

Физиология человека, 2019, T. 45, № 2, стр. 16-28
Интракортикальные связи при выполнении двойных задач – моторных и счетно-логических или пространственно-образных
Л. А. Жаворонкова 1, *, Т. П. Шевцова 2, С. И. Морареску 2, А. В. Позднеев 1, С. В. Купцова 1, 3
1 ФГБУН Институт высшей нервной деятельности и нейрофизиологии РАН
Москва, Россия
2 Московский государственный университет им. М.В. Ломоносова
Москва, Россия
3 ГБУЗ Центр патологии речи и нейрореабилитации ДМЗ
Москва, Россия
* E-mail: Lzhavoronkova@hotmail.com
Поступила в редакцию 12.12.2017
После доработки 10.07.2018
Принята к публикации 01.11.2018
Аннотация
Комплексное электроэнцефалографическое (ЭЭГ) и стабилографическое исследование проведено у 19 здоровых испытуемых (25 ± 0.7 лет) при выполнении изолированных и двойных задач – когнитивных и моторных. Изолированное выполнение счетно-логических задач сопровождалось увеличением когерентности медленных ритмов ЭЭГ – ∆–θ-диапазонов преимущественно в лобно-височных отделах левого полушария. При выполнении пространственно-образных задач наблюдалось дополнительное увеличение когерентности медленных ритмов ЭЭГ и в правом полушарии. Выполнение моторного задания сопровождалось увеличением когерентности частых ритмов ЭЭГ (α–β–γ) в правом полушарии. При выполнении двойных задач наблюдалось снижение числа связей по медленным ритмам по сравнению с количеством связей при изолированном выполнении заданий. Наряду с этим решение двойных задач сопровождалось нарастанием когерентности ритмов в α1-частотном диапазоне для лобных, центральных и теменных отделов обоих полушарий. Это может быть отражением распределения когнитивных ресурсов на выполнение обоих компонентов двойных задач, а также использованием дополнительных мозговых резервов в этих условиях.
В повседневной жизни люди часто сталкиваются с необходимостью одновременно решать две или несколько задач. Известно, что человек может одновременно управлять машиной и разговаривать по телефону или идти и слушать музыку или разговаривать с собеседником. В исследованиях, посвященных анализу специфики когнитивных процессов при одновременном выполнении разных видов деятельности, такие ситуации называют двойными задачами (dual-task). Чаще всего в качестве компонентов двойных задач используются моторные и когнитивные задания [1–4]. В клинических исследованиях в качестве моторных заданий, как правило, используются ходьба или произвольное поддержание определенной позы, которые часто бывают нарушенными в результате церебральной патологии [5–7]. В последние годы значительно возрос научный интерес к данной проблеме, и увеличилось количество специальных исследований, направленных на изучение мозговых механизмов выполнения двойных задач. В таких исследованиях в качестве когнитивных задач используются различные задания – мысленный счет, вербально-логические, зрительно-пространственные задачи, звуковые стимулы и пр., а также нередко две когнитивные или две моторные задачи [8–10].
На начальных этапах исследования двойных задач анализировались поведенческие данные, в которых акцентировалось внимание на более низком качестве выполнения каждого из компонентов двойных задач, по сравнению с изолированными заданиями [3–5 и др.]. Несмотря на кажущуюся легкость одновременного выполнения двух задач, качество позы или ходьбы здорового человека обычно замедляется в большей или меньшей степени и зависит от сложности когнитивных задач, а также в случае усложнения условий поддержание позы или ходьбы, когда они перестают выполняться автоматически [8]. Эти результаты объясняли ограничениями когнитивных ресурсов, в частности, распределением внимания между двумя задачами и развитием процессов интерференции [9–11]. Именно поэтому в подавляющем большинстве работ акцентируется внимание на преимущественном ухудшении качества выполнения именно когнитивного компонента при сохранении качества (приоритете) моторного задания [2, 11, 12].
Моторные функции, наиболее древние в филогенетическом плане и биологически важные для человека, как правило, являются приоритетными при решении двойных задач, что особенно отчетливо проявляется у пациентов с разными формами церебральной патологии [13]. Обычно пациенты выполняют компоненты двойных задач не одновременно, а последовательно, при приоритете выполнения моторного задания [2, 11, 12].
Несмотря на то, что здоровые люди также чаще выполняют двойные задачи с худшим качеством, чем отдельно каждую из задач, встречаются ситуации, когда двойные задачи выполняются более успешно, чем изолированные [14]. Известно также, что в экстремальных ситуациях некоторые люди могут осуществить сверхсложные задачи, которые они не в состоянии выполнить в обычных условиях. Профессиональные спортсмены, для которых специализированные двигательные нагрузки приобретают признаки автоматизма, могут демонстрировать более высокое качество выполнения двойных задач по сравнению с изолированными [14].
Успешность выполнения двойных задач в большой степени зависит от индивидуальных психологических особенностей испытуемых, включающих ресурсы памяти и внимания, а также скорости его переключения с одного задания на другое [15]. Совокупность всех перечисленных факторов позволяет одновременно контролировать двигательные и когнитивные процессы. Качество выполнения двойных задач в большой мере зависит от возраста испытуемых – способность успешно выполнять двойные задачи снижается по мере увеличения возраста человека [16–18].
Существует несколько гипотез, объясняющих механизмы, лежащие в основе возможности выполнения двойных задач. Согласно одной точке зрения, успешность выполнения двойных задач объясняется вовлечением дополнительных областей мозга при их выполнении по сравнению с изолированными [19]. Помимо этого высказывается мнение о возможности параллельного выполнения двух задач [20]. И, наконец, существует гипотеза, согласно которой успешное выполнение двойных задач обусловлено не только вовлечением разных областей мозга, обеспечивающих осуществление каждого из компонентов двойной задачи, а также разными временны́ми параметрами выполнения каждого из заданий [21]. Наши предыдущие исследования в большей степени согласуются с последней гипотезой [22–26].
При анализе механизмов, обеспечивающих успешное выполнение двойных задач, большая часть исследователей фокусирует внимание на активации фронтальной и париетально-окципитальной коры как ключевом элементе выполнения двойных задач [27]. В наших исследованиях были также получены сведения о том, что успешность выполнения двойных задач обеспечивается “включением” диагональных функциональных связей между ассоциативными отделами мозга – левой лобной и правой теменной, а также правой лобной и левой теменной областями [23].
Следует подчеркнуть, что изучение нейрональных механизмов, обеспечивающих осуществление двойных задач, стало возможным только в последние годы, когда появилось телеметрическое оборудование, позволяющее регистрировать биоэлектрическую активность мозга человека on-line – в процессе выполнения различных видов движений – ходьбе, поддержании или изменении позы на стабилографической платформе и др. [22–32]. В этих работах используются различные подходы – электроэнцефалография (ЭЭГ), вызванные потенциалы и другие методы [22–32]. Анализ публикаций с использованием различных методов регистрации биоэлектрической активности, а также видов их анализа – оценки мощности различных ритмов ЭЭГ, функциональных связи и др., свидетельствует об отсутствии однозначных данных об особенностях реактивных перестроек при выполнении когнитивных заданий изолированно и в составе двойных задач [28–32]. Наиболее часто описываются реактивные перестройки во фронтальных и префронтальных отделах мозга преимущественно в θ-диапазоне [28–33].
Использование когерентности ЭЭГ для оценки реактивных перестроек активности мозга человека, отражающей функциональные корково-корковые связи, продемонстрировало высокую информативность данного подхода в норме и при разных формах церебральной патологии [34–39].
При анализе изолированных и двойных задач с использованием мысленного счета для активации когнитивного компонента деятельности были выявлены реактивные перестройки когерентности θ–∆-диапазонов ЭЭГ в лобно-височных областях преимущественно левого полушария [22, 23]. В отличие от этого, выполнение моторных задач (поддержание или изменение позы) сопровождалось увеличением когерентности высокочастотных составляющих ЭЭГ преимущественно в правом полушарии [22–26]. В последующем сходные реактивные перестройки ЭЭГ при произвольном поддержании позы были подтверждены и другими авторами [29].
Несмотря на большой интерес к проблеме изучения мозговых механизмов выполнения двойных задач, данные об особенностях функционирования мозга человека в этих условиях трудно сопоставлять, так как результаты исследований получены в разных экспериментальных ситуациях и с использованием разных методических подходов [5, 7, 12]. Несмотря на появление современных технологий, которые позволили перейти на уровень изучения нейрональных механизмов работы мозга в условиях выполнения двойных задач, многие аспекты этого направления исследований остаются недостаточно изученными. В определенной степени это связано с характером когнитивных и моторных задач.
Выбор моторных заданий, включающих поддержание или изменение вертикальной позы, был в первую очередь обусловлен клиническими, реабилитационными задачами. В реабилитационные программы нейрохирургических пациентов, в частности, с черепно-мозговой травмой (ЧМТ), как правило, включается тренинг позного контроля – стабилотренинг. Это объясняется тем, что у пациентов с последствиями ЧМТ, перенесшими длительную потерю сознания, нарушается способность самостоятельно стоять и ходить. Было выявлено положительное влияние стабилотренинга на восстановление не только моторных, но и когнитивных функций, даже у пациентов с посттравматическим Корсаковским синдромом [25, 26]. Эти факты явились определяющими для дальнейшего изучения особенностей выполнения двойных задач, включающих произвольный позный контроль с одновременным выполнением когнитивных задач у здоровых людей и при реабилитации пациентов с последствиями ЧМТ [22–26].
При выборе когнитивных задач мы использовали мысленный счет, при выполнении которого наблюдалась преимущественное увеличение функциональных связей в лобно-височных отделах левого полушария [22, 23]. Однако в повседневной жизни здоровый человек и пациент сталкивается и с так называемыми “правополушарными” задачами – необходимостью ориентироваться в окружающей обстановке и пр. Известно, что у пациентов с преимущественной дисфункцией теменно-затылочных отделов правого полушария в результате повреждений разного генеза, нарушается способность ориентироваться в пространстве (анозогнозия) [13 и др.]. В связи с этим возникает вопрос о мозговых механизмах, обеспечивающих выполнение изолированных и двойных задач, включающих так называемые “правополушарные” и “левополушарные” задачи, например, при выполнении слуховых и зрительных заданий. Однако особенности латеральных перестроек ЭЭГ в настоящее время в таких условиях изучены недостаточно [32, 33]. Именно поэтому задачей данного исследования явилось сопоставление реактивных перестроек когерентности ЭЭГ при выполнении изолированных задач, включающих латерализованные – счетно-логические и пространственно-образные задания, а также двойных задач, включающих дополнительно моторное задание – произвольное поддержание позы.
МЕТОДИКА
В исследовании принимали участие 19 здоровых испытуемых (14 мужчин и 5 женщин) – правшей (25 ± 0.7 лет). Все обследованные испытуемые не имели в анамнезе неврологических заболеваний и дали письменное согласие на участие в исследовании в соответствии с положением Хельсинкского соглашения.
Задания. В данном исследовании использовали 1 моторную и 3 когнитивные задачи (рис. 1). Для осуществления моторной задачи использовали стабилограф Стабилан 0.1 (Таганрог, Россия). При выполнении задания испытуемому предлагали, стоя на стабилоплатформе, поддерживать равновесие с поддержанием общего центра давления (ОЦД) в одном и том же положении. На экране монитора ОЦД испытуемого представлен в виде крестика, который необходимо совмещать с маркером – кружком, находящемся в центре монитора при использовании обратной зрительной связи (рис. 1).
Рис. 1.
Схема проведения эксперимента. А – пример выполнения двойной задачи С2М1, включающей вербально-логическую задачу С2 и моторную задачу М1; Б – пример выполнения двойной задачи С3М1, включающей пространственно-образную задачу С3 и моторную задачу М1.

При выполнении первой когнитивной задачи (С1) испытуемому предлагали прослушать ряд высоких (800 Гц) и низких звуков (400 Гц), длительностью 80 мс каждый и одинаковых по громкости (75 дБ), которые предъявляли в случайном порядке с изменяющимся интервалом. Количество низких звуков было меньше, чем высоких – 40–45% из общего числа представленных. Испытуемому необходимо было подсчитать в уме количество предъявленных низких звуков.
При выполнении второй когнитивной задачи (С2) испытуемому также через слуховой анализатор в случайном порядке перечисляли предметы, относящиеся к категории “одежда” или “еда” (все слова состояли из двух слогов). Он должен был подсчитывать количество предметов, относящихся к одной из категорий, указанных экспериментатором, которые также составляли 40% от общего числа перечисленных предметов (рис. 1, А).
При выполнении третьей задачи (С3) испытуемому называли время, например 7.15. Он должен был мысленно представить себе циферблат часов и расположение на нем часовых стрелок, соответствующее названному времени. В том случае, если обе стрелки (минутная и часовая) находились в одной и той же половине циферблата, он говорил “да”. Если же стрелки находились в разных половинах циферблата, он молчал (рис. 1, Б).
Стимулы во всех трех задачах предъявляли с меняющимся интервалом в пределах 1.7–2.0 с. Для каждой когнитивной задачи подготовили 5 вариантов, которые предъявляли в случайном порядке. Качество выполнения когнитивных задач оценивали по количеству выполненных за минуту счетных операций с учетом количества допущенных ошибок. Для определения сложности каждого из заданий использовали результаты самоотчета испытуемых. Сложность задач была оценена испытуемыми следующим образом – 85% оценили задачу С3 как наиболее сложную. При сравнении задач С1 и С2 большая часть испытуемых (65%) оценили задание С1 как более сложное.
Процедура эксперимента. Перед проведением исследования экспериментатор рассказывал испытуемому, какие задания ему будет необходимо выполнять, и испытуемый прослушивал все варианты когнитивных заданий. После этого на нем закрепляли телеметрическое оборудование, включающее электродную шапочку и он, стоя на полу с открытыми глазами (перед стеной), выполнял все когнитивные задачи, варианты которых предъявляли в случайном порядке. Сначала осуществляли пробное выполнение задания, после чего начинали регистрацию ЭЭГ при выполнении другого варианта когнитивного задания. На следующем этапе исследования у испытуемого, стоящего на стабилоплатформе, осуществляли регистрацию ЭЭГ с открытыми глазами без каких-либо инструкций, а после этого он выполнял моторную задачу М1. После выполнения всех изолированных задач испытуемый выполнял двойные задачи – М1С1, М1С2 и М1С3, которые также предъявляли в случайном порядке. Каждое задание длилось 60 с. Качество выполнения когнитивных заданий вычисляли по количеству правильных ответов, а моторных – в процентах, которые вычислялись автоматически с помощью стабилографического оборудования и отражали время нахождения ОЦД испытуемого в заданных координатах.
Регистрацию телеметрической (беспроводной) монополярной ЭЭГ (Медикор, Таганрог, Россия) осуществляли от 19 областей коры при расположении электродов по международной системе 10–20 с частотой опроса 200 Гц в состоянии покоя с открытыми глазами и во время перечисленных заданий. Артефакты удаляли вручную, после чего проводили спектрально-когерентный анализ ЭЭГ. В дальнейший анализ включили эпохи с длительность отрезков 45–50 с. Спектры когерентности ЭЭГ вычисляли с шагом 0.4 Гц в частотной полосе 0.5–50 Гц для следующих диапазонов ритмов ЭЭГ: ∆ – 1.2–3.9 Гц, θ – 4.3–7.8 Гц, α1 – 8.2–10.2 Гц, α2 – 10.5–12.9, β – 13.3–30.1 и γ – 30.5–40.2 Гц. Анализировали не менее 9 эпох с перекрытием длительностью 5.12 с каждая. Когерентные индикаторы вычисляли как усредненные для каждого частотного диапазона значения квадратов модулей комплексной спектральной когерентности с использованием программы MatLab 6.0 фирмы Math Works Inc. Статистический анализ когерентности ЭЭГ проводили для парных выборок с распределением, отклоняющимся от нормального и использованием непараметрического критерия Вилкоксона для связанных выборок, как было описано ранее [22, 23].
РЕЗУЛЬТАТЫ ИССЛЕДОВАНИЯ
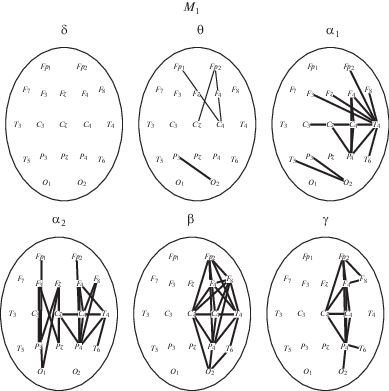
Анализ реактивных перестроек когерентности в выделенных частотных диапазонах ЭЭГ при выполнении моторной задачи представлен на рис. 2. Из рис. видно, что наиболее отчетливое увеличение когерентности наблюдается для α-, β- и γ-диапазонов, преимущественно в лобно-центрально-теменных отделах правого полушария, а также затылочных областях.
Рис. 2.
Особенности изменений когерентности для выделенных диапазонов ритмов ЭЭГ при изолированном выполнении моторного задания М1 по сравнению с состоянием покоя (стояние на стабилоплатформе с открытыми глазами) для значимых (p < 0.01) изменений когерентности ЭЭГ. Значимость различий указана на уровне парных сравнений. Толстые линии – увеличение когерентности, тонкие – снижение.
Результаты анализа изменений когерентности ЭЭГ при изолированном выполнении всех когнитивных задач представлены на рис. 3. По данным рис. 3 видно, что при выполнении двух первых когнитивных, счетно-логических задач – С1 и С2 выявлено значимое увеличение когерентности медленных ритмов ЭЭГ, преимущественно дельта-диапазона в лобно-височных отделах левого полушария. Несмотря на сходство реактивных перестроек ЭЭГ для двух первых задач, можно отметить небольшое превышение числа пар отведений в лобно-височных областях для С1 по сравнению с задачей С2. Наблюдаемое увеличение когерентности в лобно-центральных отделах правого полушария сходное с таковым при выполнении моторной задачи М1, но менее выраженное, что может быть связано с непроизвольным поддержанием вертикальной позы, так как испытуемые выполняли когнитивные задачи стоя. Возможно, что задача С1, часто воспринимая испытуемыми как более сложная, чем С2, требовала больших общих усилий.
Рис. 3.
Особенности изменений когерентности для выделенных диапазонов ритмов ЭЭГ при изолированном выполнении когнитивных задач: счетно-логических задач С1 и С2, а также пространственно-образной задачи С3 по сравнению с состоянием покоя (стояние на стабилоплатформе с открытыми глазами) для значимых (p < 0.01) изменений когерентности ЭЭГ. Остальные обозначения см. рис. 2.
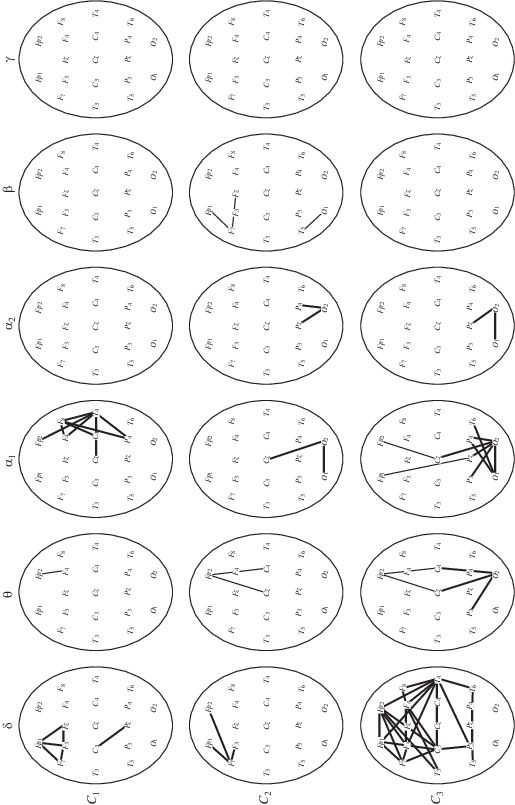
При выполнении пространственно-образной задачи С3 наблюдалось увеличение числа связей ∆-диапазона как в левом, так и в правом полушарии, со смещением фокуса в центральные и теменные области. Помимо этого наблюдалось увеличение числа связей θ- и α1-диапазона в затылочно-теменных отделах, наблюдающееся при выполнении задания С2 и наиболее выраженное для задачи С3. Можно полагать, что это связано с активацией париетально-окципитальной коры при обработке зрительно-пространственной информации при мысленном представлении условий заданий. Выполнение данной задачи сопровождалось увеличением когерентности медленных ритмов ЭЭГ в большем числе пар отведений, по сравнению с двумя предыдущими задачами. В соответствии с данными самоотчета задача С3 для большинства испытуемых была более сложной, по сравнению с задачами С1 и С2.
Выполнение двойных задач, включающих все когнитивные задания – С1, С2, и С3 в сочетании с выполнением моторной задачи, представлено на рис. 4. На рис. 4 видно, что при выполнении двойных задач, включающих задания С1 и С2 наблюдается снижение числа связей по сравнению с изолированными задачами. Исключение составляет задача М1С3. Обращает на себя внимание увеличение числа связей α1-диапазона, характерное для всех вариантов двойных задач. Нарастанием когерентности α1-диапазона наблюдается в лобно-центральных и теменно-затылочных областях с акцентом слева для двойной задачи, включающей задание С1, а при выполнении задач С2 и С3, связанных с представлением образов – преимущественно в правом полушарии. Эти данные дополнительно представлены в табл. 1 с количественной оценкой увеличения (+) числа связей или уменьшения (–) в двойной задаче, по сравнению с изолированной в соответствии с подходом, предложенным другими авторами [27]. Результаты анализа поведенческих данных свидетельствуют о незначительном снижении качества выполнения двойных задач по сравнению с изолированными: среднее количество выполненных изолированных заданий – С1 – 17 ± 2%; C2 18 ± ± 1%; C3 – 17 ± 3%; M1 – 98 ± 2% и двойных задач – М1С1 – (C1 – 16 ± 3%; M1 – 96 ± 3%); M1C2 – (C2 – 16 ± 3%; M1 – 97 ± 3%; M1C3 – (C3 – 16 ± 3%; M1 – 95 ± 4%).
Рис. 4.
Особенности изменений когерентности ЭЭГ при выполнении двойных задач (С1М1, С2М1 и С3М1), включающих выполнение счетно-логических задач (С1 и С2), а также пространственно-образной задачи (С3) одновременно с моторной задачей М1 по сравнению с состоянием покоя (стояние на стабилоплатформе с открытыми глазами) для значимых (p < 0.01) изменений когерентности ЭЭГ. Остальные обозначения см. рис. 2.

Таблица 1.
Сравнение количества функциональных связей при выполнении двойных задач по сравнению с изолированными для различных областей мозга и диапазонов ритмов ЭЭГ (+ увеличение, – снижение; значимые различия на уровне парных сравнений)
| Области коры | Диапазоны | Задания | Изменения | |
|---|---|---|---|---|
| изолир. | двойные | |||
| Лобно-височн. лев. | δ | С1 | С1М1 | – – |
| С2 | С2М1 | – | ||
| С3 | С3М1 | + | ||
| θ | С1 | С1М1 | 0 | |
| С2 | С2М1 | 0 | ||
| С3 | С3М1 | 0 | ||
| α1 | С1 | С1М1 | + | |
| С2 | С2М1 | 0 | ||
| С3 | С3М1 | 0 | ||
| α2 | С1 | С1М1 | + | |
| С2 | С2М1 | 0 | ||
| С3 | С3М1 | 0 | ||
| β и γ | С1 | С1М1 | 0 | |
| С2 | С2М1 | 0 | ||
| С3 | С3М1 | 0 | ||
| Лобно-височн. прав. | δ | С1 | С1М1 | 0 |
| С2 | С2М1 | 0 | ||
| С3 | С3М1 | + | ||
| θ | С1 | С1М1 | + | |
| С2 | С2М1 | + | ||
| С3 | С3М1 | + | ||
| α1 | С1 | С1М1 | – | |
| С2 | С2М1 | + | ||
| С3 | С3М1 | + + | ||
| α2 | С1 | С1М1 | 0 | |
| С2 | С2М1 | 0 | ||
| С3 | С3М1 | + | ||
| β и γ | С1 | С1М1 | 0 | |
| С2 | С2М1 | 0 | ||
| С3 | С3М1 | + | ||
| Центр.-темен.-затыл. лев. | δ | С1 | С1М1 | – |
| С2 | С2М1 | 0 | ||
| С3 | С3М1 | + | ||
| θ | С1 | С1М1 | + | |
| С2 | С2М1 | + | ||
| С3 | С3М1 | + | ||
| α1 | С1 | С1М1 | + | |
| С2 | С2М1 | 0 | ||
| С3 | С3М1 | – | ||
| α2 | С1 | С1М1 | + | |
| С2 | С2М1 | – | ||
| С3 | С3М1 | 0 | ||
| β и γ | С1 | С1М1 | 0 | |
| С2 | С2М1 | 0 | ||
| С3 | С3М1 | 0 | ||
| Центр.-темен.-затыл. прав. | δ | С1 | С1М1 | 0 |
| С2 | С2М1 | 0 | ||
| С3 | С3М1 | – | ||
| θ | С1 | С1М1 | + | |
| С2 | С2М1 | + | ||
| С3 | С3М1 | + | ||
| α1 | С1 | С1М1 | + | |
| С2 | С2М1 | + | ||
| С3 | С3М1 | 0 | ||
| α2 | С1 | С1М1 | + | |
| С2 | С2М1 | + | ||
| С3 | С3М1 | 0 | ||
| β и γ | С1 | С1М1 | 0 | |
| С2 | С2М1 | 0 | ||
| С3 | С3М1 | 0 | ||
ОБСУЖДЕНИЕ РЕЗУЛЬТАТОВ
Результаты данного исследования подтвердили гипотезу о том, что выполнение двойных задач обеспечивается регионарным и частотным распределением активации мозга, отличающимся для каждого из заданий – когнитивного и моторного. Результаты исследования подтвердили гипотезу об особенностях функциональной организации мозга при решении двойных задач.
Результаты в целом согласуются с ранее полученными данными, выполненными на другой группе испытуемых при использовании других когнитивных задач [22, 23]. Было выявлено “включение” медленных ритмов ЭЭГ в лобно-височных отделах левого полушария при выполнении счетно-логических когнитивных задач. В отличие от этого выполнение моторной задачи сопровождалось увеличением числа функциональных связей для более частых ритмов ЭЭГ, с большей активацией центрально-теменно-затылочных отделов правого полушария [22–26].
Результаты исследования в деталях отличаются от предыдущих исследований, в частности, выявлено преимущественное доминирование реактивных перестроек ∆-диапазона по сравнению с θ-, что может быть обусловлено использованием других когнитивных проб. Увеличение числа пар отведений, демонстрирующих усиление функциональных связей, при выполнении более сложных когнитивных задач, особенно при изолированном их выполнении, может отражать увеличение отделов коры больших полушарий, необходимых для обеспечения более сложных процессов.
Важно отметить, что в большом числе исследований, не связанных с двойными задачами, осуществление когнитивной деятельности человека связывают с высокочастотными ритмами ЭЭГ – β и γ [40]. Однако имеются работы, в которых выявлена фазовая синхронизация θ-колебаний при выполнении когнитивной деятельности как изолированно, так и в составе двойных задач [33]. Важная роль медленных ритмов, в частности ∆-диапазона, выявлена при осуществлении различных видов когнитивной деятельности и в других исследованиях [41].
Сопоставление характера реактивных перестроек при выполнении различных когнитивных заданий в данном исследовании показало, что при изолированном выполнении счетно-логических задач наблюдалось увеличение когерентности медленных ритмов ЭЭГ в лобно-височных отделах левого полушария. В отличие от этого при выполнении пространственно-образных задач “включение” медленных ритмов ЭЭГ наблюдалось также и в правом полушарии со смещением фокуса активации в теменно-затылочные области, что может быть связанно с пространственно-образным синтезом.
Результаты исследования помогли узнать – как мозг человека одновременно решает две “правополушарные” задачи? Особенностью реактивных перестроек ЭЭГ при выполнении пространственно-образной задачи являлось “включение” медленных ритмов ЭЭГ в правом полушарии. В отличие от этого правополушарные реактивные перестройки мозга при выполнении моторной задачи (произвольный позный контроль) обеспечивались более частыми (α–β–γ) ритмами ЭЭГ. Вероятно, что именно такое частотное распределение активации мозга в правом полушарии обеспечивает возможность одновременного выполнения двух “правополушарных” задач – моторной и пространственно-образной. В целом полученные в данном исследовании данные совпадают с результатами наших предыдущих исследований, в которых в качестве моторного компонента двойной задачи использовался произвольный позный контроль [22, 23]. Более того, полученные нами результаты подтверждены недавними ЭЭГ-исследованиями [29], которые также показали, что выполнение задания с произвольным поддержанием определенной позы на стабилографической платформе как изолированное, так и в составе двойной задачи, сопровождается увеличением связей в высокочастотном диапазоне α- и β-диапазонах между фронто-темпоральными и парието-окципитальными отделами правого полушария Реактивные перестройки высокочастотного α- и β-диапазонов были выявлены и при выполнении других моторных нагрузок, что может быть базисным механизмом организации моторных функций человека [42, 43].
Преимущественно правополушарная активация мозга была выявлена при поддержании и изменении вертикальной позы (положение лежа–сидя–стоя) у здоровых людей [24, 25]. Интересно заметить, что у космонавтов после длительных космических полетов, сопровождавшихся нарушениями поддержания вертикальной позы, по данными фМРТ (в состоянии покоя – resting state) наблюдалось максимальное снижение эффективности связей именно в теменно-центральных отделах правого полушария [44]. Правое полушарие мозга, более старое в филогенетическом плане, кроме позного контроля, обеспечивает и другие биологически важные базисные функции организма – поддержание гомеостаза, обеспечение функции сон–бодрствование и др. [13]. Однако эти факты вовсе не означают исключительное обеспечение перечисленных функций правым полушарием. Фронто-париетальные отделы правого полушария играют важную роль в выполнении и когнитивных функций, таких как направленное внимание и рабочая память и др., как у здоровых людей, так и у пациентов с церебральной патологией [45].
При сравнении регионарных и частотных реактивных перестроек мозга при выполнении двойных и изолированных задач было выявлено преимущественное снижение числа функциональных связей при выполнении двойных задач, что согласуется с результатами наших предыдущих исследований и работ других авторов [23, 29]. Эти ЭЭГ-данные помогают объяснить результаты поведенческих исследований, свидетельствующих о более низком качестве выполнение двойных задач, по сравнению с изолированными, преимущественно когнитивного компонента [9, 10]. Эти факты авторы объясняют дефицитом когнитивного ресурса в результате обеспечения выполнения обеих задач.
Заслуживают особого внимания увеличение корково-корковых связей α1-диапазона в лобно-центрально-теменных отделах как правого, так и левого полушария, что отчетливо проявляется при выполнении двойных задач, но не наблюдается при изолированном выполнении заданий. Вероятно эти ЭЭГ-данные подтверждают мнение о том, двойные задачи являются более сложным процессом, чем простое сложение процессов активации соответствующих областей мозга, выявленных при выполнении для каждого из заданий и требующим участия дополнительных (или других) структур мозга [6]. Ранее изменения в α1-диапазоне в лобно-центрально-теменных областях были названы нами “областями перекрытия интересов” [23]. У “успешных” испытуемых, способных более успешно выполнять двойные задачи по сравнению с изолированными, не наблюдалось увеличения количества связей или даже наблюдалось их снижение, особенно в лобных отделах [23]. Такие испытуемые характеризовались более высокими когнитивными ресурсами, что сочеталось с высоким моторным мастерством. В отличие от этого у испытуемых, выполнявших двойные задачи хуже, чем изолированные, нарастание числа связей α1-диапазона и в дополнительных отделах мозга может отражать использование дополнительных ресурсов мозга на выполнение двойных задач.
ЗАКЛЮЧЕНИЕ
У здоровых людей при изолированном и одновременном выполнении когнитивных и моторных задач выявлены регионарные и частотные реактивные перестройки ЭЭГ, характерные для каждого из заданий. Выполнение моторного компонента сопровождалось увеличением когерентности частых ритмов ЭЭГ (α-, β- и γ-) в центрально-теменно-затылочных отделах правого полушария. Специфика функциональной организации мозга при выполнении счетно-логических задач состояла в нарастании когерентности медленных составляющих ЭЭГ в лобно-височных отделах левого полушария, в то время как при выполнении пространственно-образных задач выявлено дополнительное увеличение когерентности медленных ритмов ЭЭГ и в правом полушарии. Можно полагать, что такое частотное распределение активации мозга человека в правом полушарии для двух правополушарных задач (зрительно-пространственных и моторных) может быть основным нейрофизиологическим механизмом, обеспечивающим их одновременное выполнение. При выполнении двойных задач наблюдалось снижение суммарного числа связей, характерных для изолированного выполнения моторного и особенно когнитивного компонента, что сочеталось с увеличением числа функциональных связей в α1-диапазоне в лобно-центрально-теменных отделах обоих полушарий, что является отражением использования дополнительных ресурсов мозга.
Работа поддержана грантом РФФИ-ОГН № 17-06-01012.
Список литературы
Camiocioli R., Howieson D., Lehman S. Talking while walking: the effect of a dual task in aging and Alzheimer’s disease // Neurology. 1997. V. 48. P. 955.
Resch J.E., May B., Tomporowski D., Ferrara M.S. Balance performance with cognitive tasks: a continuation of the dual-task testing paradigm // J. of Athletic Training. 2011. V. 46. P. 170.
Laufer Y., Ashkenazi T., Josman N. The effects of a concurrent task cognitive task on the postural control of young children with and without developmental coordination disorder // Gait and Posture. 2008. V. 27. № 2. P. 347.
Al-Yahya E., Dawes H., Collett J. et al. Gait adaptations to simultaneous cognitive and mechanical constraints // Exp. Brain Res. 2009. V. 199. № 1. P. 39.
Hartman P., Cockburn J. Dividing attention between cognitive and motor tasks in neurological rehabilitation // Neuropsychological Rehabilitation 1998. V. 8. № 2. P. 155.
Naschimbeni A., Gaffuri A., Penno A., Tavoni M. Dual task interference during gait in patients with unilateral vestibular disorders // J. of Neuroenginer and Rehabil. 2010. V. 7. № 1. P. 7.
Hamilton F., Rochester L., Paul L. et al. Walking and talking: an investigation of cognitive-motor dual tasking in multiple sclerosis // Mult. Scler. 2009. V. 15. P. 1215.
Salo E., Rinne T., Salonen O., Alho K. Brain activation during bimodal dual tasks depend on the nature and combination of component tasks // Frontiers in Human Neuroscience. 2015. V. 9. № 2. P. 1.
Hiraga C.Y., Garry M.I., Carson R.G., Summers J.J. Dual-task interference: attentional and neuropsychological influences // Behav. Brain Res. 2009. V. 205. № 1. P. 10.
Naschimbeni A., Gaffuri A., Penno A., Tavoni M. Dual task interference during gait in patients with unilateral vestibular disorders // J. of Neuroengeginerner and Rehab. 2010. V. 7. № 1. P. 7.
Haggard P., Cockburn J., Cock J. et al. Interference between gait and cognitive tasks in a rehabilitating neurological population // J. Neurol. Neurosurg Psychiatry. 2000. V. 69. № 4. P. 479.
Stoffregen T.A., Hove P., Bardy B.G. et al. Postural sta-bilization of perceptual but not cognitive performance // J. Mot. Behav. 2007. V. 39. № 2. P. 126.
Доброхотова Т.А., Брагина Н.Н. Функциональная асимметрия и психопатология очаговых поражений мозга. М.: Медицина, 1977. 53 с.
Schaefer S., Lindenberger U. Thinking while walking: experienced high-heel walkers flexibility adjust their gait // Frontiers in Psychology. 2013. V. 4. № 4. P. 1.
Жарикова А.В., Жаворонкова Л.А., Купцова С.В. и др. Психологические и стабилографические характеристики здоровых людей с разным качеством выполнения двойных задач // Физиология человека. 2013. Т. 39. № 4. С. 33.
Doumas M., Rapp M., Krampe R. Working memory and postural control: adult age differences in potential for improvement, task priority, and dual tasking // J. of Gerontology. 2009. V. 64. № 2. P. 193.
Schafer S., Lang K. Using dual tasks to test immediated transfer to training between naturalistic movements: a proof-of-principle study // J. Mot. Behave. 2012. V. 44. № 3. P. 313.
Schaefer S., Krampe R., Lindenberger U., Baltes P. Age differences between children and young adults in the dynamics of dual-task prioritization: body (balance) versus mind (memory) // Dev. Psychol. 2008. V. 44. № 3. P. 747.
Stelzel C., Brandt S., Schubert T. Neural mechanisms of concurrent stimulus in dual tasks // Neuroimage. 2009. V. 48. № 2. P. 237.
Schumacher E., Schwarb H. Parallel response selection disrupts sequence learning under dual-task conditions // J. of Experimental Psychology: General. 2009. V. 138. № 2. P. 270.
Low K.A., Leaver E.E., Kramer A.F. et al. Share or complete? Load-dependent recruitment of prefrontal cortex during dual-task performance // Psychophysiology. 2009. V. 46. № 5. P. 1069.
Жаворонкова Л.А., Жарикова А.В., Кушнир Е.М. и др. Особенности реактивных перестроек ЭЭГ при выполнении двойных задач здоровыми испытуемыми (произвольный позный контроль и счет) // Физиология человека. 2011. Т. 37. № 6. С. 54.
Жаворонкова Л.А., Кушнир Е.М., Жарикова А.В. и др. Электроэнцефалографические характеристики здоровых людей с разной успешностью выполнения двойных задач (позный контроль и счет) // Журн. высшей нервн. деят. 2015. Т. 65. № 5. С. 1.
Жаворонкова Л.А., Шевцова Т.П., Максакова О.А. Как мозг человека одновременно решает две задачи? Изд-во: LAP LAMBERT Academic Publishing. Deutschland, Saarbruchen, 2017. 60 с.
Zhavoronkova L., Shevtsova T., Pozdneev A. et al. Brain mechanisms impairment of dual-task processing in traumatic brain injury patients // Functional Neurology, Rehabilitation, and Ergonomics. 2017. V. 7. № 3. P. 253.
Zhavoronkova L., Zharikova A., Maksakova O. Why voluntary postural training improves recovery of mental and motor functions in patients with traumatic brain injury? // J. of Behavioral and Brain Sciences. 2013. V. 3. № 6. P. 463.
Miller E.K., Cohen J.D. An integrative theory of prefrontal cortex function // Ann. Reviewer Neuroscience. 2001. V. 24. № 2. P. 167.
Wang Y-K., Jung T-P., Lin C-T. Theta and alpha oscillations in attentional interaction during distracted driving // Frontiers in Behavioral Neuroscience. 2018. V. 12. № 3. P. 1.
Huang C.-Y. Chang G.-C., Tsai Y.-Y., Hwang I.-S. An increase in postural load facilitates an anterior shift processing resources to frontal executive function in a postural-suprapostural task // Frontiers in Human Neuroscience. 2016. V. 10. P. 420.
Pizzamiglio S., Abdalla H., Naeem U., Tuner D.L. Neural predictor of gait stability when walking freely in real-world // J. of Neuroengineering and Rehabilitation. 2018. V. 15. № 11. P. 1.
Raduntz T. Dual frequency head maps: a new method for indexing mental workload continuously during execution of cognitive tasks // Frontiers in Psychology. 2017. V. 8. № 4. P. 1019.
Blundon E., Rumak S.P., Ward L.M. Sequential search asymmetry: behavioral and psychological evidence from a dual oddball task // Plos One. 2017. V. 12. № 3. P. 1.
Kawasaki M., Kinajo K., Yamaguchi Y. Fronto-parietal and fronto-temporal theta- phase synchronization for visual and auditory-verbal working memory // Frontiers in Psychology. 2014. V. 5. № 4. P. 1.
Русинов В.С., Гриндель О.М., Болдырева Г.Н., Вакар Е.М. Биопотенциалы мозга человека (математический анализ). М.: Медицина, 1987. 254 с.
Гриндель О.М. Электроэнцефалограмма человека при черепно-мозговой травме. М.: Наука, 1988. 200 с.
Шарова Е.В., Челяпина М.В., Коробкова Е.В. и др. ЭЭГ-корреляты восстановления сознания после тяжелой черепно-мозговой травмы // Журн. Вопросы нейрохирургии им. Н.Н. Бурденко. 2014. Т. 78. № 1. С. 14.
Фарбер Д.А., Мачинская Р.И. Влияние функционального состояния коры и подкорковых структур головного мозга на системную организацию когнитивную деятельность у детей младшего школьного возраста и подростков. М.: Вентаграф, 2005. 215 с.
Rappelsberger P., Petsche H. Probability mapping: power and coherence analysis of cognitive processes // Brain Topography. 1988. V. 1. № 1. P. 46.
Thatcher R.W. Cyclic cortical reorganization, origins of human cognitive development / Human behaviour and the development brain. Eds. Dawson G., Fisher K. N.Y.: Guilfords Press, 1994. P. 232.
Danilova N.N. Frequency-selective gamma generators in processing of auditory stimuli // Psychology in Russia: State of the Art / Eds. Zinchenko Y., Petrenko V. M.: Department of Psychology MSU & IG-SOCIN, 2008. 388 p.
Harmony T. The functional significance of delta oscillation in cognitive processing // Front. Integrative Neurosciences. 2013. V. 7. № 1. P. 1.
Болдырева Л.А., Жаворонкова Л.А., Шарова Е.В. и др. фМРТ-ЭЭГ исследование реакций мозга здорового человека на функциональные нагрузки // Физиология человека. 2009. Т. 35. № 3. С. 20.
Болдырева Г.Н., Шарова Е.В., Жаворонкова Л.А. и др. Сопоставление фМРТ-реакций мозга здоровых людей при активных, пассивных и воображаемых движениях рукой // Медицинская визуализация. 2015. Т. 3. № 5. С. 1.
Demertzi A., Van Ombergen A., Tomilovskaya E. et al. Cortical Reorganization an austranaut`s brain after duration spaceflight // Brain Struct. Funct. 2016. V. 221. № 5. P. 2873.
Robertson I.H. Right hemisphere role in cognitive reserve // Neuirobiol. Aging. 2014. V. 35. № 6. P. 1375.
Дополнительные материалы отсутствуют.
Инструменты
Физиология человека