

Ботанический журнал, 2022, T. 107, № 6, стр. 544-560
АНАЛИЗ ФУНКЦИОНАЛЬНЫХ ПРИЗНАКОВ И СТРУКТУРА ИХ СВЯЗЕЙ В ЦЕНОПОПУЛЯЦИЯХ PANZERINA LANATA (LAMIACEAE)
А. Ю. Асташенков 1, 2, **, В. Н. Годин 3, ****, В. А. Черемушкина 1, *, Е. Б. Таловская 1, 2, ***
1 Центральный сибирский ботанический сад СО РАН
630090 г. Новосибирск, ул. Золотодолинская 101, Россия
2 Новосибирский национальный исследовательский государственный университет
630090 г. Новосибирск, ул. Пирогова 2, Россия
3 Московский педагогический государственный университет
119991 Москва, Малая Пироговская ул., 1, стр. 1, Россия
** E-mail: astal@bk.ru
**** E-mail: vn.godin@mpgu.su
* E-mail: cher.51@mail.ru
*** E-mail: kolegova_e@mail.ru
Поступила в редакцию 01.06.2021
После доработки 26.03.2022
Принята к публикации 29.03.2022
- EDN: VSGXAZ
- DOI: 10.31857/S0006813622060047
Аннотация
Изучена взаимосвязь 10 функциональных признаков у Panzerina lanata в ценопопуляциях, расположенных в различных местообитаниях Сибири (Тувы и Горного Алтая). Установлено, что исследованные признаки более вариабельны, но слабо скоррелированы в ненарушенных песчаных и каменистых степях, чем в нарушенной песчаной степи. Выявлено, что в ненарушенных вариантах песчаных и каменистых степей P. lanata развивает мощные особи с большей разветвленностью побеговой системы и высокой семенной продуктивностью. В нарушенной степи в процессе зарастания залежи P. lanata формирует небольшие, слабоветвящиеся особи с низкой семенной продуктивностью. Анализ общей и согласованной изменчивости признаков показал, что лучший индикаторный функциональный признак – потенциальная семенная продуктивность, значение которого определяет способность к самоподдержанию ценопопуляций в различных местообитаниях. Высокая его детерминированность и скоординированность с другими признаками, на примере P. lanata, дает возможность использовать его для анализа устойчивости популяций многолетних стержнекорневых растений. Онтогенетическая структура ценопопуляций в ненарушенных местообитаниях отражает результат сбалансированного оборота поколений, выровненные темпы развития особей и периодичную инспермацию, что ведет к волновым процессам и формированию многовершинного спектра. В нарушенном степном фитоценозе отмечается асинхронный темп развития особей и несбалансированный оборот поколений, низкая и нерегулярная инспермация, что обуславливает формирование одновершинного левостороннего спектра.
Популяции растений исследуются в различных аспектах: фитоценологами – как структурные и функциональные единицы ценоза (Smirnova, 1987; Smirnova et al., 1993; Zaugol’nova, 1994; Vostochnoevropejskie…, 2004; Onipchenko, 2014), эволюционистами и типологами – как структурные единицы эволюционного процесса (Stebbins, 1974; Grant, 1984; Kurchenko, 1985, 2010). В настоящее время в фитоценологии оценку устойчивости популяций растений дают с помощью функциональных признаков (Römermann et al., 2016; Wood et al., 2017). Функциональные признаки – это морфологические, (эколого-) физиологические, а также репродуктивные признаки растений, которые влияют на рост, воспроизводство и выживание особи и, таким образом, не только косвенно, но и напрямую влияют на приспособленность растений (Cornelissen et al., 2003; Karbstein et al., 2020). Полученные из прямых измерений или баз данных они используются для изучения реакции популяций, видов, сообществ и экосистем на окружающую среду (Bernhardt-Römermann et al., 2011).
Анализ функциональных признаков привел к настоящему прорыву в области функциональной экологии. По этой теме накоплен большой фактических материал и созданы глобальные базы данных (Cornelissen et al., 2003; Pérez-Harguindeguy et al., 2013; Klimešová et al., 2019; Wigley et al., 2020). Полученные сведения являются важными показателями пластичности видов и размера экологической ниши, что дает ценную информацию о процессах, структурирующих растительные сообщества (Gross et al., 2013; Dudova et al., 2019; Karbstein et al., 2020; Onipchenko et al., 2020) и популяции отдельных таксонов (Zlobin et al., 2013; Bjorkman et al., 2018; Wigley et. al., 2020; Adler et. al 2014). Также анализ корреляций между функциональными признаками различных видов и климатическими условиями дает возможность более глубокого понимания общих функциональных взаимосвязей и эволюционных процессов у видов разных жизненных форм (деревьев, полудревесных, трав) в разнообразных биомах и климатических регионах (Reich et al., 1997; Rostova, 1999; 2000; Ferraro et al., 2021; Xi et al., 2021). Тем не менее, многие аспекты в функциональной экологии все еще остаются мало изученными. Среди них исследование внутри- и межпопуляционных взаимосвязей функциональных признаков и их роль в устойчивом развитии популяций растений (González de Andrés et al., 2021). Популяционно-онтогенетический подход, включающий в себя изучение биологии вида, оказывается более информативным в демографических исследованиях и исследованиях сукцессий, что имеет большое значение для интерпретации моделей сосуществования видов в сообществе (Niinemets, 2004; Flores, 2014).
В популяционной экологии растений представляется возможным использовать функциональные признаки как показатели, связанные с биологическими особенностями вида конкретной жизненной формы. В зависимости от биоморфы анализ внутри- и межпопуляционной изменчивости функциональных признаков на эколого-ценотическом градиенте позволит определить уровень их варьирования и скоординированность, а их совместное использование с популяционными параметрами будет отражать успешность самоподдержания и жизнеспособность популяций. Такие исследования могут использоваться для мониторинга популяций как на отдельных естественных полигонах, так и на экспериментальных участках.
В качестве объекта исследования нами выбран травянистый моноцентрический многолетник – Panzerina lanata (L.) Soják (Панцерина шерстистая) (Lamiaceae). Северная граница P. lanata проходит в широтном направлении с востока на запад от степей Бурятии до степей Горного Алтая. Вид встречается от среднегорий до высокогорий на каменистых и песчаных субстратах как в ненарушенных, так и сильно нарушенных сообществах (Kupriyanova, 1954; Kuminova, 1960, 1976; Ershova, Namzalov, 1985). Если вопрос диверсификации и генетического полиморфизма этого вида в основном решен (Wang et al., 2016; Zhao et al., 2018), то остаются не выясненными механизмы устойчивого развития его популяций, дающие возможность P. lanata успешно существовать в степном поясе гор юга Сибири. Решение этой задачи, с одной стороны, позволит раскрыть особенности адаптации P. lanata в разных экологических и ценотических условиях, а с другой – оценить потенциал его природных популяций. Цель исследования – анализ функциональных признаков и их структурной связи в ценопопуляциях P. lanata, расположенных в различных экологических и ценотических условиях гор юга Сибири.
МАТЕРИАЛЫ И МЕТОДЫ
Panzerina lanata – травянистый многолетник, гемикриптофит, развивающийся по симподиальной полурозеточной модели побегообразования (Cheryomushkina, Astashenkov, 2014). В северо-западной части ареала, охватывающего Горный Алтай и Туву, P. lanata входит в состав каменистых и песчаных степей, реже встречается в луговых степях, на засоленных и остепненных лугах и залежах (Kupriyanova, 1954; Kuminova, 1960, 1976; Ershova, Namzalov, 1985). В данном исследовании использован ранее опубликованный материал авторов, в котором отражены основные биологические особенности вида (Astashenkov, Cheryomushkina, 2006; Cheryomushkina, Astashenkov, 2014).
Ценопопуляции P. lanata изучены на территории Тувы (ценопопуляции 1, 2, 3, 4) и Горного Алтая (ценопопуляции 5, 6, 7, 8, 9). Климат районов исследования характеризуется как резко континентальный, однако отличается некоторыми особенностями. В Туве абсолютный минимум температур приходится на январь: –45...–50°С, максимум – на июль: +50°С. Среднегодовые температуры воздуха зависят от абсолютной высоты местности и географического положения. Южные районы Тувы, где были сосредоточены основные наши исследования, оказываются самыми теплыми, в отличие от горных систем северо-западной части. По существующему природному районированию большая часть территории Тувы относится к единой засушливой зоне с гидротермическим коэффициентом 1.0–0.4. Среднегодовое количество осадков колеблется от 144 до 330 мм; 80–98% осадков выпадает за теплые сезоны года; осадки носят ливневый характер. В районах исследования, которые сосредоточены в Юго-Восточном Алтае, средняя температура января колеблется от –28°С до – 32°С; летние температуры невысоки: средняя температура июля варьирует от 13°С до 18°С. В теплое время минимум осадков приходится на май, максимум на июль–август. Различие между малыми зимними и значительными летними осадками составляет 85–95%. Среднегодовое количество осадков колеблется от 100 до 250 мм (Namzalov, 2015; Makunina, 2016).
Описание сообществ с участием P. lanata проводили по общепринятым в геоботанике методам (Polevaya…, 1964). Нарушенность сообщества определяли по участию сорных видов (Ershova, 1992, 1995; Gorchakovskij, 1999). Латинские названия растений приведены в соответствии с The Plant List (2019). Краткая эколого-ценотическая характеристика мест обитания приведена в табл. 1.
Таблица 1.
Краткая характеристика местообитаний изученных ценопопуляций Panzerina lanata Table 1. Brief ecological-phytocenotic characteristics of habitats of studied Panzerina lanata cenopopulations (CP)
| № ЦП CP no. |
Район исследования Research area | Тип растительности/ особенность субстрат Vegetation type/substrate feature | Видовой состав сообщества/Species composition of community | |
|---|---|---|---|---|
| 1 | Республика Тыва/Republic of Tyva | N 50°12'10.4" E 95°08'13.9" Надпойменная терраса р. Эрзин, окр. пос. Эрзин, Эрзинский р-н. h = 1100 м над ур. м. Above-floodplain terrace of the Erzin River, near Erzin village, Erzinsky dis-trict. h = 1100 m a.s.l. |
Ненарушенная степь/песчаный субстрат Undisturbed steppe/sandy substrate |
Сообщество/Community: разнотравно-караганово-панцериновое. Общее проективное покрытие/Total projective cover – 12%. Число видов/Number of species – 10, из них/of them: Caragana pygmaea (L.) DC – 1%, C. bungei Ledeb. – 1%, Panzerina lanata – 6%, Agropyron desertorum (Fisch. ex Link) Schult. – 2%, Stipa krylovii Roshev. – 1%, Serratula centauroides L. – 0.5%, Potentilla acaulis L. – 0.2%, Eremogone meyeri (Fenzl) Ikonn. – 0.2%. Численность особей Panzerina lanata 21.3 ± 0.7 экз./1 м2/Number of Panzerina lanata plants 21.3 ± 0.7 ind./1 m2 |
| 2 | N 50°12'54.5" E 95°24'41.4" Долина р. Нарын, окр. пос. Нарын, Эрзинский р-н. h = 1216 м над ур. м. Naryn River valley, near Naryn village, Erzinsky district. h = 1216 m a.s.l. |
Ненарушенная степь/песчаный субстрат Undisturbed steppe/sandy substrate |
Сообщество/Community: бесстебельнолапчатково-змеевковое с Caragana bungei. Общее проективное покрытие/Total projective cover – 25%. Число видов/Number of species – 12, из них/of them: Caragana bungei – 4%, Cleistogenes squarrosa (Trin. ex Ledeb.) Keng – 6%, Artemisia tomentella Trautv. – 4%, Panzerina lanata – 4%, Potentilla acaulis – 3%, Dracocephalum foetidum Bunge – 2%. Численность особей Panzerina lanata 8.5 ± 0.2 экз./1 м2/Number of Panzerina lanata plants 8.5 ± 0.2 ind./1 m2 | |
| 3 | N 50°18'58.4" E 90°41'03.5" Перевал Коге-Дава, хребет Цаган-Шибэту, Улуг-Хемский р-н. h = 2246 м над ур. м. Kogeh-Dawa Pass, Tsagan-Shibetu Ridge, Ulug-Khemsky district. h = 2246 m a.s.l. |
Ненарушенная высокогорная степь/подвижная каменистая осыпь с глинистой почвой Undisturbed high-altitude step-pe/ mobile rocky scree with clayey soil |
Сообщество/Community: разнотравно-злаково-луковое. Общее проективное покрытие/Total projective cover – 25%. Число видов/Number of species – 19, из них/of them: Allium altaicum Pall. – 5%, Festuca kryloviana Reverd. – 3%, Stipa orientalis Trin. – 2%, Agropyron cristatum (L.) Gaertn. – 2%, Panzerina lanata – 5%, Artemisia santolinifolia Turcz. ex Bess. – 2%, Gypsophila patrinii Ser. – 1%. Численность особей Panzerina lanata 7.2 ± 0.4 экз./1 м2/Number of Panzerina lanata plants 7.2 ± 0.4 ind./1 m2 | |
| 4 | N 51°15'41.0" E 91°29'59.1" Дорога Хондергей–Чадан, Овюрский р-н. h = 821 м над ур. м. Hondergey–Chadan road, Ovyursky district. h = 821 m a.s.l. |
Нарушенная степь (зарастающая залежь)/песчаный субстрат Disturbed steppe (overgrown fallow)/sandy substrate | Сообщество/Community: разнотравно-полынно-панцериновое. Общее проективное покрытие/Total projective cover – 45%. Число видов/Number of species – 12, из них/of them: Panzerina lanata – 20%, Artemisia scoparia Waldst. et Kit. – 7%, A. glauca Pall. ex Willd. – 5%, A. frigida Willd. – 2%, Stipa krylovii – 2%, Sonchus oleraceus L. – 3%, Potentilla bifurca L. – 3%, Galatella angustissima (Tausch) Novopokr. – 1%. Численность особей Panzerina lanata 15.4 ± 0.7 экз./1 м2/Number of Panzerina lanata plants 15.4 ± 0.7 ind./1 m2 | |
| 5 | N 49°43′64.2″ E 88°24′74.3″ Ущелье р. Тархата, склон хребта Сайлюгем, Кош-Агачский р-н. h = 2347 м над ур. м. Gorge of the Tarkhata River, slope of Sailugem Ridge, Kosh-Agachsky district. h = 2347 m a.s.l. |
Ненарушенная высокогорная степь/каменистый субстрат Undisturbed high-altitude step-pe/rocky substrate |
Сообщество/Community: разнотравно-дерновиннозлаковое. Общее проективное покрытие/Total projective cover – 20%. Число видов / Number of species – 16, из них/of them: Agropyron cristatum – 4%, Festuca kryloviana – 3%, Poa attenuata Trin. – 3%, Panzerina lanata – 3%, Artemisia frigida – 2%, Thalictrum foetidum L. – 0.8%, Pedicularis anthemifolia Fisch. ex Colla – 0.1%, Saussurea leucophylla Schrenk – 0.1%. Численность особей Panzerina lanata 3 ± 0.1 экз./1 м2/Number of Panzerina lanata plants 3 ± 0.1 ind./1 m2 | |
| 6 | Республика Алтай/Republic of Altai | N 49°35′44.1″ E 88°44′30.1″ Верхняя терраса р. Чаган-Бургазы, подножье хребта Сайлюгем, Кош-Агачский р-н. h = 2128 м над ур. м. Upper terrace of the Chagan-Burgazy river, foot of Sailugem Ridge, Kosh-Agachsky district. h = 2128 m a.s.l. |
Ненарушенная высокогорная степь/песчаный субстрат Undisturbed high-altitude step-pe/sandy substrate | Сообщество/Community: пырейно-тонконогово-овсяницевое. Общее проективное покрытие/Total projective cover – 30%. Число видов/Number of species – 15, из них/of them: Festuca kryloviana – 8%, Koeleria altaica (Domin) Krylov – 6%, Elytrigia repens L. – 5%, Artemisia rutifolia – 4%, Panzerina lanata – 3%, Scrophularia altaica Murray – 1%. Численность особей Panzerina lanata 2.5 ± 0.3 экз./1 м2/Number of Panzerina lanata plants 2.5 ± 0.3 ind./1 m2 |
| 7 | N 50°04′39.6″ E 89°17′10.0″ Ущелье р. Бугузун, склон хребта Чихачева, Кош-Агачский р-н. h = 2128 м над ур. м. Gorge of the Buguzun River, slope of Chikhachev Ridge, Kosh-Agachsky district. h = 2128 m a.s.l. |
Ненарушенная высокогорная степь/каменистый субстрат Undisturbed high-altitude step-pe/rocky substrate | Сообщество/Community: бесстебельнолапчатково-полынное с Ziziphora clinopodioides Lam. Общее проективное покрытие/Total projective cover – 50%. Число видов/Number of species – 14, из них/of them: Artemisia santolinifolia – 20%, Ziziphora clinopodioides – 15%, Potentilla acaulis – 5%, Panzerina lanata – 3%, Artemisia depauperata Krasch. – 2%, Carex duriuscula C.A. Mey. – 1%, Stellaria dichotoma L. – 1%. Численность особей Panzerina lanata 3 ± 0.3 экз./1 м2/Number of Panzerina lanata plants 3 ± 0.3 ind./1 m2 | |
| 8 | N 50°01′19.4″ E 89°13′39.0″ Ущелье р. Бугузун, склон хребта Чихачева, Кош-Агачский р-н. h = 2173 м над ур. м. Gorge of the Buguzun River, slope of Chikhachev Ridge, Kosh-Agachsky district. h = 2173 m a.s.l. |
Ненарушенная высокогорная степь/каменистый субстрат Undisturbed high-altitude step-pe/rocky substrate | Сообщество/Community: панцериново-житняковое с Berberis sibirica Pall. Общее проективное покрытие/Total projective cover – 25%. Число видов/Number of species – 12, из них/of them: Berberis sibirica – 3%, Agropyron cristatum – 8%, Panzerina lanata – 6%, Artemisia santolinifolia – 2%, Dracocephalum peregrinum L. – 3%, Stellaria dichotoma L. – 2%. Численность особей Panzerina lanata 7.5 ± 0.2 экз./1 м2/ Number of Panzerina lanata plants 7.5 ± 0.2 ind./1 m2 | |
| 9 | N 50°29′04.8″ E 86°37′31.9″ Окр. пос. Иня, подгорная терраса р. Катунь, хребет Салджар, Онгудайский р-н. h = 789 м над ур. м. Vicinity of Inya village, foothill terrace of the Katun River, Saljar Ridge, Ongudai district. h = 789 m a.s.l. |
Ненарушенная степь/каменистый субстрат Undisturbed steppe/rocky substrate | Сообщество/Community: закустаренное стоповидноосоково-житняковое. Общее проективное покрытие/Total projective cover – 20%. Число видов/ Number of species – 15, из них/of them: Caragana pygmaea – 2%, C. pygmaea subsp. altaica (Kom.) Bondareva – 2%, Spiraea media Franz Schmidt – 1%, Agropyron kazachstanicum (Tzvelev) Peschkova – 3%, Carex pediformis C.A. Mey. – 3%, Artemisia santolinifolia – 2%, Thymus mongolicus (Ronniger) Ronniger – 1%, Panzerina lanata – 3%, Dianthus versicolor Fisch. ex Link – 0.5%, Bupleurum bicaule Helm – 0.5%. Численность особей Panzerina lanata 2.5 ± 0.1 экз./1 м2/Number of Panzerina lanata plants 2.5 ± 0.1 ind./1 m2 | |
Исходя из жизненной формы и особенностей биологии вида выбраны 10 функциональных (организменных) признаков: длина генеративного побега, число ветвей каудекса, число генеративных побегов, длина соцветия, число мутовок в соцветии, число цветков в соцветии, масса особи (в воздушно-сухом состоянии), масса соцветия (в воздушно-сухом состоянии), репродуктивное усилие особи, потенциальная семенная продуктивность особи.
Для статистического анализа регулярным/случайным образом по трансекте шириной 1 м в контуре изучаемого сообщества полностью выкапывали 25–30 особей, находящихся в зрелом генеративном состоянии. С каждой площадки 1 м2 выкапывали 1–2 экземпляра, при высокой численности брали растения с каждой площадки, при низкой – через 1–2 метра. Собранные растения гербаризировали, в дальнейшем эти особи использовали для биометрических вычислений. Для анализа биометрических значений из всей побеговой системы исследовался один генеративный побег с наиболее развитыми структурными элементами. Длину побега измеряли от его базальной части до верхнего метамера соцветия. Для выявления потенциальной семенной продуктивности особи подсчитывали число цветков на всех генеративных побегах особи, затем полученное число умножали на 4 (у всех представителей сем. Lamiaceae в каждом из четырех частей гинецея находится по одному фертильному семязачатку). Показатели биомассы включали в себя надземные (фотосинтезирующую часть) и подземные (каудекс и главный корень) органы растений. Репродуктивное усилие определяли, как отношение биомассы соцветия к биомассе всей особи, включая надземную и подземную части, полученное значение переведено в проценты (Markov, Pleshchinskaya, 1987). Значение числа ветвей каудекса получены в результате прямого подсчета сохранившихся его многолетних глав. Календарный возраст особей определяли с помощью прямого подсчета годичных колец на анатомическом срезе корня.
С целью изучения онтогенетической структуры закладывали учетные площадки 1 м2, число которых варьировало от 30 до 250 в зависимости от численности особей в конкретной ценопопуляции (чем меньше плотность особей в ценопопуляции, тем больше число площадок). Для анализа ценопопуляции выбраны следующие параметры: доля каждого онтогенетического состояния (от ювенильного до сенильного “j, im, v, g1, g2, g3, ss, s”). Ординацию по организменным признакам проводили по средним значениям параметров 25 измеренных особей. Для ординации изученных ценопопуляций по организменным и популяционным параметрам использовали метод главных компонент в пакете программы Statistica 6.0. Поскольку исследованные параметры имеют разную размерность, перед определением главных компонент проводили нормировку организменных признаков путем центрирования их значений к средним арифметическим (Zhivotovskii, 1979). Для значений популяционных параметров встречаемости разных онтогенетических состояний использовали арксинус-преобразование. Для оценки влияния степени нарушенности сообществ использовали ранговый дисперсионный анализ для групп неравного объема по методу Краскелла и Уоллиса (Sokal, Rohlf, 2012).
Для каждой выборки вычислена матрица корреляций, характеризующая связи всех 10 функциональных признаков друг с другом. В дальнейшем использовали метод анализа корреляционных матриц и связей признаков, базирующийся на работах Н.С. Ростовой (Rostova, 2002). Полученные корреляционные матрицы сравнивали по силе связей и их структуре. Для оценки среднего уровня связей использовали коэффициент детерминации, усредненный по всей матрице или по отдельным признакам. В качестве показателя сходства корреляционных матриц по структуре применяли их ординацию с помощью метода главных компонент, который проводили по z‑преобразованным матрицам для приближения распределения коэффициентов корреляции к нормальному (Rostova, 2002).
Степень асимметричности распределения показателей организменных признаков вычисляли через отношение моментов третьего порядка к кубу среднего квадратичного отклонения. Оценка достоверности показателя асимметрии производилась через отношение показателя асимметрии к его ошибке. Значения показателей асимметрии морфологических признаков в разных ценопопуляциях варьировали от –0.78 до 0.97, ошибка – от 0.46 до 0.50. Распределение изученных функциональных признаков в разных ценопопуляциях достаточно близко к симметричному.
Программа построения корреляционных колец была любезно предоставлена П.А. Лезиным (ЗИН РАН). Для характеристики функциональных признаков в ценопопуляции сопоставляли относительную изменчивость (коэффициент вариации) и ее согласованность (коэффициент детерминации).
Анализ онтогенетической структуры ценопопуляции проводили по общепринятым методикам (Rabotnov, 1950; Uranov, 1975; Cenopo-pulyacii…, 1988). Сравнение онтогенетических спектров осуществляли по критерию сходства Л.А. Животовского (Zhivotovskii, 1979).
ОРДИНАЦИЯ ЦЕНОПОПУЛЯЦИЙ
По всем изученным ценопопуляциям основная часть изменчивости функциональных признаков и популяционных параметров (56.2%) включается в первые две главные компоненты (рис. 1а,b): общие размеры особей (F1 = 37.8% дисперсии) и соотношение особей разных онтогенетических состояний (F2 = 18.4% дисперсии). Наибольшие различия по первой главной компоненте проявляются между ценопопуляциями из разных типов сообществ: самые крупные и сильно разветвленные особи с большим числом цветков, высоким репродуктивным усилием и большой потенциальной семенной продуктивностью характерны для ненарушенных песчаных степей (ценопопуляции 1, 2). Противоположное положение занимают особи ценопопуляций, расположенных в каменистых, в том числе высокогорных, степях (ценопопуляции 5–9), а также в нарушенной песчаной степи (ценопопуляция 4), промежуточное – особи ценопопуляции 3, произрастающие на подвижном осыпном склоне. В анализируемых ненарушенных песчаных степных сообществах в долинах рек локальное увлажнение и хорошая аэрация песчаного субстрата, скорее всего, приводят к увеличению значений элементов надземной сферы, в отличие от каменистых степей, где недостаток локального увлажнения и температурный режим заметно сказываются на развитии особей в целом, что отражается в низких показателях признаков (табл. 2).
Рис. 1.
Распределение ценопопуляций Panzerina lanata в плоскости главных компонент (а) и факторная структура изменчивости признаков (b).
По оси X – первая главная компонента (PC1), по оси Y – вторая главная компонента (PC2).
А – длина генеративного побега, B – число ветвей каудекса, C – число генеративных побегов, D – длина соцветия, E – число мутовок в соцветии, F – число цветков в соцветии, G – масса особи (в воздушно-сухом состоянии), H – масса соцветия (в воздушно-сухом состоянии), I – репродуктивное усилие особи, J – потенциальная семенная продуктивность особи. ЦП 1–9 – номера ценопопуляций.
Fig. 1. Distribution of Panzerina lanata coenopopulations in the principal component plane (a) and the factor structure of traits variability (b).
X-axis: first principal component (PC1), Y-axis: second principal component (PC2).
A – generative shoot length, B – number of branches of caudex, C – number of generative shoots, D – inflorescence length, E – number of whorls in inflorescence, F – number of flowers in inflorescence, G – plant weight (air-dry), H – inflorescence weight (air-dry), I – reproductive effort of plant, J – potential seed productivity of plant. 1–9 – coenopopulation numbers.

Таблица 2.
Значения параметров организменных признаков Panzerina lanata Table 2. Values of individual characters in Panzerina lanata
| 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 |
|---|---|---|---|---|---|---|---|---|
| Длина генеративного побега, см/Generative shoot length, сm | ||||||||
| 24.5 ± 1.0 20.09 |
20.3 ± 0.9 22.54 |
23.3 ± 1.1 24.46 |
13.0 ± 0.8 26.17 |
18.3 ± 0.8 20.19 |
24.6 ± 0.9 17.14 |
17.0 ± 0.7 19.99 |
18.0 ± 0.9 25.75 |
20.2 ± 1.1 26.67 |
| Число ветвей каудекса, шт./Number of caudex branches | ||||||||
| 6.1 ± 0.7 55.83 |
3.4 ± 0.3 47.98 |
2.6 ± 0.3 56.57 |
2.1 ± 0.2 71.26 |
2.6 ± 0.3 57.23 |
3.7 ± 0.3 32.72 |
3.8 ± 0.3 40.79 |
3.1 ± 0.3 46.51 |
2.2 ± 0.3 58.09 |
| Число генеративных побегов, шт./Number of generative shoots | ||||||||
| 8.3 ± 0.8 48.41 |
7.9 ± 0.7 45.30 |
6.6 ± 0.4 29.78 |
4.1 ± 0.4 42.47 |
5.5 ± 0.3 28.58 |
7.6 ± 0.6 38.58 |
6.5 ± 0.7 54.36 |
5.8 ± 0.6 64.78 |
3.4 ± 0.3 47.78 |
| Длина соцветия, см/Inflorescence length, сm | ||||||||
| 10.7 ± 0.6 25.89 |
6.2 ± 0.5 35.51 |
10.4 ± 0.5 24.98 |
3.7 ± 0.4 50.29 |
7.3 ± 0.6 38.85 |
11.1 ± 0.6 33.43 |
7.2 ± 0.3 20.40 |
7.7 ± 0.6 38.80 |
6.3 ± 0.5 40.91 |
| Число мутовок в соцветии, шт./Number of whorls per inflorescence | ||||||||
| 5.3 ± 0.4 33.09 |
4.2 ± 0.3 29.29 |
4.9 ± 0.2 17.69 |
3.0 ± 0.2 31.55 |
4.2 ± 0.3 28.43 |
5.3 ± 0.2 22.31 |
3.5 ± 0.1 19.8 |
3.5 ± 0.2 33.33 |
3.8 ± 0.3 41.31 |
| Число цветков в соцветии, шт./Number of flowers per inflorescence | ||||||||
| 12.4 ± 1.1 41.83 |
13.3 ± 0.6 20.74 |
12.9 ± 0.5 19.68 |
11.0 ± 0.5 20.65 |
10.9 ± 0.4 15.37 |
14.9 ± 0.6 18.36 |
12.2 ± 0.4 17.79 |
12.3 ± 0.5 19.74 |
12.5 ± 0.5 20.47 |
| Масса особи, г/Plant weight, g | ||||||||
| 36.2 ± 3.9 49.92 |
24.7 ± 2.4 59.50 |
12.0 ± 0.8 33.08 |
6.2 ± 0.6 43.46 |
8.7 ± 0.5 26.42 |
14.0 ± 1.4 47.43 |
10.2 ± 1.1 56.87 |
7.2 ± 0.4 33.74 |
6.3 ± 0.6 48.72 |
| Масса соцветия, г/Inflorescence weight, g | ||||||||
| 14.3 ± 1.9 64.18 |
11.6 ± 1.1 57.52 |
6.1 ± 0.5 36.80 |
1.7 ± 0.3 70.09 |
2.9 ± 0.1 36.14 |
6.5 ± 0.7 54.73 |
2.3 ± 0.3 65.28 |
2.4 ± 0.1 54.49 |
2.1 ± 0.2 43.88 |
| Репродуктивное усилие, %/Reproductive effort, % | ||||||||
| 38.1 ± 2.4 46.76 |
46.8 ± 1.8 18.61 |
49.9 ± 1.5 35.69 |
25.9 ± 2.0 36.15 |
34.8 ± 1.7 41.58 |
45.7 ± 2.0 21.99 |
24.1 ± 1.8 83.19 |
33.7 ± 2.0 56.70 |
37.8 ± 3.7 49.26 |
| Потенциальная семенная продуктивность, шт./Number of ovules per plant | ||||||||
| 2328.8 ± 397.4 91.52 |
1900.7 ± 281.3 71.15 |
1616.3 ± 84.5 26.13 |
616.2 ± 92.0 68.09 |
1076.9 ± 126.8 51.61 |
2675.3 ± 377.1 68.16 |
1185.8 ± 170.9 68.89 |
973.7 ± 128.1 56.70 |
696.2 ± 101.7 74.52 |
Примечание. Данные приведены в виде M ± m/V. M – среднее арифметическое значение признака, m – ошибка среднего арифметического, V – вариабельность. 1–9 – номера ценопопуляций (характеристика ценопопуляций приведена в таблице 1).
Note. The data are given in the form M ± m/V. M – mean value, m – mean error, V – variability. 1–9 – numbers of populations: see Table 1 for their characteristics.
По значениям F2 наиболее отличаются ценопопуляции по их онтогенетическому составу: максимальная доля особей от ювенильного до молодого генеративного состояний (j–g1) и минимальная доля зрелых и старых генеративных особей (g2 и g3) отмечаются в ценопопуляциях 3, 4, 8, 9, противоположная ситуация – в ценопопуляциях 1, 2, 5 и 7. В первом случае, это связано с тем, что молодые генеративные (g1) и особи прегенеративного периода (j, im, v) характеризуются ускоренными темпами развития и быстрым переходом к цветению и элиминацией растений в последующих состояниях; во втором случае – с периодичностью инспермации и растянутыми темпами развития особей g2, g3 состояний (табл. 3). Доминирующая группа j–g1 отмечена во всех ценопопуляциях (48.5–83.4%), кроме 2 (34.5%) и 5 (19.5%). Несмотря на то, что в целом эта фракция особей преобладает в большинстве ценопопуляций, доля особей отдельных онтогенетических состояний очень динамична. Такое распределение говорит о возможности пополнения семенами во всех ценопопуляциях при отсутствии или малой доле особей g2–g3 состояний.
Таблица 3.
Темпы развития особей Panzerina lanata в разных условиях обитания Table 3. Rate of development of Panzerina lanata plants in different habitat conditions
| Местообитания/Habitats | Прегенеративный период (j–v), лет Pregenerative period (j–v), years |
Генеративный период (g1–g3), лет Generative period (g1–g3), years |
Постгенеративный период (ss–s), лет Postgenerative period (ss-s), years |
|
|---|---|---|---|---|
| Ненарушенные степи Undisturbed steppes |
песчаный субстрат sandy substrate |
6–9 | 10–15 | 2–3 |
| каменистый субстрат rocky substrate |
4–6 | 9–12 | 2–3 | |
| Нарушенные степи Disturbed steppes |
песчаный субстрат sandy substrate |
3–4 | 6–7 | 1 |
ОБЩАЯ И СОГЛАСОВАННАЯ ИЗМЕНЧИВОСТЬ ПРИЗНАКОВ
Изученные ценопопуляции распределились на 3 группы: ненарушенные песчаные степи (1, 2, 6), ненарушенные каменистые, в том числе высокогорные, степи (3, 5, 7–9) и нарушенная песчаная степь (4). Применение рангового дисперсионного анализа позволило разделить организменные признаки на две группы. К первой относятся признаки, параметры которых достоверно (χ2 >11.8 при P = 0.05) зависят от нарушенности сообщества: это число генеративных побегов, масса особей и соцветий, потенциальная семенная продуктивность. Для остальных признаков внутрипопуляционные различия особей преобладают над межпопуляционными (χ2 < 1.5 при P = 0.05). Следовательно, основные структурные изменения особей в нарушенных и ненарушенных сообществах выражаются в таких признаках, как число генеративных побегов, масса особей и соцветий, потенциальная семенная продуктивность.
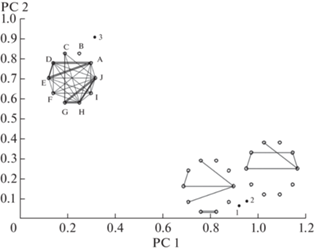
Анализ индивидуальной изменчивости 10 изученных признаков Panzerina lanata показал, что максимальной общей и согласованной изменчивостью характеризуется число семязачатков у особи (потенциальная семенная продуктивность) (рис. 2). Такие признаки, как число мутовок в соцветии, длина соцветия и длина генеративного побега обладают высокой скоррелированностью с другими признаками при относительно незначительном варьировании. Изменения этих признаков являются ключевыми для особи в целом, поскольку именно они представляют собой основные компоненты, определяющие потенциальную семенную продуктивность особи.
Рис. 2.
Общая и согласованная изменчивость морфологических признаков Panzerina lanata.
По оси Х – согласованная изменчивость (коэффициент детерминации) признаков, по оси Y – общая изменчивость (коэффициент вариации) признаков.
Условные обозначения признаков (A–J) приведены в рис. 1.
Fig. 2. General and concordant variability of morphological traits in Panzerina lanata.
X-axis: concordant variability (coefficient of determination) of the traits; Y-axis: general variability (coefficient of variation) of the traits.
See Fig. 1 for symbols of the traits (A–J).

Самой низкой вариабельностью и уровнем связей с другими признаками обладает число цветков в соцветии. Низкое варьирование этого признака мы связываем с емкостью почки, в которой закладывается определенное число зачатков цветков. Такой признак, как число ветвей каудекса характеризуется средним варьированием, которое слабо согласовано с изменениями других признаков. Ранее было установлено, что разветвленность каудекса зависит от особенностей субстрата и интенсивности побегообразования (Cheryomushkina, Astashenkov, 2014). Так, в песчаных ненарушенных степях за счет подвижности субстрата формируется онтобиоморфа с “рыхлым” многоглавым каудексом, у которого длинные резиды несут большое число почек возобновления. В ненарушенных каменистых степях из-за статичности субстрата образуется онтобиоморфа с компактным многоглавым каудексом. В нарушенных местообитаниях формируется онтобиоморфа с одноглавым компактным каудексом и малым числом почек возобновления.
Исходя из значений общей и согласованной изменчивости, потенциальную семенную продуктивность мы рассматриваем как признак системной адаптивной изменчивости для P. lanata.
СТРУКТУРА СВЯЗЕЙ ПРИЗНАКОВ
Сравнение структуры связей выявило среднюю степень сходства между корреляционными матрицами (67.9%) (рис. 3). При этом в группировке матриц четко обнаруживается соответствие типам сообществ по степени их нарушенности, что отражает, по нашему мнению, значительное влияние экологических и фитоценотических условий на согласованную изменчивость признаков особей. Наиболее сходны матрицы выборок из ненарушенных каменистых и песчаных степей (ценопопуляции 1, 2, 3, 5–9). Сильнее всего отличается матрица из нарушенной песчаной степи (ценопопуляция 4).
Рис. 3.
Распределение выборок ценопопуляций Panzerina lanata в плоскости первых двух главных компонент по корреляционным матрицам.
По оси X – фактор сходства матриц (PC1), по оси Y – фактор специфичности матриц (PC2 1). Сечения корреляционного цилиндра (r > 0.6) приведены для групп ценопопуляций. Сильные связи (r > 0.9) обозначены двойными линиями. 1 – ЦП, расположенные в ненарушенных песчаных степях, 2 – ЦП, расположенные в ненарушенных каменистых степях, 3 – ЦП, расположенная в нарушенной песчаной степи. Условные обозначения признаков (A–J) и ценопопуляций приведены в рисунке 1.
Fig. 3. Distribution of Panzerina lanata coenopopulations in the principal component plane according to correlation matrices.
X-axis: matrix similarity factor (PC1); Y-axis: matrix specificity factor (PC2). The cross sections of the correlation cylinder (r > 0.6) are shown for groups of populations. Double lines indicate strong links (r > 0.9). 1 – coenopopulation of sandy steppes (undisturbed habitats), 2 – coenopopulation of stony steppes (undisturbed habitats), 3 – coenopopulation of sandy steppes (disturbed habitat). See Fig. 1 for symbols of the traits (A–J) and the population numbers (1–9).
Несмотря на различную вариабельность метрических значений признаков, которые зависят от конкретных экологических условий в ценопопуляции, нами выявлена разная степень связи между ними, которая отражает биологические особенности вида и адаптационные механизмы. Так, в ненарушенных сообществах наблюдается довольно низкая их скоррелированность (от 0.12 до 0.19). В нарушенном типе сообщества (зарастающая залежь – ценопопуляция 4) отмечается увеличение как общей, так и согласованной изменчивости признаков особей. В таких условиях скоординированность признаков увеличивается в 2‒3 раза и достигает значения 0.37.
Взаимосвязь между биомассой особей, массой соцветий, репродуктивным усилием и потенциальной семенной продуктивностью, особенно двух последних признаков, отчетливо отражает биологические особенности вида и характер самоподдержания ценопопуляций. Масса соцветий является производным нескольких компонентов – длины соцветия, числа мутовок соцветия, числа цветков в соцветии, числа генеративных побегов у особи и структуры самого генеративного побега. Как показывает анализ корреляционных связей, взаимоотношения между этими признаками не так однозначны, что связано со структурными изменениями синфлоресценции особей, произрастающих в различных условиях. Как было показано ранее (Cheryomushkina, Astashenkov, 2014), у Panzerina lanata возможно формирование шести разных вариантов строения синфлоресценции, различающихся по степени разветвленности и, соответственно, образующих широкий спектр вариаций: от неразветвленных до сильно разветвленных соцветий с большим числом паракладиев. Разнообразие строения генеративных побегов в пределах отдельной особи сильно видоизменяет корреляции между массой соцветий особи и структурными элементами соцветий (длиной соцветия, числом мутовок, цветков в соцветии и генеративных побегов у особи), что демонстрирует различные связи в ненарушенных каменистых, песчаных и нарушенных степях (рис. 3).
В ненарушенных песчаных и каменистых степях увеличение массы репродуктивной сферы у особей достигается разными путями, что отражается в системе корреляций и отсутствии однозначной корреляционной связи между массой соцветия и другими структурами генеративной сферы особей. Отсутствие устойчивых линейных связей между массой соцветий и массой особей в песчаных и каменистых степях мы связываем со структурными изменениями синфлоресценции. Выявленные варианты синфлоресценции (Cheryomushkina, Astashenkov, 2014) обладают разным соотношением между метамерной протяженностью зон обогащения и торможения. У неразветвленных или слабо разветвленных синфлоресценций вклад главного соцветия в общую массу особи будет непропорционально меньше, чем у сильно разветвленных растений, формирующих парциальные соцветия. Так, в условиях ненарушенных песчаных степей с мощно развитыми и разветвленными побеговыми системами (ценопопуляции 1, 2) отмечается сильная устойчивая связь между рассматриваемыми параметрами (r > 0.9). В этих же условиях отмечается высокая потенциальная семенная продуктивность (около 1900–2700 семязачатков на особь в среднем). В условиях ненарушенных каменистых степей образуются менее разветвленные варианты синфлоресценции, что приводит к нарушению корреляционных связей между массой соцветий и массой особей (r < 0.6), и как итог – снижение потенциальной семенной продуктивности особей (около 700–1600 семязачатков на особь в среднем). Неустойчивость корреляций между такими ключевыми для существования этого вида параметрами, как масса соцветий, масса особей, репродуктивное усилие и потенциальная семенная продуктивность оказывает сильное влияние и на характер корреляционных связей других признаков.
Таким образом, в ненарушенных сообществах у особей Panzerina наблюдается довольно низкая скоррелированность всех признаков; в нарушенном (зарастающая залежь) отмечается увеличение как общей, так и согласованной изменчивости признаков.
ИЗМЕНЕНИЕ СИСТЕМЫ КОРРЕЛЯЦИЙ ПРИЗНАКОВ В ЗАВИСИМОСТИ ОТ УСЛОВИЙ ОБИТАНИЯ
Изменение экологических и ценотических условий сказывается как на морфометрических показателях признаков и их изменчивости у особей Panzerina lanata, так и на перестройке системы их корреляций.
В ненарушенных каменистых (ценопопуляции 3, 5, 7–9) и песчаных степях (ценопопуляции 1, 2, 6) особи характеризуются, с одной стороны, средними или близкими к ним показателями большинства признаков, с другой стороны – сходной структурой корреляций (рис. 3). В ненарушенных сообществах вид находится в естественных условиях, испытывая выровненное влияние со стороны фитоценотического окружения, в отличие от нарушенного сообщества (ценопопуляция 4), в котором протекают сукцессионные процессы. В ненарушенном местообитании отмечается довольно низкая скоррелированность между признаками, что связано с многочисленными структурными перестройками тела особей, слабо скоординированными друг с другом. Такая низкая скоррелированность дает возможность особям формировать различные морфоструктуры, связанные с репродуктивной способностью вида. Одновременно с этим влияние конкретного экотопа приводит к поливариантности онтогенеза, что увеличивает адаптационные возможности вида.
У особей Panzerina в ценопопуляции 4, находящейся в сукцессионном сообществе, выявлено снижение значений многих признаков (в том числе потенциальной семенной продуктивности) и увеличение размаха их варьирования по сравнению с ненарушенными вариантами песчаных степей. По всей видимости, близкое к пределам нормы реакции вида местообитание приводит к значительному усилению многих корреляционных связей. При этом происходит усиление как сильных, так и слабых связей. Особи в этой ценопопуляции находятся в самых пессимальных условиях существования, что, в первую очередь, сказывается на самых ключевых показателях – репродуктивном усилии и потенциальной семенной продуктивности. В этой ценопопуляции отмечаются самые минимальные показатели семенной продуктивности (50–70 семязачатков на особь). Скорее всего, повышенная гетерогенность особей по степени развития их морфометрических показателей сказывается на увеличение размаха их варьирования, а сукцессионные процессы, протекающие в данном фитоценозе, заставляют особи реагировать на локальные изменения “жесткостью” связей. Таким образом, в условиях зарастающей залежи формируются особи с самыми минимальными показателями и минимальной семенной продуктивностью и одновременно с максимальной скоррелированностью всех признаков.
ОНТОГЕНЕТИЧЕСКАЯ СТРУКТУРА ЦЕНОПОПУЛЯЦИЙ
Анализ соотношения относительного варьирования и детерминированности исследованных признаков показывает, что индикатором изменчивости особей Panzerina lanata является потенциальная семенная продуктивность. Этот признак является биологически значимым для P. lanata, так как обеспечивает устойчивое самоподдержание и успешное развитие ценопопуляции этого вида в разных условиях обитания.
Низкая скоррелированность признаков в ненарушенных сообществах дает возможность особям формировать различные элементы, связанные с репродуктивной способностью вида, это напрямую влияет на процесс инспермации, который обеспечивает различные волны развития в естественных популяциях. Так, полученный усредненный онтогенетический спектр ценопопуляций для всех ненарушенных сообществ (коэффициент сходства R = 0.86 ± 0.015) многовершинный и описывает две волны развития (рис. 4а). Пик первой волны приходится на молодые генеративные особи, затем наблюдается постепенный спад от зрелых генеративных растений до особей постгенеративного периода. Высокое содержание зрелых и старых генеративных особей связано с растянутыми темпами развития; низкое содержание субсенильных и сенильных – с ускоренными темпами. Пик второй волны приходится на ювенильные особи, затем происходит плавный спад на виргинильную группу. Вторая волна развития свидетельствует о периодичности инспермации (при этом периодичность может быть регулярной и нерегулярной). Подобные типы спектров формируются в результате выровненных темпов развития и сбалансированном обороте поколений в ненарушенных сообществах. Также морфологическая поливариантность особей приводит к разнообразию онтобиоморф (с многоглавым, “рыхлым” и компактным, реже одноглавым каудексом), что обеспечивает устойчивость особей в популяциях.
Рис. 4.
Усредненные онтогенетические спектры ценопопуляций Panzerina lanata
a – ненарушенные местообитания, b – нарушенное местообитание
По оси Х – онтогенетические состояния, по оси Y – количество особей на единицу площади, % от общего числа.
Fig. 4. Averaged ontogenetic spectra of Panzerina lanata coenopopulations.
a – undisturbed habitats, b – disturbed habitat.
X-axis: ontogenetic states; Y-axis: number of plants per unit area, % of the total number.

В условиях зарастающей залежи формируется типичный одновершинный левосторонний спектр (рис. 4b) с максимумом на молодых генеративных особях. Левая часть спектра отражает низкую инспермацию и неравномерное развитие особей прегенеративного периода, правая часть спектра – высокую элиминацию растений зрелого генеративного и последующих онтогенетических состояний. Подобный вариант спектра формируется в результате асинхронного развития особей в ценопопуляции и несбалансированного оборота поколений в нарушенных сообществах. Поливариантность развития особей Panzerina в таких сообществах слабо выражена, формируется только одна онтобиоморфа (с одноглавым компактным каудексом), что влечет за собой неустойчивость особей в популяции.
Высокая численность особей (max 21.3 экз/м2) характерна для ненарушенных песчаных степей, низкая – для каменистых высокогорных степей (max 7.2 экз/м2). Различный состав сообществ приводит к колебанию численности в изученных ценопопуляциях. В целом степи на песчаных почвах более благоприятны для возобновления и развития особей P. lanata, чем степи на каменистых почвах.
С помощью функциональных характеристик обычно используемых в литературе (удельная листовая поверхность, водный потенциал, фотосинтез, устьичная проводимость, биомасса особей и т.д.) изучаются внутри- и межвидовая, внутри- и межпопуляционная изменчивость признаков. Ранее, сопоставляя изменчивость системы корреляций и общую согласованность морфологических признаков у различных видов растений, было выявлено изменение структуры связей и индикаторных признаков в зависимости от условий произрастания (Rostova, 1999, 2000). Полученные нами результаты согласуются с данными многих авторов о том, что в нарушенных и неблагоприятных местообитаниях корреляционная связь морфологических признаков выше, чем в ненарушенных и благоприятных условиях. Однако в популяционной биологии важное место занимают анализ и поиск детерминированных признаков, которые отражают биологию вида и являются определяющими для устойчивого развития популяции растений с конкретной жизненной формой.
В большинстве случаев для оценки функционирования отдельных видов используются такие организменные показатели, как биомасса особи (надземная/подземная) или отдельные ее элементы и связанные с ними физиологические процессы, как, например, фотосинтез и дыхание (Cornelissen et al., 2003; Pérez-Harguindeguy et al., 2013; Klimešová et al., 2019; Wigley et al., 2020). Нет сомнений, что биомасса особей и связанные с ней компоненты, являются ключевыми показателями успешности популяций и сообществ, однако в целом эти параметры не в полной мере отражают механизмы функционирования и устойчивость популяции растений. Так, например, для вегетативно-подвижных длиннокорневищных трав устойчивость популяций определяют такие признаки, как длина и скорость развития годичного прироста и интенсивность ветвления корневища (Smirnova, 1987).
Ранее для некоторых травянистых растений разных жизненных форм были установлены макроморфологические признаки (от метамера до особи) и биологические признаки (тип самоподдержания, биомасса, темпы развития особей), дающие возможность охарактеризовать динамические процессы, жизнеспособность популяций и популяционную стратегию видов (Vostochnoevropejskie, 2004; Cheryomushkina, 2004). Наши исследования показали, что одним из индикаторных функциональных признаков можно рассматривать потенциальную семенную продуктивность. Высокая его детерминированность и скоординированность с другими признаками, как, например, у P. lanata, дает возможность использовать его для анализа устойчивости популяций многолетних стержнекорневых растений в различных экологических и ценотических условиях.
ЗАКЛЮЧЕНИЕ
Общая и согласованная изменчивость признаков у Panzerina lanata в большей мере определяются нарушенностью сообщества. В ненарушенных песчаных и каменистых степях наблюдаются средний размах варьирования функциональных признаков и их низкая степень взаимообусловленности. В нарушенной песчаной степи, наоборот, у особей P. lanata признаки имеют высокий размах варьирования и сильные корреляционные связи.
Анализ общей и согласованной изменчивости признаков позволил выявить индикаторный функциональный признак – потенциальную семенную продуктивность, значение которого определяет способность к самоподдержанию ценопопуляции в различных местообитаниях. Установлено, что в ненарушенных вариантах песчаных и каменистых степей, в том числе высокогорных, Panzerina развивает мощные особи с большей разветвленностью побеговой системы и высокой семенной продуктивностью. В нарушенной степи в процессе зарастания залежи P. lanata формирует небольшие, обычно слабоветвящиеся особи с низкой семенной продуктивностью.
Онтогенетическая структура ценопопуляций в ненарушенных местообитаниях отражает результат сбалансированного процесса оборота поколений, который тесно связан с многоуровневой системой взаимодействия элементов особей, выровненными темпами развития растений и регулярной инспермацией. В результате этого в ненарушенных степных сообществах образуются волны развития. Нарушенность фитоценоза приводит к асинхронным темпам развития особей и несбалансированному обороту поколений, низкой и нерегулярной инспермации. Это обуславливает формирование в нарушенных степных сообществах одновершинного левостороннего спектра.
Список литературы
Adler P.B., Salguero-Gómez R., Compagnoni A., Joanna S.H., Ray-Mukherjee J., Mbeau-Ache C., Franco M. 2014. Functional traits explain variation in plant life history strategies. – Proc. Natl. Acad. Sci. USA. 111 (2): 740–745. https://doi.org/10.1073/pnas.1315179111
[Astashenkov, Cheryomushkina] Асташенков А.Ю., Черемушкина В.А. 2006. Онтогенетическая структура ценопопуляций Panzerina lanata subsp. argyracea и P. canescens. – Раст. ресурсы. 42 (3): 1–9.
Bernhardt-Römermann M., Gray A., Vanbergen A.J., Bergès L., Bohner A., Brooker R.W., … Stadler J. 2011. Functional traits and local environment predict vegetation responses to disturbance: A pan-European multi-site experiment. – J. Ecol. 99 (3): 777–787. https://doi.org/10.1111/j.1365-2745.2011.01794.x
Bjorkman A.D., Myers-Smith I.H., Sarah C., Elmendorf S.N., Rüger N., Beck P.S.A., …, Weiher E. 2018. Plant functional trait change across a warming tundra biome. – Nature. 562: 57–62. https://doi.org/10.1038/s41586-018-0563-7
[Cenopopulyacii…] Ценопопуляции растений (очерки популяционной биологии). 1988. М. 182 с.
[Cheryomushkina] Черемушкина В.А. 2004. Биология Луков Евразии. Новосибирск. 279 с.
Cheryomushkina V.A., Astashenkov A.Yu. 2014. Morphological Adaptation of Panzerina Sojak (Lamiaceae) Species to Various Ecological Conditions. – Contemporary Problems of Ecology. 7 (5): 520–525.
Cornelissen J.H.C., Lavorel S., Garnier E., Díaz S., Buchmann N., Gurvich D.E., …, Poorter H. 2003. A handbook of protocols for standardised and easy measurement of plant functional traits worldwide. – Austral. J. Bot. 51 (4): 335–380. https://doi.org/10.1071/BT02124
[Dudova et al.] Дудова К.В., Атабаллыев Г.Г., Ахметжанова А.А., Гулов Д.М., Елумеева Т.Г., Клинк Г.В. …, Онипченко В.Г. 2019. Высота как функциональный признак альпийских растений. – Бюл. МОИП. Отд. Биол. 124 (1): 33–45.
[Ershova] Ершова Э.А. 1992. Растительность межгорных котловин. В кн.: Антропогенная трансформация растительного покрова Западной Сибири. Новосибирск. С. 35–51.
[Ershova] Ершова Э.А. 1995. Антропогенная динамика растительности юга средней Сибири. Препринт. Новосибирск. 27 с.
[Ershova, Namzalov] Ершова Э.А., Намзалов Б.Б. 1985. Степи. – В кн.: Растительный покров и естественные кормовые угодья Тувинской АССР. Новосибирск. С. 119–149.
Ferraro A., Santos da Silva G., Lima de Aguiar C., Appezzato-da-Glória B. Evaluating belowground bud banks of native species from Cerrado: Structural, chemical, and ecological approaches. Evaluating belowground bud banks of native species from Cerrado: Structural, chemical, and ecological approaches. 2021. – Flora. 281. 151852. https://doi.org/10.1016/j.flora.2021.151852
Flores O., Hérault B., Delcamp M., Garnier É., Gourlet-Fleury S. 2014. Functional Traits Help Predict Post-Disturbance Demography of Tropical Trees. – Plos one. 9 (9): e105022. https://doi.org/10.1371/journal.pone.0105022
González de Andrés E., Rosas T., Camarero J.J., Martínez-Vilalta J. 2021. The intraspecific variation of functional traits modulates drought resilience of European beech and pubescent oak. – J. Ecol. 109 (10): 3652–3669. https://doi.org/10.1111/1365-2745.13743
[Gorchakovskij] Горчаковский П.Л. 1999. Антропогенная трансформация и восстановление продуктивности луговых фитоценозов. Екатеринбург. 156 с.
[Grant] Грант В. 1984. Видообразование у растений. М. 584 с.
Gross N., Börger L., Soriano-Morales S.I., Le Bagousse-Pinguet Y., Quero J.L., García-Gómez M., Valencia-Gómez E., Maestre F.T. 2013. Uncovering multiscale effects of aridity and biotic interactions on the functional structure of Mediterranean shrublands. – J. Ecol. 101 (3): 637–649. https://doi.org/10.1111/1365-2745.12063
Karbstein K., Prinz K., Hellwig F., Römermann C. 2020. Plant intraspecific functional trait variation is related to within-habitat heterogeneity and genetic diversity in Trifolium montanum L. – Ecol. Evol. 10 (11): 5015–5033. https://doi.org/10.1002/ece3.6255
Klimešová J., Martínkováa J., Pausasc J.G., Gomes de Moraesd M., Herbenb T., Yuf F.-H., Puntierih J., …, Ottaviania G. 2019. Handbook of standardized protocols for collecting plant modularity traits. – Ecol. Evol. 7 (5): 1553–1560. https://doi.org/10.1002/ece3.2721
[Kuminova] Куминова А.В. 1960. Растительный покров Алтая. Новосибирск. 449 с.
[Kuminova] Куминова А.В. 1976. Растительный покров Хакасии. Новосибирск. 1976. 423 с.
[Kupriyanova] Куприянова Л.А. 1954. Род Panzerina Moench. – В кн.: Флора СССР. Т. 21. М., Л. С. 158–159.
[Kurchenko] Курченко Е.И. 1985. Динамика ценопопуляций и микроэволюционный процесс (на примере полевиц (Agrostis L.). – В кн.: Динамика ценопопуляций растений. М. С. 174–186.
[Kurchenko] Курченко Е.И. 2010. Род полевица (Agrostis L., сем. Poaceae) России и сопредельных стран. М. 516 с.
[Markov, Pleshchinskaya] Марков М.Вит., Плещинская Е.Н. 1987. Репродуктивное усилие у растений. – Журн. общ. биол. 48 (1): 77–83.
[Makunina] Макунина Н.И. 2016. Растительность лесостепи Западно-Сибирской равнины и Алтае-Саянской горной области. Новосибирск. 183 с.
[Namzalov] Намзалов Б.Б. 2015. Степи Тувы и Юго-Восточного Алтая. Новосибирск. 294 с.
Niinemets Ü. 2004. Adaptive adjustments to light in foliage and whole-plant characteristics depend on relative age in the perennial herb Leontodon hispidus. – New Phytol. 162 (3): 683–696. https://doi.org/10.1111/j.1469-8137.2004.01071.x
[Onipchenko] Онипченко В.Г. 2014. Функциональная фитоценология: Синэкология растений. Учебное пособие. М. 576 с.
[Onipchenko et al.] Онипченко В.Г., Казанцева Е.С., Елумеева Т.Г., Захарова Е.А., Петрова С.Е., Ахметжанова А.А., Царегородцева А.А., Текеев Д.К. 2020. Абиотические факторы сильнее влияют на функциональные признаки, чем биотические: эксперименты и пересадки в высокогорьях. – Журн. общ. биол. 81 (5): 352–361. https://doi.org/10.31857/S0044459620050097
[Polevaya…] Полевая геоботаника Т. 3. М., Л. 1964. 535 с.
Pérez-Harguindeguy N., Díaz S., Garnier E., Lavorel S., Poorter H., Jaureguiberry P., …, Cornelissen J.H.C. 2013. New handbook for standardised measurement of plant functional traits worldwide. – Austral. J. Bot. 61 (3): 167–234. https://doi.org/10.1071/BT12225
[Rabotnov] Работнов Т.А. 1950. Жизненный цикл многолетних травянистых растений в луговых ценозах. – Труды БИН АН СССР. Геоботаника. 3 (6): 176–196.
Reich P.B., Walters M.B., Ellsworth D.S. 1997. From tropics to tundra: global convergence in plant functioning. – Proc. Natl. Acad. Sci. USA. 94 (25): 13730-4. https://doi.org/10.1073/pnas.94.25.13730
Römermann C., Bucher S.F., Hahn M., Bernhardt-Römermann M. 2016. Plant functional traits – fixed facts or variable depending on the season. – Folia Geobotanica. 51 (2): 143–159. https://doi.org/10.1007/s12224-016-9250-3
[Rostova] Ростова Н.С. 1999. Изменчивость системы корреляций морфологических признаков. 1. Естественные популяции Leucanthemum vulgare (Asteraceae). – Бот. журн. 84 (11): 50–66.
[Rostova] Ростова Н.С. 2000. Изменчивость системы корреляций морфологических признаков. 2. Популяции Leucanthemum vulgare (Asteraceae) в природе и в условиях культивирования. – Бот. журн. 85 (1): 46–67.
[Rostova] Ростова Н.С. 2002. Корреляции: структура и изменчивость. СПб. 308 с.
[Smirnova] Смирнова О.В. 1987. Структура травяного покрова широколиственных лесов. М. 206 с.
[Smirnova et al.] Смирнова О.В., Заугольнова Л.Б., Пападюк Р.В. 1993. Популяционная концепция в биогеоценологии. – Журн. общ. биол. 54 (3): 438–448.
Stebbins G.L. 1974. Flowering Plants: Evolution above the species level. Cambridge. 399 p.
Sokal R.R., Rohlf F.J. 2012. Biometry: the principles and practice of statistics in biological research. New York. 937 p.
The Plant List. 2019. www.theplantlist.org (Accessed 10.10.2019).
[Uranov] Уранов А.А. 1975. Возрастной спектр фитоценопопуляций как функция времени и энергетических волновых процессов. – Биол. науки. 2: 7–34.
[Vostochnoevropejskie…] Восточноевропейские леса: история в голоцене и современность. 2004. Т. 1. М. 479 с.
Wang P., Zhang X.Z., Tang N., Liu J.J., Xu L.R., Wang K. 2016. Phylogeography of Libanotis buchtormensis (Umbelliferae) in Disjunct Populations along the Deserts in Northwest China. – Plos One. 11: e0159790. https://doi.org/10.1371/journal.pone.0159790
Wigley B.J., Charles-Dominique T., Hempson G.P., Stevens N., TeBeest M., Archibald S., Bond W.J., Zhou Y., Kruger L.M. 2020. A handbook for the standardised sampling of plant functional traits in disturbance-prone ecosystems, with a focus on open ecosystems. – Austral. J. Bot. 68 (8) 473–531. https://doi.org/10.1071/BT20048
Wood C.M., McKinney S.T., Loftin C.S. 2017. Intraspecific functional diversity of common species enhances community stability. – Ecol. Evol. 7 (5): 1553–1560. https://doi.org/10.1002/ece3.2721
Xi N., Adler P.B., Chen D., Wu H., Catford J.A., Bodegom P.M., Bahn M., Crawford K.M., Chu C. 2021. Relationships between plant–soil feedbacks and functional traits. – J. Ecol. 109 (9): 3411–3423. https://doi.org/10.1111/1365-2745.13731
[Zaugol’nova] Заугольнова Л.Б. 1994. Структура популяций семенных растений и проблемы их мониторинга: Автореф. дис. … док. биол. наук. СПб. 70 с.
Zhao Y., Zhang H., Pan B., Zhang M. 2019. Intraspecific divergences and phylogeography of Panzerina lanata (Lamiaceae) in northwest China. – PeerJ. 7: e6264. https://doi.org/10.7717/peerj.6264
[Zhivotovskii] Животовский Л.А. 1979. Показатель сходства популяций по полиморфным признакам. – Журн. общ. биол. (40) 4: 587–602.
[Zlobin et al.] Злобин Ю.А., Скляр В.Г., Клименко А.А. 2013. Популяции редких растений: теоретические основы и методы изучения. Сумы. 439 с.
Дополнительные материалы отсутствуют.
Инструменты
Ботанический журнал