

Ботанический журнал, 2022, T. 107, № 9, стр. 906-916
SACCHARINA LATISSIMA (OCHROPHYTA, LAMINARIALES) В ПРИКАМЧАТСКИХ ВОДАХ: ЭКОЛОГИЧЕСКИЕ ОСОБЕННОСТИ, РАСПРОСТРАНЕНИЕ И МОЛЕКУЛЯРНАЯ ФИЛОГЕНИЯ
Т. А. Клочкова 1, *, А. В. Климова 1, **, Г. Х. Ким 2, ***, Н. Г. Клочкова 3, ****
1 Камчатский государственный технический университет
683003 Петропавловск-Камчатский, ул. Ключевская, 35, Россия
2 Национальный университет Конджу
32588 Конджу, Республика Корея
3 Камчатский филиал Тихоокеанского института географии ДВО РАН
683000 Петропавловск-Камчатский, ул. Партизанская, 6, Россия
* E-mail: tatyana_algae@mail.ru
** E-mail: annaklimovae@mail.ru
*** E-mail: ghkim@kongju.ac.kr
**** E-mail: ninakl@mail.ru
Поступила в редакцию 07.06.2021
После доработки 16.07.2022
Принята к публикации 19.07.2022
- EDN: SZDCHD
- DOI: 10.31857/S000681362209006X
Аннотация
Представлены результаты изучения экологических особенностей, морфометрического разнообразия и распространения цельнолистного вида камчатских ламинариевых, ранее указывавшегося в регионе под разными названиями: Laminaria gurjanovae, Laminaria gurjanovae f. lanciformis, Laminaria lanciformis, Laminaria sp. avgub 1, Saccharina gurjanovae. Проведенный нами молекулярно-филогенетический анализ последовательностей COI показал высокий уровень сходства камчатских представителей с образцами Saccharina latissima с атлантического побережья Европы и канадского сектора Арктики – Гудзонова залива (98.7–99% и 99.8%, соответственно), что свидетельствует об их принадлежности к данному виду. Полученные нами молекулярно-филогенетические данные не распространяются на типовую форму L. gurjanovae, заметно отличающуюся от S. latissima, в том числе и от ее камчатских представителей, морфологией и экологическими характеристиками.
Представители порядка Laminariales имеют исключительную хозяйственную ценность и играют важную экосистемную роль. В холодных и умеренных водах Северного полушария в районах с высокой амплитудой приливно-отливных колебаний они являются основными доминантами растительных сообществ нижнего горизонта литорали и сублиторали. Большинство их родов и видов имеют узкое широтное и меридиональное распространение. Изучение морфологического разнообразия, современного распространения и экологических особенностей ламинариевых дает информацию для реконструкции геологической истории Земли и прогнозирования будущих изменений морской биоты в условиях глобальных изменений климата (Climate Change…, 2013; Vtoroy…, 2014; Zhang et al., 2021). Достоверность этих прогнозов во многом зависит от правильного определения видов-доминантов донных сообществ, к числу которых принадлежат представители обсуждаемой группы.
Самыми широко распространенными и многочисленными в порядке Laminariales являются роды Laminaria и Saccharina. Их представители характеризуются простотой организации слоевища. Степень их изменчивости даже в пределах одной популяции определяется сочетанием генетического и адаптивного полиморфизма, а вариабельность их морфологических и анатомических признаков имеет характер постепенных модификаций (Petrov, 1972). Это, в свою очередь, обуславливает значительную перекрываемость морфологических признаков между разными видами и приводит к тому, что, несмотря на длительное изучение таксономии порядка Laminariales, объем родов Laminaria и Saccharina и их распространение в Мировом океане окончательно не установлены.
В ламинариевой альгофлоре Камчатки встречается вид, который разные исследователи определяли как Laminaria saccharina (L.) J.V. Lamour. (Zinova, 1933, 1954a, 1954b; Gayl, 1936), Laminaria gurjanovaе f. lanciformis Petrov (Petrov, 1972), Laminaria gurjanovaе Zinova (Vinogradova et al., 1978), L. lanciformis (Petrov) Kloczkova et Beliyi (Klochkova et al., 2009), Laminaria sp. 1 (Belij, 2013), Laminaria sp. avgub1 (Koneva, Klochkova, 2014), Saccharina gurjanovae (Zinova) Selivanova, Zhigadlova et G.I. Hansen (Selivanova et al., 2007). В настоящей статье для камчатской популяции вида представлены результаты изучения его фенотипических и генотипических признаков, а также экологических особенностей. На основании молекулярно-филогенетического анализа установлена его таксономическая принадлежность.
МАТЕРИАЛЫ И МЕТОДЫ
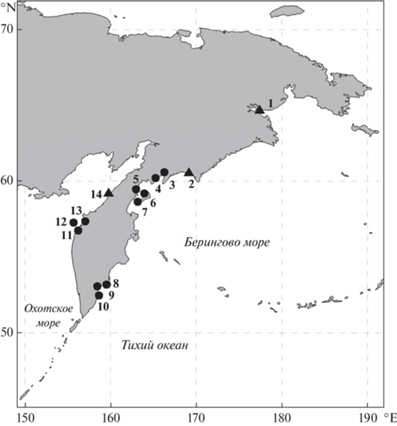
Изучение вида основано на материалах, собранных у Камчатки во время альгопромысловых съемок КамчатНИРО и альгофлористических исследований, проводившихся авторами в разных районах восточного и западного побережий Камчатки в период 2004–2020 гг. (рис. 1). Это позволило получить сведения о его размерно-массовых показателях, возрастной структуре популяций, ценотической роли, требованиях к условиям произрастания, особенностях размножения в разных районах камчатского побережья.
Рис. 1.
Карта-схема мест сбора изученного материала. 1 – Анадырский залив, окрестности города Анадырь, 2 – Олюторский залив, окрестности поселка Пахачи, 3 – залив Корфа, 4 – залив Анапка. 5 – Карагинский залив, бухта Оссора, 6, 7 – остров Карагинский, 8 – бухта Бечевинская, 9 – Авачинская губа, 10 – бухта Вилючинская, 11 – окрестности поселка Усть-Хайрюзово, 12 – остров Птичий, 13 – район между устьем реки Ковран и мысом Южный, 14 – окрестности поселка Палана. Условные обозначения: ● – места проведения альгопромысловых исследований, ▲ – места сбора альгофлористического материала.
Fig. 1. Map of sampling sites. 1 – Anadirskiy inlet, nearby Anadyr, 2 – Olyutorsky inlet, nearby village Pakhachi, 3 – Korfa inlet, 4 – Anapka inlet. 5 – Karaginsky inlet, Ossora bay, 6, 7 – Karaginskiy island, 8 – Bechevinskaya bay, 9 – Avacha bay, 10 – Vilyuchinskaya bay, 11 – coast nearby Ust-Khayryuzovo, 12 – Ptichij island, 13 – coast between estuary of Kovran river and Yuzhnyi cape, 14 – coast nearby Palana. ● – site of kelp resources studies, ▲ – sampling site of floristic materials.
В ходе альгопромысловых исследований у водорослей измеряли длину стволиков, длину и массу пластин. Для определения их средних показателей в разных районах побережья обычно измеряли 20–25 образцов из каждой возрастной группы. Большинство исследований было проведено в июле-августе.
Образцы для молекулярно-филогенетического анализа были собраны 9 июля 2018 г. в бухте Бечевинская (53°14'00'' N, 159°45'13'' E) на глубине 4 м. Это были слоевища второго года жизни, 1.3 м длины с достаточно тонкими вальковатыми стволиками, тонкими густо разветвленными ризоидами. Их пластинчатая часть имела округло-клиновидное основание, слабо волнистые края, более тонкие, чем широкая центральная полоса. В средней центральной части пластины были развиты хорошо видимые сорусы спорангиев.
Выделение ДНК, ПЦР-анализ, олигонуклеотидные праймеры и методы построения филогенетического дерева подробно описаны в наших статьях по ламинариевым водорослям российского Дальнего Востока (Klochkova et al., 2010; Klimova et al., 2018). Полученные последовательности зарегистрированы в базе данных NCBI (2022) под именем Saccharina latissima (L.) C.E. Lane, C. Mayes, Druehl et G.W. Saunders с присвоением следующих порядковых номеров: MH778276, MH778277 (COI, ваучерные образцы KamGTU_0013 и KamGTU_0014).
РЕЗУЛЬТАТЫ
Представители вида, относившегося нами ранее к L. gurjanovae f. lanciformis, L. lanciformis (Klochkova et al., 2009), Laminaria sp. avgub1 (Ko-neva, Klochkova, 2014), у берегов Камчатки распространены достаточно широко, но крайне неравномерно. В целом частота их встречаемости и ценотическая роль уменьшаются в направлении с севера на юг. У юго-восточной Камчатки вид встречается лишь на отдельных участках, близких к его эколого-ценотическому оптимуму. То же можно сказать и об особенностях его распределения у западного побережья полуострова. Во флоре Командорских островов данный вид не встречается. Его указания здесь как L. bullata Kjellm. (Kardakova-Prejentzoffa, 1938; Zinova, 1940) и L. gurjanovae (Ivanyushina et al., 1991) следует считать ошибочными. Это было показано в нашем обзоре бурых водорослей этого района, в том числе ламинариевых (Klochkova et al., 2020).
Массовое распространение обсуждаемый вид имеет на участке беринговоморского побережья, расположенном от зал. Анадырский до зал. Карагинский включительно. Здесь он является одним из доминантов подводной растительности. Типичным местом его произрастания являются участки побережья с пониженной прибойностью, расположенные в кутах мелководных заливов, обильно заселенных морской травой Zostera marina. В сообществе зостеры встречаются единично слоевища этого вида или их небольшие группы. В сторону от кутовой к мористой части он формирует более плотный покров и селится от сублиторальной каймы до нижней границы фитали. Его слоевища, прикрепленные к небольшим камням и гальке, приливами и волнами активно переносятся с одного места на другое и выбрасываются на берег. В местах, подверженных постоянному прибою, характер распределения вида меняется. Здесь на менее подвижных грунтах он формирует достаточно плотный растительный покров в среднем и нижнем горизонтах фитали.
Размерно-массовые показатели вида и его морфология в прикамчатской части ареала зависят от условий произрастания. Также для вида характерен широкий размах индивидуальной изменчивости. На форму слоевищ большое совокупное влияние оказывают возраст, фенофаза, прибойность и глубина произрастания. Различное сочетание этих факторов приводит к разным формообразовательным процессам. В результате чего в прикамчатских водах вид демонстрирует высокую изменчивость формы и размеров пластины, длины стволика, плотности ризоидальных пучков (рис. 2).
Рис. 2.
Морфология Saccharina latissima в прикамчатских водах. A – ювенилы из залива Анапка, B – слоевища первого года жизни из залива Корф, C, D – представители первого года жизни из Авачинской губы, E – разновозрастные образцы из бухты Бечевинская, F–G – образцы второго года жизни из Авачинской губы. Масштабные линейки: 20 см
Fig. 2. Morphology of Saccharina latissima from Kamchatka. A – juvenile specimens from Anapka inlet, B – 1st year old specimens from Korfa inlet, C, D – 1st year old specimens from Avacha bay, E – different samples from Bechevinskaya bay, F–G – 2nd year old specimens from Avacha bay.
A. Different samples collected from Avacha bay. B. Specimen with bullate blade from the tidal zone D. Branched haptera of young sporophyte. E. Group of algae with intertwined rhizoids. Scale bars: 20 cm.

Наши исследования показывают, что значительное гидродинамическое воздействие приводит к укорачиванию и суживанию пластин, уплотнению ризоидов. Цвет пластин и стволиков у представителей вида из прибойных мест обычно более темный. В местах со слабым волновым воздействием размеры слоевищ увеличиваются, ризоидальный пучок становится более рыхлым. Представленная ниже таблица (табл. 1) дает общее представление о размахе колебаний длины и массы пластин, а также длины стволиков у слоевищ, измеренных во время альгопромысловых съемок, проводившихся в разных районах камчатского побережья.
Таблица 1.
Размерно-массовые характеристики представителей Saccharina latissima у побережья Камчатки за период 2004–2020 гг. Table 1. Size and weight characteristics of Saccharina latissima samples from Kamchatka during 2004–2020
| Район Collection site |
Год иссле-дования Collection date |
Возраст, год Age, year |
Длина
черешка, см Stipe length, cm |
Длина
пластин, см Blade length, cm |
Масса пластин, г Blade wet weight, g |
Количество
образцов Number of samples |
|---|---|---|---|---|---|---|
| Залив Корфа, глубина 4–6 м Korfa inlet, depth 4–6 m |
май 2004 May 2004 |
1 | 16–22 | 67–135 | – | 28 |
| 2 | 19–28 | 80–220 | – | 22 | ||
| Залив Анапка, глубина 2.5 м Anapka inlet, depth 2.5 m |
май 2004 May 2004 |
1 | 13–18 | 138–183 | 136–151 | 45 |
| 2 | 16–22 | 156–240 | 233–267 | 31 | ||
| Остров Карагинский, глубина 8 м Karaginskiy island, depth 8 m |
август 2006 August 2006 |
2 | 22–38 | 296–430 | 940–1020 | 27 |
| Бухта Оссора, выбросы Ossora bay, cast ashore |
август 2006 August 2006 |
1 | 12–17 | 86–135 | – | 25 |
| 2 | 16– 24 | 142–176 | – | 29 | ||
| Бухта Бечевинская, глубина 8 м Bechevinskaya bay, depth 8 m |
август 2006 August 2006 |
1 | 18–21 | 130–267 | 217–242 | 19 |
| 2 | 21–26 | 320–468 | 788–831 | 33 | ||
| Авачинская губа, глубина 4 м Avacha bay, depth 4 m |
октябрь 2009; август 2013 October 2009, August 2013 |
1 | 18–24 | 96–124 | 112–137 | 41 |
| 2 | 22–31 | 138–196 | 142–347 | 127 | ||
| Бухта Вилючинская, глубина 6 м Vilyuchinskaya bay, depth 6 m |
июль 2014 July 2014 |
1 | 16–25 | 138–164 | – | 56 |
| 2 | 18–37 | 178–234 | – | 45 | ||
| Побережье у поселка Усть-Хайрюзово, выбросы Coast nearby Ust-Khayryuzovo, cast ashore | август 2020 August 2020 |
1 | 6–12 | 74–154 | 166–183 | 43 |
| 2 | 10–15 | 120–260 | 176–408 | 28 | ||
| Остров Птичий, глубокие литоральные ванны Ptichiy Island, pools of intertidal zone | август 2004, август 2020 August 2004, August 2020 |
1 | 12–16 | 138–145 | 172–186 | 32 |
| 2 | 15–23 | 161–240 | 438–465 | 21 |
Обсуждаемый вид у берегов Камчатки вегетирует два года. Максимальная длина его пластин у северо-восточной Камчатки достигает 430 см, масса может превышать 1 кг. В частности, такие размерно-массовые показатели были зарегистрированы у образцов, собранных у о. Карагинский. Ширина их пластин в конце августа достигала 42 см. У побережья острова, обращенного к проливу Литке, вид формирует монодоминантные сообщества. Они расположены ниже зоны разрушения крупных волн, возникающих во время сильных штормов, и тянутся вдоль всего юго-западного берега острова.
Южнее Карагинского залива вид сокращает свое присутствие. В заливе Озерной у открытых морских берегов он уже практически не встречается. Здесь доминируют представители родов Laminaria и Hedophyllum, имеющие рассеченные пластины. В заливах Камчатский и Кроноцкий он, судя по нашим наблюдениям, также не встречается. Не исключено его присутствие здесь в солоноватоводных озерах лагунного типа, расположенных вдоль Кроноцкого залива, но их альгофлора до сих пор не изучена.
В Авачинском заливе нам известно несколько мест произрастания вида. Здесь он встречается в бухтах фиордового типа Бечевинской (рис. 2 E) и Вилючинской, а также во внутренней части Авачинской губы, на участке побережья, расположенном от сопки Никольской до бухты Раковая включительно (рис. 2 C, D, F, G). В последней до конца прошлого столетия он формировал промысловое поле, простиравшееся между входными мысами. В настоящее время из-за сильного антропогенного загрязнения и заиления донных грунтов губы оно практически уничтожено. Большой ценотической роли в указанных районах юго-восточного побережья Камчатки вид не имеет. Южнее бухты Вилючинская вплоть до южной оконечности полуострова он, вероятно, отсутствует.
У юго-восточной Камчатки вид имеет два пика активного роста. Один приходится на позднюю весну, второй – на середину осени, но последний реализуется не столько в нарастание длины пластин, сколько в накопление массы. Самые крупные для юго-восточной Камчатки представители были собраны у южного входного мыса в бухту Бечевинская (2Е). Здесь на глубине 8 м средняя длина пластин у второгодних образцов достигала 462 ± 6.1 см, при максимальной ширине 27 см.
Вдоль южного берега бухты Вилючинская, самой южной из районов произрастания вида у восточной Камчатки, на глубинах 4–6 м он образует разреженное сообщество с плотностью 1–4 экз./м2. Здесь пластины второгодних образцов не превышают 234 см длины и 38 см ширины, при этом у слоевищ первого года жизни основание пластины широко клиновидное, края волнистые, були хорошо различимы по всей длине пластинчатой части слоевища. У зрелых второгодних образцов основание пластин почти округлое, боковые края ровные, були едва приметны в базальной части пластины или отсутствуют.
Изучение спороношения вида показывает, что у северо-восточной Камчатки из-за гораздо большей продолжительности длины дня зрелые сорусы спорангиев у обеих возрастных генераций имеются уже в мае. У представителей первого года жизни они обычно односторонние, второго – двусторонние. Они занимают центральную часть пластины, образуют широкую сплошную полосу или отдельные пятна, их очертания с двух сторон пластины не совпадают. В августе площадь сорусов спорангиев у представителей обеих возрастных групп увеличивается более чем в два раза. Изучить размножение вида в осеннее время в северных районах восточной Камчатки не представилось возможным.
У юго-восточной Камчатки вид в начале мая имеет еще незрелые сорусы спорангиев. Его спороношение здесь растянутое, с конца мая по ноябрь включительно, с двумя пиками наибольшей активности: поздневесенним и раннеосенним. Наибольший вклад в воспроизводство вида дает последний. В связи с постоянным более или менее активным выбросом зооспор разновозрастные проростки вида можно встретить в течение почти всего года.
У западной Камчатки в распространении вида просматривается та же тенденция, что и у восточного побережья: частота его встречаемости снижается с севера на юг. Точные данные о северной границе его распространения здесь отсутствуют, но, судя по всему, он встречается вплоть до кутовой части Пенжинского залива. Южной границей его распространения является мыс Амбон. На много километров к югу от него тянется пологий берег с песчаными грунтами, лишенными какой-либо подводной растительности. Твердые грунты замещают их только на самом юге западной Камчатки, в заливе Камбальном. Здесь господствуют ламинариевые беринговоморского флористического комплекса, среди которых обсуждаемый нами вид не встречается. Его размерно-массовые характеристики в центральных районах западной Камчатки приведены в таблице 1. По внешнему виду западнокамчатские образцы очень близки к представителям вида, произрастающим на северо-востоке полуострова. Проследить биологию развития вида и его размножение у западной Камчатки не удалось. На момент сбора в августе все образцы имели сорусы спорангиев.
Родственные филогенетические связи обсуждаемого вида с другими близкородственными ламинариевыми показаны на дереве, выстроенном по данным анализа гена СОI, как наиболее репрезентативного в филогении этого порядка (рис. 3). Для группы S. latissima мы выбрали нуклеотидные последовательности образцов этого вида из Европы, в том числе собранные в типовом местообитании, и образцы из США и Канады. Местом сбора североамериканских образцов является морское побережье штата Мэн, близкое к Атлантическому океану (KY319059) и арктическое побережье Канады – Гудзонов залив (FJ409199).
Рис. 3.
Молекулярно-филогенетическое дерево видов рода Saccharina, выстроенное методом байесовского выведения с использованием последовательностей субъединицы 1 цитохром c-оксидазы (COI). Значения напротив ветвей дерева – статистический бутстрэп/апостериорная вероятность сходства (учитывались только значения выше 75). Последовательности Saccharina latissima с юго-восточной Камчатки выделены жирным шрифтом. N – ветвь выстраивалась только в дереве MrBayes, при этом она отсутствовала в дереве RAxML. СН * – место сбора образца не указывалось в NCBI и других источниках. Показаны типичные морфологические особенности Saccharina cichorioides (A; образец из Японского моря) и S. latissima с юго-востока Камчатки (B; по Klochkova, Klochkova, 2018).
Fig. 3. Bayesian inference of phylogenies of Saccharina spp. based on cytochrome c-oxidase subunit 1 (COI) sequences. Consensus support (%) and posterior probabilities are shown (only the values above 75 were included). Sequences of Saccharina latissima from Southeast Kamchatka are highlighted in bold. N – branch was present only in the MrBayes tree, and it was absent in the RAxML tree. CH * – collected site of sample was not indicated in the NCBI and other sources. Specimens with typical morphology of Saccharina cichorioides from the Sea of Japan (A) and S. latissima from Southeast Kamchatka (B; by Klochkova, Klochkova, 2018).

ОБСУЖДЕНИЕ
Представители родов Laminaria и Saccharina в морфологическом отношении делятся на две группы. У одних пластины с возрастом рассекаются на продольные лопасти, у других они в любом возрасте остаются цельными. В камчатской альгофлоре цельнолистные пластины характерны только для двух видов. Один из них, L. longipes Bory, характеризуется наличием стелющихся ризомов и узколинейных пластин с одной или двумя перетяжками. Его трудно спутать с любым другим представителем камчатских ламинариевых. Второй цельнолистный вид разные исследователи, как было упомянуто выше, определяли по-разному.
В альгофлоре северо-западной Пацифики впервые этот цельнолистный вид был указан для Охотского моря еще Ф.И. Рупрехтом (Ruprecht, 1851), как L. saccharina. Это название он носил до тех пор, пока А.Д. Зинова не описала для российского Дальнего Востока новый для науки вид L. gurjanovae (Zinova A., 1964, 1969), полагая, что он отличается от L. saccharina наличием более темного и толстого, чем остальная часть пластины, длинного узкоклиновидного основания, составляющего 25–30% общей ее длины. Интересной особенностью описанного ею вида были его разорванный ареал, а также приуроченность к большим глубинам и районам с высокой подвижностью воды.
Ю.Е. Петров (Petrov, 1972), проведя ревизию рода Laminaria в морях российского Дальнего Востока, расширил диагноз L. gurjanovae. Кроме типовой формы он отнес к ней образцы, ранее определявшиеся в альгофлоре этого района как L. saccharina. Он выделил их в отдельную форму L. gurjanovae f. lanciformis. Мнение Ю.Е. Петрова о том, что L. saccharina в приазиатских водах не встречается, стало общепризнанным среди российских альгологов.
В 2006 г. по данным секвенирования хлоропластной (Rubisco operon), митохондриальной (nad6) и ядерной (ITS и LSU rDNA) ДНК род Laminaria был разделен на роды Laminaria и Saccharina (Lane et al., 2006). Вид L. saccharina попал в кладу Saccharina, был признан типом этого рода и ему был возвращен видовой эпитет “latissima”, данный еще К. Линнеем (Linnaeus, 1753). Название L. saccharina, таким образом, стало синонимом Saccharina latissima. Ламинариевые дальневосточных морей России, в том числе образцы L. gurjanovae, собранные в типовым местообитании, у южного Сахалина, к вышеупомянутому исследованию канадских ученых не привлекались. С учетом предложенного К. Лэйном с соавторами разделения рода Laminaria таксономическое положение этого и других приазиатских видов ламинарий требовало уточнения. Позднее в 2007 г. О.Н. Селиванова с соавторами отнесла обе формы L. gurjanovae к роду Saccharina (Selivanova et al., 2007). Детально эти номенклатурные изменения обсуждены в наших публикациях (Koneva, Klochkova, 2014; Klochkova, Klochkova, 2018).
Принадлежность камчатской цельнолистной ланцетовидной ламинарии к роду Saccharina была установлена нами по результатам секвенирования гена СОI. Молекулярно-филогенетический анализ показал совпадение всех последовательностей S. latissima в группе образцов, собранных в разных районах атлантического побережья Европы. Разница между ними по гену COI колеблется от 99.5 до 99.7%; между европейскими и канадскими образцами из Гудзонового залива она составляет 98.9%; между камчатскими и европейскими – 98.7–99%; между камчатскими и канадскими – 99.8% (рис. 3). Столь высокий уровень генетического сходства камчатских образцов, особенно с представителями арктического побережья Канады, является свидетельством их принадлежности к S. latissima. Это подтверждается также высоким морфологическим сходством представителей атлантической и тихоокеанской популяций этого вида. Следуя принятой среди альгологов интерпретации молекулярно-генетических данных (McDevit, Saunders, 2010), мы полагаем, что различие 1.3% между последовательностями камчатской и европейской S. latissima следует рассматривать как межпопуляционное и недостаточное для отнесения камчатских и североамериканских образцов из Гудзонового залива к самостоятельному, отличному от S. latissima, виду или его форме.
В отношении последовательностей S. angustata (Kjellm.) C.E. Lane, C. Mayes, Druehl et G.W. Saunders, S. japonica (Aresch.) C.E. Lane, C. Mayes, Druehl et G.W. Saunders, S. longissima (Miyabe) C.E. Lane, C. Mayes, Druehl et G.W. Saunders, S. religiosa (Miyabe) C.E. Lane, C. Mayes, Druehl et G.W. Saunders и S. ochotensis (Miyabe) C.E. Lane, C. Mayes, Druehl et G.W. Saunders, использовавшихся для филогенетического анализа, ситуация довольно странная. Исходя из того, что совпадение всех последовательностей в этой группе равно 100%, их следовало бы объединить в один вид. Детальное изучение представленных в NCBI (NCBI, 2021) данных по гену COI (JX442493–JX442496) позволяет предположить, что в определении указанных выше видов (Liu et al., 2014) были допущены ошибки. Проверить это сейчас достаточно сложно, поскольку в указанной работе не приведены фотографии ваучерных образцов, использовавшихся для молекулярно-филогенетического анализа, и не указаны места их сбора. В процитированной выше работе нет ссылок на статьи Йотсукуры с соавторами, в которых на основании филогенетического анализа показано, что виды S. religiosa и S. ochotensis являются формами S. japonica, а S. angustata и S. longissima – самостоятельными видами (Yotsukura et al., 2008, 2010; Zhang et al., 2019).
В построенном нами молекулярном дереве совпадение последовательностей S. groenlandica (Rosenv.) C.E. Lane, C. Mayes, Druehl et G.W. Saunders (GU097739) с камчатскими образцами Hedophyllum bongardianum (Postels et Rupr.) Yendo (GU097729) по гену COI составляет 98.2%. Этого достаточно для сведения S. groenlandica в синонимы Н. bongardianum, но только в том случае, если определение образца S. groenlandica, зарегистрированного в Генбанке под номером GU097739, было правильным. Между тем, согласно данным AlgaeBase (Guiry, Guiry, 2022), S. groenlandica является синонимом S. latissimа, хотя различия по гену COI между этими видами составляют 4.5–5.3%.
Выше уже отмечался высокий уровень генетического сходства между представителями камчатской и канадской популяций S. latissima из атлантического сектора Арктики. Для объяснения этого, безусловно, необходимо обращение к геологической истории арктического региона и наличие молекулярно-генетических данных из других приполярных районов Тихого океана и прилегающего к нему сектора Арктики. Такие данные для S. latissima пока отсутствуют, поэтому мы оставляем этот вопрос открытым.
ЗАКЛЮЧЕНИЕ
Проведенное нами исследование позволило определить таксономическое положение цельнолистного представителя рода Saccharina, распространенного вдоль берегов Камчатки, и показать, что он принадлежит широко распространенному в Северном полушарии виду S. latissima. Все названия, под которыми он указывался во флоре Камчатки (L. gurjanovae f. lanciformis, L. lanciformis, Laminaria sp. avgub 1, Saccharina gurjanovae), являются синонимами названия Saccharina latissima.
В отношении типовой формы L. gurjanovae, имеющей очень специфическое морфологическое строение, вопрос остается открытым. Для того чтобы определить, является она самостоятельным видом или формой вида S. latissima, необходимо проведение специального молекулярно-генетического анализа с обязательным включением в него образцов из типового местообитания вида, залива Анива (Охотском море, о. Сахалин).
Список литературы
[Belij] Белый М.Н. 2013. Водоросли-макрофиты северной части Охотского моря и их значение как нерестового субстрата сельди. Магадан. 194 с.
Climate Change 2013: The Physical Science Basis. Contribution of Working Group I to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change. 2013. Cambridge, New York. 1535 p.
[Gajl] Гайл Г.И. 1936. Ламинариевые водоросли дальневосточных морей. – Вестник ДВФ АН СССР. 19: 31–65.
Guiry M.D., Guiry G.M. 2022. AlgaeBase. World-wide electronic publication, National University of Ireland, Galway. http://www.algaebase.org. (Accessed 19.03.2022).
[Ivanyushina et al.] Иванюшина Е.А., Ржавский А.В., Селиванова О.Н., Ошурков В.В. 1991. Структура и распределение сообществ бентоса мелководий Командорских островов. – В кн.: Природные ресурсы Командорских островов (снабжение, состояние, вопросы охраны и использования). М. С. 155–170.
[Kardakova–Prejentzoffa] Кардакова–Преженцова Е.А. 1938. Водорослевая растительность Командорских островов. – Известия ТИНРО. 14: 77–108.
Klimova A.V., Klochkova N.G., Klochkova T.A., Kim G.H. 2018. Morphological and molecular identification of Alaria paradisea (Phaeophyceae, Laminariales) from the Kurile Islands. – Algae. 33 (1): 37–48.
[Klochkova et al.] Клочкова Н.Г., Королева Т.Н., Кусиди А.Э. 2009. Атлас водорослей-макрофитов прикамчатских вод. Т. 1. Зеленые и бурые водоросли. Петропавловск-Камчатский. 218 с.
Klochkova T.A., Kim G.H., Lee K.M., Choi H.-G., Belij M.N., Klochkova N.G. 2010. Brown algae (Phaeophyceae) from Russian Far Eastern seas: re-evaluation of Laminaria multiplicata Petrov et Suchovejeva. – Algae. 25: 77–87.
[Klochkova et al.] Клочкова Н.Г., Клочкова Т.А., Климова А.В. 2020. Флора водорослей-макрофитов Командорских островов: ревизия 2020. I. Chlorophyta, Ochrophyta. – Вестник КамчатГТУ. 54: 82–107.
[Klochkova, Klochkova] Клочкова Т.А., Клочкова Н.Г. 2018. Проблемы геносистематики и изменение родовой принадлежности и авторства у видов бурых ламинариевых водорослей Saccharina bongardiana и Saccharina gurjanovae. – Вестник КамчатГТУ. 43: 87–95.
[Koneva, Klochkova] Конева А.А., Клочкова Н.Г. 2014. Сезонная динамика общего химического состава у Laminaria sp. (avb_1) (Laminariales, Рhaeophyta), произрастающей в загрязненных районах Авачинской губы (юго-восточная Камчатка). – Исследования водных биологических ресурсов Камчатки и северо-западной части Тихого океана. 33: 78–86.
Lane C.E., Mayes C., Druehl L.D., Saunders G.W. 2006. A multi-gene molecular investigation of the kelp (Laminariales, Phaeophyceae) supports substantial taxonomic re-organization. – Journal of Phycology. 42: 493–512.
Linnaeus C. 1753. Species plantarum, exhibentes plantas rite cognitas, ad genera relatas, cum differentiis specificis, nominibus trivialibus, synonymis selectis, locis natalibus, secundum systema sexuale digestas. Holmiae. Stockholm. 2: 561–1200.
Liu et al. 2014. A comprehensive phylogeny of Laminariales (Phaeophyceae) based on mitochondrial-encoded Cytochrome Oxidase Subunit I, plastid-encoded Rubisco Large Subunit and nuclear-encoded ITS sequence comparisons (Unpublished record, only present in NCBI).
McDevit D.C., Saunders G.W. 2010. A DNA barcode examination of the Laminariaceae (Phaeophyceae) in Canada reveals novel biogeographical and evolutionary insights. – Phycologia. 49: 235–248.
National Center for Biotechnology Information (NCBI). 2022. GenBank. http//www.ncbi.nlm. nih.gov. (Accessed 19.03.2022).
[Petrov] Петров Ю.Е. 1972. Систематика некоторых дальневосточных видов рода Laminaria Lamour. – Новости сист. низш. раст. 9: 47–58.
Ruprecht F.I. 1851. Tange des Ochtskischen Meeres. – Sibirishe Reise. Botanik. St.-Peterburg. 1 (2): 191–435.
[Selivanova et al.] Селиванова О.Н., Жигадлова Г.Г., Хэнсен Г.И. 2007. Пересмотр систематики водорослей порядка Laminariales (Phaeophyta) в дальневосточных морях России на основании молекулярно-генетических данных. – Биология моря. 33 (5): 329–340.
[Vinogradova et al.] Виноградова К.Л., Клочкова Н.Г., Перестенко Л.П. 1978. Список водорослей литорали восточной Камчатки и западной части Берингова моря – В кн.: Литораль Берингова моря и юго-восточной Камчатки. М. С. 150–155.
[Vtoroy…] Второй оценочный доклад Росгидромета об изменениях климата и их последствиях на территории Российской Федерации. 2014. М. 1003 с.
Yotsukura N., Kawashima S., Kawai T., Abe T., Dreuhl L.D. 2008. A systematic re-examination of four Laminaria species: L. japonica, L. religiosa, L. ochotensis and L. diabolica. – Journal of Japanese Botany. 83: 165–176.
Yotsukura N., Shimizu T., Katayama T., Druehl L.D. 2010. Mitochondrial DNA sequence variation of four Saccharina species (Laminariales, Phaeophyceae) growing in Japan. – J. Appl. Phycol. 22: 243–251.
Zhang J., Wang X., Yao J., Yotsukura N., Duan D. 2019. Screening of polymorphic microsatellites and their application for Saccharina angustata and Saccharina longissima population genetic analysis. – J. Appl. Phycol. 31: 3295–3301.
Zhang J., Yotsukura N., Jueterbock A., Hu Z.-M., Assis J., Nagasato C., Yao J., Duan D. 2021. Detecting no natural hybridization and predicting range overlap in Saccharina angustata and Saccharina japonica. – J. Appl. Phycol. 33: 693–702.
[Zinova] Зинова Е.С. 1933. Водоросли Камчатки. – Исследования морей СССР. 17: 7–42.
[Zinova] Зинова Е.С. 1940. Морские водоросли Командорских островов. – Научные труды Тихоокеанского комитета. 5: 165–243.
[Zinova] Зинова Е.С. 1954a. Водоросли Охотского моря. – Тр. Ботан. ин-та АН СССР. 9: 259–307.
[Zinova] Зинова Е.С. 1954b. Морские водоросли юго-восточной Камчатки. – Тр. Ботан. ин-та АН СССР. 9: 365–400.
[Zinova A.] Зинова А.Д. 1964. Новый вид Laminaria у берегов Сахалина. – Новости сист. низш. раст. 1: 125–126.
[Zinova A.] Зинова А.Д. 1969. Дополнение к статье о новом виде ламинарии с острова Сахалин. – Новости сист. низш. раст. 6: 65–68.
Дополнительные материалы отсутствуют.
Инструменты
Ботанический журнал