комплексного возраста равно: 0.99 ± ± 0.018 при лимитах 0.6–1.4. Оно близко к гипотетической величине, равной 1.0. Фактическая величина нормированного отклонения составляет: t = 0.55. Данное значение меньше стандартного нормированного отклонения равного: t = 1.96 при P = 0.05, что указывает на принадлежность анализируемых данных к одной генеральной совокупности в пределах нормального распределения (Lakin, 1973). Этот результат не отвергает выдвинутую гипотезу о наличии некоторого общего возрастного максимума, когда различные по высоте ствола участки камбиальных зон одновременно достигают контрольного возраста. Он согласуется с выводом Кренке (Krenke, 1940) о зависимости потенциала жизнеспособности отдельных метамеров от времени их формирования по отношению к общему периоду жизни растения. В этом плане контрольный возраст в нижней части ствола можно рассматривать как показатель, характеризующий возрастное состояние всего ствола в целом.
На рис. 12b представлено распределение балльных значений, характеризующих варианты связи соотношения ширины поздней и ранней древесины с нормированным возрастом, для участков камбиальных зон, расположенных по относительной высоте стволов модельных деревьев. Как видно, в стволе дерева могут формироваться камбиальные зоны, существенно различающиеся по значениям вариантов связи соотношения ширины поздней и ранней древесины в годичном кольце, включая все возможные варианты от “Var-2” до “Var 2”.
Результаты анализа, приведенные на рис. 11а, 11b и 12а, 12b, указывают на то, что макроструктура годичных колец ствола дуба черешчатого в течение его онтогенеза формируется под влиянием двух факторов. С одной стороны, участки камбиальных зон на разной высоте ствола относительно синхронно завершают этапы своего развития, что предполагает закономерное уменьшение со временем доли поздней древесины (рис. 11а, 11b). С другой стороны, соотношение поздней и ранней древесины в разных частях ствола может быть различным.
Ниже приведены параметры годичных радиальных приростов, сформированных камбиальными зонами в нижней, средней и верхней части ствола у 4 моделей (табл. 3).
Таблица 3.
Характеристика модельных деревьев Table 3. Characteristics of model trees
| Таксационные параметры ствола Taxation parameters of a trunk |
Характеристики камбиальной зоны Characteristics of a cambial zone |
|||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| 1* | 2* | 3* | 4* | 5* | 6* | 7* | 8* | 9* | 10* | 11* |
| 1 | 32 | 72 | Живое Alive | 231 | 1.3 | 226 | 36.0 | 180 | 1.20 | 1.0 |
| 13 | 204 | 25.7 | 150 | 0.86 | –1.0 | |||||
| 25 | 157 | 13.5 | 150 | 0.50 | –1.0 | |||||
| 2 | 33 | 100 | Усохшее Dried |
185 | 1.3 | 180 | 49.3 | 235 | 1.70 | –0.2 |
| 17 | 141 | 20.7 | 100 | 1.15 | –1.0 | |||||
| 27 | 101 | 5.0 | 84 | 0.47 | –2.0 | |||||
| 3 | 19 | 27 | Живое Alive |
107 | 1.3 | 103 | 13.5 | 145 | 0.70 | –0.8 |
| 8 | 89 | 10.3 | 150 | 0.75 | –0.8 | |||||
| 15 | 57 | 3.3 | 63 | 0.36 | –1.0 | |||||
| 4 | 18 | 16 | Усохшее Dried |
74 | 1.3 | 69 | 8.1 | 64 | 0.22 | 2.0 |
| 7 | 55 | 7.0 | 39 | 0.18 | 1.9 | |||||
| 16 | 27 | 2.3 | 16 | 0.14 | 0.0 | |||||
Примечание. 1* – № модели; 2* – высота, м; 3* – диаметр на 1.3 м без коры, см; 4* – состояние дерева; 5* – возраст дерева, лет; 6* – высота участка камбиальной зоны, м; 7* – возраст участка камбиальной зоны, лет; 8* – радиус ствола без коры, см; 9* – контрольный возраст, лет; 10* – максимальное значение радиального прироста ранней древесины, мм; 11* – варианты отношения нормированного возраста и соотношения ширины поздней и ранней древесины, баллы.
Note. 1* – № of model; 2* – height, m; 3* – diameter at 1.3 m without bark, cm; 4* – vital state of tree; 5* – age of tree, years; 6* – height of the cambial zone portion, m; 7* – age of the cambial zone portion, years; 8* – trunk radius without bark, cm; 9* – control age, years; 10* – maximum value of the earlywood radial growth, mm; 11* – versions of relationship between the normalized age and the latewood / earlywood width ratio, points.
Между локальными камбиальными зонами в пределах одного дерева на разной высоте ствола наблюдаются различия по следующим показателям: контрольный возраст, максимальный размер годичного радиального прироста ранней древесины в реперной зоне, вариант взаимосвязи соотношения ширины поздней и ранней древесины с нормированным возрастом. Контрольный возраст участков камбиальных зон ствола на высоте 1.3 м варьирует от 64 до 235 лет, что указывает на существенные различия в продолжительности онтогенеза модельных деревьев.
На рис. 13 представлены данные по соотношению нормированного возраста участков камбиальных зон на разной высоте стволов 4 модельных деревьев в процессе их роста.
Рис. 13.
Изменение нормированного возраста участков камбиальных зон, расположенных на разной высоте ствола, в течение жизни дерева.
М – 1, 2, 3, 4 – номер модели. 1, 2, 3 – высота расположения участка камбиальной зоны по стволу. 1 – 1.3 м; 2 и 3 соответственно: М1 – 13 и 25 м; М2 – 17 и 27 м; М3 – 8 и 15 м; М4 – 7 и 16 м.
По оси абсцисс – возраст дерева, год; по оси ординат – значение нормированного возраста.
Fig. 13. Normalized age of cambium portions located at different trunk height through the tree life.
M – 1, 2, 3, 4 – model number. 1, 2, 3 – location of the cambium portion at the trunk height. 1 – 1.3 m; 2 and 3, respectively: M1 – 13 and 25 m; M2 – 17 and 27 m; M3 – 8 and 15 m; M4 – 7 and 16 m.
X-axis – tree age, years; Y-axis – normalized age.

Характер изменения нормированного возраста участков камбиальных зон по стволу дерева имеет ряд общих закономерностей. В большинстве случаев их значения со временем сближаются и в графическом изображении даже пересекаются. Это значит, что до пересечения нормированного возраста вышерасположенные по стволу участки камбиальных зон находятся на более ранних этапах развития, а после пересечения они выравниваются или становятся в стадийном отношении более старыми. Время таких пересечений зависит от индивидуальных особенностей дерева. На тот факт, что у молодых деревьев вышерасположенные участки камбиальных зон по развитию являются более молодыми, чем нижние, указывает ряд авторов (Wimmer, 1994; Zobel, Jett; 1995).
Нормированный возраст участка камбиальной зоны позволяет выделить три периода ее функционального состояния. До величины нормированного возраста равного 0.2–0.3 наблюдается потенциальное увеличение размеров радиального прироста ранней древесины и, соответственно, период активного роста. Далее, до значения 0.8 – время активной жизнедеятельности, но одновременно это и период старения. У большинства отмерших деревьев значение конечного нормированного возраста участков камбиальных зон находится в пределах: 0.8–1.4. Этот диапазон может служить ориентиром при оценке возрастного состояния деревьев дуба черешчатого с повышенной вероятностью отмирания. Интенсивность развития деревьев в онтогенезе может быть различной. Дерево дуба может достичь контрольного возраста, и, соответственно, нормированного возраста равного 1, характеризующих период возможного скорого отмирания, как в возрасте около 200 лет (рис. 13, М1), так и в возрасте около 70 лет (рис. 13, М4).
Ствол дерева представляет собой целостную функциональную систему. Вместе с тем, в различные периоды жизни отдельные участки в комлевой и вершинной его части могут существенно различаться по своему состоянию. На графиках рис. 13 в позиции М1 и М3 представлены живые деревья, а в позиции М2 и М4 усохшие. У усохших деревьев в последние годы все участки камбиальных зон ствола по значению нормированного возраста были близки или выше единицы, особенно в верхней части. Вероятнее всего эти деревья испытывали ослабление по вершинному типу. Дерево М1 живое, но камбиальные зоны имеют нормированный возраст 1 и выше. Вероятно, дерево находилось в критическом состоянии. Дерево М3 более благополучно, но нормированный возраст вершины приближается к 1, что предполагает начало процессов значительного ослабления.
Таким образом, сравнительный анализ нормированного возраста участков камбиальных зон по высоте ствола позволяет дать общую оценку этапов развития дерева дуба в различные периоды его жизни.
Рассматривая процессы формирования ранней и поздней древесины в стволе дуба черешчатого в качестве критерия состояния, следует отметить следующие на первый взгляд противоречивые связи между характером прироста и особенностями роста дерева. С одной стороны, в лучших условиях роста деревья дуба формируют больший процент поздней древесины (Korovin, Oganesyants, 2007), а в процессе старения или ослабления дерева доля поздней древесины уменьшается. Вместе с тем, как показано выше, деревья изначально могут заметно различаться по данному признаку (табл. 1 и 3).
Наблюдается прямая зависимость между размером поздней древесины предшествующего года и размером ранней древесины последующего года (Belov, 1987; Tikhomirov, 1989). Это связано с тем, что условия роста предшествующего года во многом определяют интенсивность формирования поздней, а в следующем году и ранней древесины. Прослеживается прямая связь между содержанием крахмала в заболони осенью предыдущего года и приростом ранней древесины последующего года (Rubtsov, Utkina, 2008). Вместе с тем, не прослеживается четкой взаимосвязи между максимальными размерами радиального годичного прироста ранней древесины в реперной зоне и такими признаками, как доли поздней древесины в годичном кольце (рис. 8а) и контрольным возрастом (рис. 8b).
Вопрос заключается в том, насколько величина годичного прироста и доля поздней древесины могут служить показателем состояния дуба? С одной стороны, их уменьшение наблюдается у ослабленных и стареющих деревьев, вместе с тем, в природе существуют нормально развивающиеся деревья, значительно различающиеся по данным признакам. Для одних деревьев одинаковые значения признака служат показателями их ослабления, для других они являются нормой.
Данные противоречия можно устранить, если предположить в качестве гипотезы, что в начальный период жизнедеятельности участка камбиальной зоны формируется определенный тип нормы реакции генотипа с характерным для нее уровнем метаболической активности, определяющий затем величину соотношения размеров поздней и ранней древесины. Под нормой реакции генотипа понимается наследственно обусловленная амплитуда возможных изменений в реализации генотипа, которая определяет число и характер возможных вариантов фенотипа, или модификаций, при различных условиях внешней среды (Dudka et al., 1984). Как отмечал П.Б. Раскатов (Raskatov, 1979), норма реакции имеет приспособительный характер к воздействию различных факторов. Выделенные варианты взаимосвязи показателя отношения приростов с нормированным возрастом указывают на существенное различие камбиальных зон по данному признаку в течение всего периода их жизнедеятельности (рис. 4а), которое может наблюдаться как между деревьями, так и в пределах одного ствола (рис. 12b).
Существенное различие в соотношении поздней и ранней древесины в годичных кольцах наблюдается и на разной высоте одного ствола (рис. 12b). Условия деятельности камбиальных зон в стволе могут зависеть от состояния ветвей кроны. В литературе имеются данные, указывающие, что между суммой площадей сечения ветвей первого порядка и массой фракции крон, включая скелет и листья, существует очень тесная связь (Gul’be et al., 1988). У деревьев дуба черешчатого с более развитой кроной формируется больший радиальный размер ранней и поздней древесины, чем у деревьев с менее развитой кроной (Kaplina, 2019).
Формирование древесины в годичном кольце происходит под воздействием различных стимуляторов роста. Важнейшими из них являются гормоны, играющие определенную роль в формировании годичных колец в зависимости от зоны развития, в которой они действуют. Градиент распределения по стволу ауксина и гиббереллина во многом определяет структуру годичного кольца (Menailo, 1987; Buttò et al., 2020). Как указывает R. Aloni (2007), ауксин является основным гормональным сигналом, контролирующим образование древесины. Он в основном производится в молодых побегах, движется вниз через камбий и стимулирует формирование древесины. Уменьшение концентрации ауксина вызывает изменения состава и плотности древесины. Молодая древесина на деревьях индуцируется молодыми листьями, в то время как взрослая древесина может образовываться дальше от этих листьев. Цитокинин от корней движется вверх, увеличивает чувствительность камбия к сигналу ауксина и стимулирует деление камбиальных клеток. Гиббереллин способствует удлинению побегов и образованию длинных волокон и трахеид. Центробежное движение этилена из дифференцирующихся клеток ксилемы наружу к коре индуцирует радиальные сосудистые лучи. Johnsson C. et al. (2019) пришли к выводу, что образование древесины регулируется интегрированным сочетанием как ауксина, так и гиббереллиновой кислоты.
При этом выдвинуто мнение, что переход от ювенильной древесины к зрелой ее форме контролируется уменьшающимся вниз градиентом ауксина. Повышенная концентрация ауксина увеличивает активность камбиальной зоны и интенсивность формирования ксилемы (Uggla, Mellerowitcz et al., 1998). Количество сахара, его доступность в зоне камбия влияют на формирование структуры древесины. Резкие градиенты концентрации сахаров предполагают роль передачи сигналов в процессе формирования структуры сосудов у сосны и образованию ранней и поздней древесины (Uggla et al., 2001). Моделирование позволяет лучше понять роль сахаров в процессе формирования древесины. По данным Cartenì F. et al. (2018) модель достоверно воспроизводит анатомические особенности и кинетику трахеид четырех видов хвойных. В начале вегетационного периода низкая доступность сахара в камбии приводит к медленному отложению стенки, что позволяет растянуть время увеличения, таким образом, получаются большие клетки с тонкими стенками (т.е. ранняя древесина). В конце лета и в начале осени из-за высокой доступности сахара образуются более узкие клетки с толстыми клеточными стенками (т.е. поздняя древесина).
В этой связи логично сделать предположение, что состояние ветвей, степень их облиственности определяет количество ауксина, поступающего в ствол дерева, и оказывает влияние на процесс формирования ранней и поздней древесины и их соотношение. Имеются данные о генетическом контроле формирования древесины. Результаты изучения генома древесных растений подтверждают роль генов на разных этапах формирования древесины и на взаимосвязанное функционирование различных меристем (Schrader, 2003; Groover, Robischon, 2006). Выделены два гена MADS-бокса, которые регулируют активность сосудистого камбия и вторичный рост, модулируя гомеостаз ауксина у рода Populus (Zhu et al., 2018).
Обобщая данные литературы о формировании ранней и поздней древесины в стволе дерева, можно сделать вывод, что этот процесс является следствием взаимосвязи многих внутренних факторов, таких как продуцирование, взаимосвязь и концентрация различных гормонов и углеводов, состояния дерева и его ветвей, генетического контроля. Представленные в статье данные указывают на регулирующую роль этих процессов со стороны целостной системы дерева, как в пределах ствола, так и во времени. С увеличением высоты ствола дуба формирование ранней и поздней древесины в процессе прохождения возрастных стадий от ювенильной к зрелой и затем стареющей ускоряется. В результате к моменту глубокой старости ствола по всей его высоте камбиальные зоны находятся в примерно одинаковом состоянии завершения своего возрастного развития.
ВЫВОДЫ
В течение жизнедеятельности участка камбиальной зоны отношение радиальных приростов поздней древесины к ранней уменьшается, стремясь к нулю. Предложено понятие контрольного возраста, при котором для дуба черешчатого соотношение средних значений радиального годичного прироста поздней древесины к ранней древесине равно 0.4, что соответствует состоянию деревьев с повышенной вероятностью его отмирания.
Нормированный возраст, определяемый как отношение возраста годичного кольца к контрольному возрасту, позволяет дать различным камбиальным зонам сравнительную оценку в относительных величинах по возрастным периодам их жизнедеятельности и старения. В онтогенезе он изменяется от 0.1 до 1.0 и выше. До значения равного 0.3 наблюдается закономерное увеличение прироста ранней древесины по радиусу и по площади. Далее радиальный прирост ранней древесины постепенно уменьшается, а прирост по площади может быть различным, увеличиваться или уменьшаться. При значении нормированного возраста близкого к единице высока вероятность отмирания дерева.
Камбиальные зоны при одинаковом нормированном возрасте могут существенно различаться по величине отношения приростов. Для оценки подобного различия в пределах возможного спектра изменений было выделено пять равномерно отдаленных трендов, описывающих варианты связи отношения приростов и нормированного возраста за период от 0.1 до 1.0. Этим трендам присвоены балльные значения, позволяющие более детально характеризовать промежуточные варианты. Приведена методика расчета указанных трендов.
Не наблюдается тесной взаимосвязи между продолжительностью жизни участка камбиальной зоны и максимальной величиной радиального годичного прироста ранней древесины в период ее наиболее интенсивного роста.
В начальный период жизнедеятельности камбиальных зон, при нормированном возрасте в пределах его значений 0.1–0.3, происходит увеличение радиального прироста ранней древесины до некоторого максимума, после которого наблюдается уменьшение ее прироста. При этом характер формирования годичного прироста площади ранней древесины, ее увеличение или уменьшение, зависит от интенсивности понижения радиального прироста ранней древесины.
В стволе дерева по направлению от комля к вершине ускоряются процессы старения локальных участков камбиальных зон, так как, различаясь по календарному возрасту, они примерно одновременно достигают контрольного возраста. Этот факт указывает на существование общего механизма, регулирующего жизнедеятельность участков камбиальных зон ствола дерева в процессе его онтогенеза. Контрольный возраст участков камбиальных зон, расположенных в нижней части ствола, более полно отражает общий период развития дерева.
На разной высоте ствола в однотипные периоды развития участков камбиальных зон средние значения соотношения годичного прироста поздней и ранней древесины могут быть различными.
Деревья дуба черешчатого в процессе онтогенеза могут существенно различаться по скорости своего развития. Критерием такого процесса служат значения нормированного возраста для участков камбиальных зон на различной высоте ствола.
Список литературы
Aloni R. 2007. Phytohormonal mechanisms that control wood quality formation in young and mature trees. – In: The compromised wood workshop. P. 1–22.
[Belov] Белов А.Н. 1987. Соотношение прироста ранней и поздней древесины в очагах массового размножения насекомых фитофагов. – Изв. ТСХА. 4: 192–201.
Bigler C.J. 2003. Growth-dependent tree mortality: ecological processes and modeling approaches based on tree-ring data: A dissertation submitted to the Swiss Federal Institute of Technology Zurich for the degree of Doctor of Sciences, co-examiner. Diss. ETH No. 15145. 131 p.
Bigler C., Bugmann H. 2004. Predicting the time of tree death using dendrochronological data. – Ecol. Appl. 14: 902–914.
Botkin D.B., Janak J.F., Wallis J.R. 1972. Some ecological consequences of a computer model of forest growth. – J. Ecol. 60: 849–872.
Buttò V., Deslauriers A., Rossi S., Morin H., Rozenberg P., Shishov V. 2020. The role of plant hormones in tree-ring formation. – Trees – structure and function. 34 (2): 315–335.
Cartenì F., Deslauriers A., Rossi S., Morin H. 2018. The Physiological Mechanisms Behind the Earlywood-To-Latewood Transition: A Process-Based Modeling Approach. – Frontiers in Plant Science. 9:1053: 1–12.
[Chailakhyan] Чайлахян М.Х. 1980. Целостность организма в растительном мире. – Физиология растений. 27 (5): 917–941.
[Chevedaev] Чеведаев А.А. 1963. Дуб, его свойства и значение. М. 234 с.
Dobbertin M. 2005. Tree growth as indicator of tree vitality and of tree reaction to environmental stress: a review. – European Journal of Forest Research. 124 (4): 319–333.
[Dudka] Дудка И.А., Вассер С.П., Голубинский И.Н., Шеляг-Сосонко Ю.Р., Блюм О.Б., Зиман С.Н., Мусатенко Л.И., Брайон А.В., Лагутина А.В. 1984. Словарь ботанических терминов. Киев. 308 с.
[Elagin] Елагин И.Н. 1962. Методика изучения процессов формирования годичного кольца у дуба. – В кн.: Физиология древесных растений. М. С. 262–275.
[Ezau] Эзау К. 1980. Анатомия семенных растений. Кн. 1. М. 224 с.
Groover A., Robischon M. 2006. Developmental mechanisms regulating secondary growth in woody plants. – Curr. Opin. Plant Biol. 9 (1): 55–58.
[Grudzinskaya] Грудзинская И.А. 1962. Зависимость строения годичного кольца древесины от развития побегов у дуба. – В кн.: Физиология древесных растений. М. С. 246–261.
[Gul’be] Гульбе Т.А., Рождественский С.Г., Уткин А.И., Каплина Н.Ф., Гульбе Я.И., Ермолова Л.С. 1988. Связь между массой кроны и суммой площадей сечения ветвей первого порядка у деревьев разных пород. – В кн.: Анализ продуктивной структуры древостоев. М. С. 28–48.
Johnsson C., Jin X., Xue W., Dubreuil C., Lezhneva L., Fischer U. 2019. The plant hormone auxin directs timing of xylem development by inhibition of secondary cell wall deposition through repression of secondary wall NAC-domain transcription factors. – Physiol. Plant. 165 (4): 673–689.
[Kaplina] Каплина Н.Ф. 2019. Влияние развития кроны на радиальный прирост ранней и поздней древесины ствола дуба черешчатого. – В кн.: Вестник Поволжского гос. технологического ун-та. Серия: лес, экология, природопользование. 2 (42): 17–25
[Korovin, Oganesyants] Коровин В.В., Оганесянц Л.А. 2007. Дуб в лесоводстве и виноделии. М. 480 с.
[Kosichenko] Косиченко Н.Е. 1999. Влияние генотипа – среды на формирование микроструктуры стебля и диагностика технических свойств, роста и устойчивости древесных растений: Автореф. дис. … докт. биол. наук. Воронеж. 40 с.
[Krenke] Кренке Н.П. 1940. Теория циклического старения и омоложения растений и практическое ее применение. М. 136 с.
[Lakin] Лакин Г.Ф. 1973. Биометрия. М. 343 с.
[Lebedenco] Лебеденко Л.А. 1955. Онтогенез древесины корней и стволов некоторых представителей порядка букоцветных: Автореф. дис. … канд. биол. наук. М. 21 с.
[Lebedenco] Лебеденко Л.А. 1959. Формирование древесины корней и ствола у дуба восточного (Quercus macranthera F. et M.). – Биол. науки. Науч. доклады высшей школы. 2: 131–162.
Leprovost G., Stokes A. 2001. Wood Formation in Trees. – Plant Physiol. 127: 1513–1523.
[Lovelius] Ловелиус Н.В. 1979. Изменчивость прироста деревьев. Л. 232 с.
[Menailo] Менайло Л.Т. 1987. Гормональная регуляция ксилогенеза хвойных. Новосибирск. 184 с.
[Mikhailov] Михайлов М.М. 1959. Физико-механические свойства древесины дуба Чувашской АССР у деревьев различного возраста. – Науч. докл. высш. школы. Лесоинженерное дело. 1: 160–163.
[Palandzhyan] Паланджян В.А. 1989. Структурно-функциональные изменения древесины в онтогенезе растения. – Биологич. журн. Армении. 42 (3): 181–186.
[Raskatov] Раскатов П.Б. 1950. О зависимости структуры годичных колец древесины от онтогенеза образовательных тканей. – Докл. АН СССР. Новая серия. 71 (4): 749–751.
[Raskatov] Раскатов П.Б. 1956. Анатомические исследования древесины в целях познания закономерностей онтогенеза дерева. – В сб.: Науч. зап. Воронежского лесотехнического ин-та. Воронеж. 15. С. 163–176.
[Raskatov] Раскатов П.Б. 1979. Экологическая анатомия вегетативных органов деревьев и кустарников. Воронеж. 181 с.
Rohner B. 2012. Growth and mortality of oak (Quercus spp.): a combined analysis of monitoring and tree-ring data from Swiss forest reserves: A dissertation submitted to ETH ZURICH for the degree of Doctor of Sciences presented by DISS. ETH NO. 20595. 145 p.
[Rubtsov, Utkina] Рубцов В.В., Уткина И.А. 2008. Адаптационные реакции дуба на дефолиацию. М. 302 с.
[Rusalenko] Русаленко А.И. 1986. Годичный прирост деревьев и влагообеспеченность. Минск. 238 с.
[Samtsov] Самцов А.С. 1966. Периодичность роста и формирование структуры древесины рано и поздно распускающихся форм дуба черешчатого в БССР: Автореф. дис. … канд. биол. наук. Минск. 22 с.
[Savina] Савина А.В. 1941. Влияние рубок ухода на строение древесины дуба. – В сб.: Физиологические исследования древесных пород. ВНИИЛХ. Вып. 21. Пушкино. С. 137–144.
[Savina] Савина А.В. 1956. Физиологическое обоснование рубок ухода. М.– Л. 74 с.
Schrader. J. 2003. Developmental Biology of Wood Formation – Finding Regulatory Factors Through Functional Genomics: Diss. … Doct. Sci. 60 p. ISSN. 1401–6230. ISBN 91-576-6526-5
[Shiyatov] Шиятов С.Г. 1973. Дендрохронологическая шкала кедра сибирского на северной границе его произрастания в долине р. Таз. – Лесоведение. 4: 40–45.
[Shiyatov] Шиятов С.Г. 1986. Дендрохронология верхней границы леса на Урале. М. 136 с.
[Shirnin] Ширнин В.К. 1986. Селекция дуба черешчатого на качество древесины. – В сб: науч. тр. Селекция и семеноводство дуба черешчатого. Воронеж. С. 40–52.
StatSoft Statistica 7.0.61.0 En. 2006. Platform: Windows.
[Tikhomirov] Тихомиров А.В. 1989. Структура радиального годичного прироста как показатель состояния дуба черешчатого. – В кн.: Состояние дубрав лесостепи. М. С. 77–97.
Uggla C., Magel E., Moritz T., Sundberg B. 2001. Function and dynamics of auxin and carbohydrates during earlywood/latewood transition in Scots pine. – Plant Physiol. 125: 2029–2039.
Uggla C., Mellerowitcz E., Sunberg B. 1998. Indole-3-acetic acid controls cambial growth in Scots pine by positional signaling. – Plant Physiol. 117: 113–121.
[Vakin] Вакин А.Т. 1954. Фитопатологическое состояние дубрав Теллермановского леса. – Тр. инст. леса АН СССР. 16: 50–109.
[Vikhrov] Вихров В.Е. 1953. Строение и физико-механические свойства ранней и поздней древесины дуба. – Тр. инст. леса АН СССР. 9: 29–38.
[Vikhrov] Вихров В.Е. 1954. Строение и физико-механические свойства древесины дуба. М. 264 с.
Wimmer R. 1994. Structural, chemical and mechanical trends within coniferous trees. – In: Modeling tree-ring development cell structure and environment. Freiburg. P. 2–11.
[Yakovlev] Яковлев Ф.С. 1949. Фитоценологическое и эколого-анатомическое изучение сухостоя дуба. – Тр. Воронежского заповедника. Вып. 3. М. С. 34–69.
[Yatsenko-Khmelewski] Яценко-Хмелевский А.А., Кобак К.И. 1978. Анатомическое строение древесины основных лесообразующих пород СССР. Л. 64 с.
[Zavada] Завада Н.Д. 1983. Дендрохронологический анализ прироста в связи с кумулятивным эффектом дефолиации насаждения. – В кн.: Пути повышения продуктивности лесов Украины и Молдавии. Киев. С. 83–86.
Zhu Y., Song D., Xu P. 2018. A HD-ZIP III gene, PtrHB4, is required for interfascicular cambium development in Populus. – Plant Biotechnol. J. 16 (3): 808–817.
Zimmermann M.H. 1983. Xylem structure and ascent of sap. Springer-Verlag. Berlin, Heidelberg, New York, Tokyo. 143 p.
Zobel B.J., Jett J.B. 1995. Genetics of wood production. Berlin. 337 p.
Zobel B.J., Sprague J.R. 1998. Juvenile wood in forest trees. Berlin. 300 p.
Дополнительные материалы отсутствуют.
Инструменты
Ботанический журнал