

Физиология человека, 2021, T. 47, № 2, стр. 83-91
Анализ нелинейных свойств ЭМГ для обнаружения изменений синхронизации двигательных единиц при эксцентрическом и концентрическом режимах мышечного сокращения
К. В. Сергеева 1, *, Р. В. Тамбовцева 1
1 ФГБОУ ВО Российский государственный университет
физической культуры, спорта, молодежи и туризма
Москва, Россия
* E-mail: sergeeva_xenia@mail.ru
Поступила в редакцию 10.01.2020
После доработки 31.01.2020
Принята к публикации 01.06.2020
Аннотация
Впервые с помощью рекуррентного количественного анализа данных поверхностной ЭМГ проведена косвенная оценка синхронизации импульсной активности двигательных единиц (ДЕ) четырехглавой мышцы бедра в эксцентрическом и концентрическом режимах мышечного сокращения при выполнении упражнения разгибание голени с интенсивностью 25, 50, 80 и 100% от индивидуального эксцентрического и концентрического произвольного максимума. В частности, было установлено, что % детерминизма (% DET), являющегося чувствительным индексом, позволяющим обнаруживать синхронизацию ДЕ, был незначительно выше во время эксцентрических сокращений. Также выявлено, что с увеличением величины отягощения степень синхронизации снижается для обоих режимов мышечных сокращений (p < 0.05). % DET, вычисляемый в ходе количественного анализа рекуррентной диаграммы, показал чувствительность к изменению режима и интенсивности сокращения и его можно использовать для оценки синхронности работы ДЕ.
Большая часть классических исследований физиологии мышечной деятельности, сформировавших основу базового понимания работы мышц, основаны на изучении двух типов мышечных сокращений: изометрическом (постоянная длина мышцы) и концентрическом (укорочение мышцы при постоянной нагрузке). К сожалению, о механике и энергетике мышц во время активного удлинения известно гораздо меньше. Этот режим представляет большой интерес и заслуживает подробного рассмотрения.
Если момент прилагаемого мышечного усилия меньше момента противодействующей силы, то мышца удлиняется под воздействием отягощения в уступающем или эксцентрическом режиме, который требует небольшого количества метаболической энергии и характеризуется высоким производством силы. Человек в состоянии опустить вес на 20–60% больше концентрического максимума [1]. Способность мышц проявлять значительную механическую силу обусловлена в значительной мере пассивными силами структурного белка титина [2]. По этой причине уступающая фаза движения требует активации меньшего совокупного количества волокон или/и работы двигательных единиц (ДЕ) с меньшей частотой разрядов потенциалов действия (ПД) мотонейрона, что в итоге дает меньшую электромиографическую (ЭМГ) активность. Даже в условиях максимального эксцентрического сокращения, когда активность ЭМГ была сопоставима с концентрическим режимом [3, 4], импульсация мотонейронов оставалась ниже. Авторы полагают, что помимо структурных белков бо́льшая выработка силы в эксцентрическом режиме может обеспечиваться за счет избирательного рекрутирования быстрых мышечных волокон [5] и увеличения синхронизации двигательных единиц, приводящей к суммации сокращений отдельных ДЕ [4, 6]. Таким образом, важно подтвердить вероятность бо́льшей синхронной активности ДЕ в эксцентрическом режиме, как механизма, контролирующего силу мышечных сокращений. Только в одном исследовании J.G. Semmler et al. [7] изучалась степень синхронизации между парами ДЕ во время эксцентрических сокращений. Авторы обнаружили, что частота синхронных разрядов была на 50% больше во время эксцентрических сокращений по сравнению с концентрическими сокращениями первой дорсальной межкостной мышцы кисти, когда испытуемые поднимали и опускали нагрузку, не превышающую 5% от максимальной произвольной силы (МПС). Однако используемая техника кросс-корреляционного анализа активности пар ДЕ во временнóй области имеет ряд недостатков: 1) она инвазивна и вызывает дискомфорт, 2) записи пар произвольно активированных ДЕ могут быть достоверно получены только при слабых уровнях тонического сокращения мышц, 3) этот метод не может дать никакой информации о нелинейных особенностях сигнала [8], являющихся отличительной чертой сложных динамических систем. Между тем, за последние десятилетия набор традиционных (линейных) методов исследования временны́х рядов был существенно расширен нелинейными методами, полученными из теории нелинейной динамики и хаоса, которые нашли свое применение во многих сферах науки, в частности, оказались более надежным по сравнению с традиционными линейными методами в оценке степени синхронизации ДЕ для ранней диагностики болезни Паркинсона, дифференциальной диагностики похожих нейродегенеративных заболеваний, разных форм тремора [9], в том числе для оценки степени утомления мышц [10].
Наиболее перспективным в данном направлении исследования является нелинейный метод рекуррентного количественного анализа (Recurrence Quantification Analysis, RQA), который не требует априорных ограничений на размер данных, стационарность временны́х рядов и статистическое распределение [8, 11]. Физиологические сигналы характеризуются фундаментальным свойством – рекуррентностью (повторяемостью), выражающейся в том, что при введении определенного уровня энтропии в ограниченную и упорядоченную динамическую систему, она через некоторое время достигает состояния, некоторым образом близкого к предыдущему, и проходит при этом подобные этапы эволюции. RQA определяет количество и продолжительность повторений динамической системы, представленной траекторией фазового пространства. Рекуррентные диаграммы, предложенные в 1987 г. Экманом (Eckmann), позволяют отобразить фазовую траекторию любой размерности на двумерную двоичную квадратную матрицу, размер которой определяется длиной временнóго ряда, через ее рекуррентности [12]. Помимо мощных визуальных возможностей, существует метод количественного анализа структур, формируемых на изображении рекуррентной диаграммы, предложенные J.P. Zbilut и C.L. Webber, Jr. [13]. Так, % детерминизма (% DET) является чувствительным индексом, позволяющим обнаруживать синхронизацию импульсной активности ДЕ, поскольку отражает количество подчиняющихся правилам структур в стохастическом сигнале. Эта переменная доказала свою надежность, как неинвазивного и эффективного средства для извлечения информации о синхронном поведении ДЕ наряду с кросс-корреляционным анализом [14].
Цель исследования – рекуррентный анализ данных поверхностной ЭМГ для косвенной оценки возможных различий в синхронной активности ДЕ во время эксцентрических и концентрических мышечных сокращений с различной интенсивностью.
МЕТОДИКА
В эксперименте принимали участие 9 представителей скоростно-силовых видов спорта. Экспериментальное исследование проводили в лаборатории “работоспособности НИИ спорта и спортивной медицины” на базе “Российского государственного университета физической культуры, спорта, молодежи и туризма (ГЦОЛИФК)” (г. Москва).
Биоэлектрическую активность четырехглавой мышцы бедра получали при 25, 50, 80 и 100% от максимального произвольного эксцентрического и концентрического максимума с использованием 16-канального электромиографа ME6000 BiomonitorSystem (Mega Electronics Ltd., Финляндия) при выполнении упражнения разгибание голени в тренажере. Эксцентрический максимум соответствовал 140% от концентрического максимума. Данный процент был выбран согласно литературным экспериментальным данным в разнице максимальной эксцентрической и концентрической динамической силе [15]. Темп работы задавался метрономом. Длительность каждого сокращения составляла 2 с. Тестовое движение выполнялось по команде с фиксацией исходной и финальной позы, что отражалось на паттерне ЭМГ, по которому и определяли временны́е границы биоэлектрической активности исследуемых мышц. Сигнал ЭМГ записывали с помощью электродов Ag/AgCl (диаметр 50 мм). Частота дискретизации сигнала 1000 Гц. Полоса пропускания ЭМГ составила 3–500 Гц. Электроды располагали на средней части мышечного брюшка параллельно продольной оси мышечных волокон с межэлектродным расстоянием 20 мм. Кожу испытуемых предварительно обрабатывали спиртом. Для анализа электрической активности четырехглавой мышцы плеча находили среднеквадратическое значение (RMS). Расчет нелинейных динамических характеристик исследуемых временны́х рядов производили с помощью программного продукта 4.9.1.1 Visual Recurrence Analysis. Записанные временны́е ряды были разбиты на более короткие окна (N = 4000) или эпохи, каждая длиной 0.5 с. Соседние окна были смещены на 2000 точек (50% перекрытия). Для каждого скользящего окна вычисляли % DET, затем значения усредняли.
Более подробная информация об этом аналитическом инструменте описана в работе [12]. Процедура была основана на внедрении миоэлектрических данных в N-мерное евклидово пространство. Дискретные данные ЭМГ s(k) (k = = 0, …, n – 1), n = 4000 задерживались во времени на число λ (“временнáя задержка”) между последующими временны́ми точками. Значение временнóй задержки вычисляли с помощью функции средней взаимной информации, чтобы свести к минимуму взаимодействие между точками измеряемого временнóго ряда. В настоящем исследовании значения λ находились в диапазоне от 1 до 2.
Данные встраивались в размерное пространство (M). Оптимальные размеры вложения получали с помощью алгоритма ложного ближайшего соседа. Следующий набор M-мерных векторов (n) извлекался из исходного миоэлектрического временнóго ряда:
(1)
$\begin{array}{*{20}{c}} {v\left( 1 \right) = }&{~\{ s\left( 1 \right)}&{s\left( {\lambda + 1} \right)~\,\,~\,\, \ldots }&{s\left[ {\left( {M - 1} \right)\lambda } \right]\} } \\ {v\left( 2 \right) = }&{~\{ s\left( 2 \right)}&{s\left( {\lambda + 2} \right)\,\,\,\,~ \ldots }&{~s\left[ {\left( {M - 1} \right)\lambda + 1} \right]\} } \\ \ldots & \ldots &{ \ldots ~}& \ldots \\ {v\left( N \right) = }&{~\{ s\left( N \right)}&{s\left( {N + \lambda } \right)~\,\, \ldots }&{~s\left[ {N + \left( {M - 1} \right)\lambda } \right]\} ,} \end{array}$Состояния рассматриваемой динамической системы представляются векторами v(n), определенными в уравнении (1). Чтобы определить рекуррентность между состояниями, их близость должна быть количественно оценена. Для этого вычисляли евклидово расстояние (d) между всеми возможными векторными парами:
(2)
$d\left( {i,j} \right) = {{\{ \left\langle {{{{[v\left( i \right) - v\left( j \right)]}}^{2}}} \right\rangle \} }^{{\frac{1}{2}}}}~.$(3)
$DM = \left\| {\begin{array}{*{20}{c}} {d\left( {1,1} \right)}&{d\left( {1,2} \right)}&{d\left( {1,N} \right)} \\ {d\left( {2,1} \right)}&{d\left( {2,2} \right)}&{d\left( {2,N} \right)} \\ \ldots & \ldots & \ldots \\ {d\left( {N,1} \right)}&{d\left( {N,2} \right)}&{d\left( {N,N} \right)} \end{array}} \right\|~~.$Внешний вид полученной и визуализированной матрицы дает представление о динамике встроенных сигналов в виде точек, изначально представленных в виде временнóго ряда. Одна точка соответствует повторению состояния при некотором времени i в некоторое другое время j, а обе координатные оси являются осями времени. Две точки считаются рекуррентными, если находятся достаточно близко друг к другу, т.е. расстояние d (i, j) меньше порогового значения (называемого радиус, R). В случае, когда сегмент траектории в фазовом пространстве пролегает параллельно другому сегменту, т.е. траектория повторяет саму себя, возвращаясь в одну и ту же область фазового пространства в разное время, появляются диагональные линии. Системы, обладающие детерминированной (подчиняющейся правилам) динамикой, характеризуются диагональными линиями, указывающими на повторяющиеся рецидивы внутри состояния. % DET измеряет долю повторяющихся точек (Re), образующих диагональные линии (L), которые состоят из 2 или более точек без пробелов:
Статистическую обработку результатов исследования выполняли при помощи пакета программ Statistica 13.3. Для проверки значимости различий результатов проводили 2-х факторный дисперсионный анализ с повторениями. Факторы: режим (эксцентрический/концентрический) и % МПС (25, 50, 80, 100%). Для выявления значимых различий в результатах между группами были проведены апостериорные тесты с поправкой Бонферрони по факторам “режим” и “% МПС”. Корреляционный анализ между параметрами производили с помощью критерия Пирсона. Для всех тестов был установлен уровень значимости р < 0.05.
РЕЗУЛЬТАТЫ ИССЛЕДОВАНИЯ И ИХ ОБСУЖДЕНИЕ
Для % DET был обнаружен значимый эффект “% МПС” (F = 5, 4; p < 0.05), “режима” (F = 8, 9; p < 0.05), но не их взаимодействия. Как видно на рис. 1 и 2, эксцентрические сокращения сопровождались несколько более высокими значениями % DET, что косвенно может отражать более синхронный паттерн работы ДЕ, но апостериорный тест выявил статистически значимые различия между группами только при интенсивности 25% МПС (р < 0.05).
Рис. 1.
Динамика процента детерминизма сигнала (% DET, кривая а) и среднеквадратического значения (RMS, кривая б) электромиограммы (ЭМГ) четырехглавой мышцы бедра при увеличении интенсивности от 25 до 100% МПС в эксцентрическом режиме мышечного сокращения. Значения представлены как среднее ± σ.
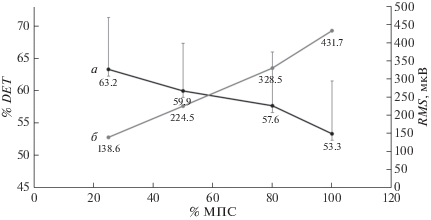
Рис. 2.
Динамика процента детерминизма сигнала (% DET, кривая а) и среднеквадратического значения (RMS, кривая б) электромиограммы (ЭМГ) четырехглавой мышцы бедра при увеличении интенсивности от 25 до 100% МПС в концентрическом режиме мышечного сокращения. Значения представлены как среднее ± σ.
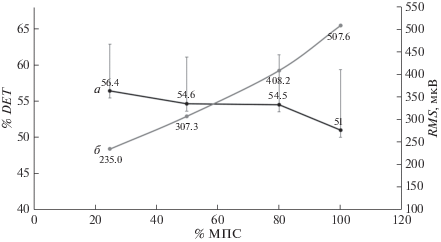
Кроме того, с увеличением интенсивности сокращения и, как следствие, суммарной амплитуды сокращения, представленной среднеквадратическим значением (RMS), синхронизация снижалась в обоих режимах сокращения (рис. 1, 2). Наблюдалась значительная отрицательная корреляция (данные не показаны) между % МПС и % DET (r = 0.9 и 0.8 для эксцентрического и концентрического режима соответственно).
На рис. 3, А показан пример рекуррентной диаграммы при эксцентрическом сокращении с интенсивностью 100% МПС, где можно наблюдать короткие диагональные структуры, характерные для хаотических процессов, количественно выраженные низким значением % DET. На рис. 3, Б показан пример рекуррентной диаграммы при эксцентрическом сокращении, но уже с интенсивностью 25% МПС. Очевидна рекуррентная динамика, которая имеет вид более длинных диагональных линий, разделенных фиксированным расстоянием. Диагональные отрезки линий из точек, идущих близко друг к другу последовательно во времени указывают на стабильную периодичность для временнóго интервала, соответствующего длине диагонали и, следовательно, на более синхронную работу ДЕ.
Рис. 3.
Рекуррентные диаграммы ЭМГ-сигнала при эксцентрических сокращениях. А – интенсивность – 100% от МПС; % DET = 49; RMS = 786 мкВ. Б – интенсивность – 25% от МПС; % DET = 80; RMS = 74 мкВ. Оси координат имеют размерность “время”, равную длине временнóго ряда. Участки, выделенные черными квадратами, представлены в увеличенном масштабе внизу.

Наши данные согласуются с результатами современных работ [16, 17], посвященных изучению произвольных изометрических сокращений с применением метода декомпозиции осцилограммы, где было установлено, что степень синхронизации уменьшается по мере увеличения интенсивности, что противоречит классическим представлениям о роли синхронизации в контроле силы мышечного сокращения. Авторы данных работ заключили, что степень синхронизации, являясь не более чем эпифеноменом паттерна импульсации ДЕ, зависит от двух параметров: одинаковой частоты импульсации ДЕ (одинаковый порог рекрутирования) и крутизны наклона кривой частоты импульсации. Другими словами, совпадение во времени отдельных импульсов увеличивается, если рекрутируемые ДЕ имеют одинаковую частоту импульсации и высокую скорость нарастания частоты импульсации. Так, при мышечных сокращениях с постепенно нарастающим усилием первые вовлекаемые низкопороговые ДЕ, наиболее чувствительные к произвольному возбуждению, имеют одинаковый порог рекрутирования и одинаковую, инкрементно возрастающую частоту импульсации, что способствуют увеличению вероятности совпадения во времени отдельных сократительных циклов разных ДЕ в начале движения, так как соблюдаются оба фактора. По мере увеличения силы, создаваемой мышцей, низкопороговые ДЕ увеличивают частоту разрядов, крутой начальный наклон плавно переходит в более плоскую, линейную область, чувствительность к произвольному возбуждению и вместе с ним синхронизация снижаются [18]. При этом в работу включаются высокопороговые быстрые ДЕ с одинаковой низкой частотой импульсации, но более пологим наклоном кривой импульсации ДЕ, что приводит к относительно меньшему количеству синхронных моментов разрядов ПД. И так далее, с увеличением амплитуды сокращения все более крупные по размеру ДЕ и меньшими углами наклона частоты импульсации вовлекаются в работу, а уже работающие ДЕ увеличивают частоту разряда и также снижают наклон [17]. Таким образом, из вышесказанного следует вывод, что, чем выше % МПС, тем ниже синхронность ДЕ.
Необходимо отметить один принципиальный момент, указанный выше, который остается предметом споров в настоящее время, а именно, различия в скорости импульсации высокопороговых и низкопороговых ДЕ. Более пяти десятилетий назад J.C. Eccles et al. [19] опубликовали данные экспериментов, проведенных на кошках, где сообщалось о том, что при электростимуляции фазные (высокопороговые) мотонейроны демонстрируют более короткую постгиперполяризацию, чем тонические (низкопороговые) мотонейроны, постулируя, что длительность постгиперполяризации является доминирующим фактором в определении частоты импульсации мотонейронов. Согласно этим представлениям, низкопороговые ДЕ, в отличие от высокопороговых, характеризуются низкой частотой импульсации, и с повышением силы сокращения мышцы частота разрядов медленных мотонейронов возрастает незначительно. Эти данные впоследствии были подтверждены рядом исследований [20–22]. Принципиальный момент возник, когда перешли к изучению особенностей работы ДЕ во время произвольных изометрических сокращений у людей. Многочисленные исследования, начиная с 1940 г., когда данные игольчатой ЭМГ анализировались путем визуальной оценки [23–25], и заканчивая современными исследованиями, в которых использовались различные сложные методы декомпозиции ЭМГ-сигналов [26–38], предоставляют данные об иерархической обратной зависимости между порогом рекрутирования и частотой разрядов ПД мотонейронов. Оказалось, что работы, где была обнаружена положительная связь между порогом рекрутирования и частотой импульсации [20–22], страдают ошибочной методологией измерений. В частности, в данных экспериментах возбуждение мотонейрона поддерживалось при минимальной фиксированной скорости разрядов ПД путем выполнения градуированного увеличения или уменьшения силы, не превышая при этом 5% МПС, что ни в коей мере нельзя экстраполировать на модель работы ДЕ при произвольных сокращениях. В случае с естественным переменным возбуждением, после первого срабатывания мотонейрона, поток нервных импульсов к нему возрастает до последующего срабатывания. Такое увеличение возбуждения приводит к тому, что мотонейрон срабатывает быстрее, уменьшая межимпульсный интервал и приводя к большей частоте разрядов ПД. Ранее рекрутированные (меньшего диаметра, с низким порогом возбуждения) мотонейроны имеют меньшую константу времени [26, 27] и, следовательно, экспоненциальная функция, описывающая частоту импульсации, увеличивается быстрее в зависимости от времени, что делает ранее рекрутированные мотонейроны более чувствительными к возрастающему возбуждению. Следовательно, ранее рекрутированные низкопороговые ДЕ имеют бо́льшую частоту импульсациии и скорость ее нарастания, чем позже рекрутированные высокопороговые ДЕ при любом заданном уровне силы [26], что отражается на степени синхронизации.
В схеме регуляции силы, описанной выше, сокращения высокопороговых быстрых волокон не сливаются в тетанус, а сокращения низкопороговых медленных волокон, наоборот, тетанизируются. Данная структура регуляции мышечных сокращений обеспечивает, по крайней мере, два благоприятных аспекта, целесообразных для контроля двигательных действий в повседневной жизни. Во-первых, она оптимизирует комбинацию величины развиваемой силы и времени поддержания данной силы. Мотонейроны высокопороговых ДЕ не способны в течение продолжительного времени поддерживать относительно устойчивые разряды без значительного снижения их частоты, т.е. они быстро утомляются. Поэтому отсутствие тетанизации быстрых волокон обеспечивает способность генерировать силу и способность поддерживать ее более продолжительное время. Во-вторых, если механизм регуляции силы сокращения мышцы путем изменения частоты импульсации ДЕ наиболее выражен у медленных ДЕ, то возможности их частотного механизма в полной мере обеспечат выполнение повседневных низкоуровневых сокращений, не вовлекая дополнительное количество быстрых ДЕ.
Хотя причина синхронизации требует дальнейшего изучения, сам факт ее существования вызывает соблазн присвоить ей некоторую физиологическую и/или механическую ценность для регуляции мышечной силы. Первые попытки определить функциональную значимость синхронизации ДЕ показали, что уровень синхронизации между парами ДЕ в первой дорсальной межкостной мышце был наибольшим у тяжелоатлетов по сравнению с музыкантами и нетренированными лицами [39]. Cинхронизация может также увеличиваться после участия в программе силовой тренировки [40]. Но исследователям так и не удалось предоставить убедительных эмпирических доказательств того, что увеличение синхронизации само по себе вызывает увеличение мышечной силы. Напротив, в экспериментальных условиях [16, 41] и условиях компьютерной симуляции ЭМГ и моделирования скелетной мышцы [42] зависимости между синхронизацией и флуктуациями силы не обнаружено. Характер временнóй взаимосвязи активности различных ДЕ не влиял на силу сокращения мышцы. Следует обратить внимание на то, почему синхронизация активности ДЕ может увеличиваться в мышцах, подвергнутых силовому тренингу. Наиболее вероятный ответ заключается в том, что синхронизация активности ДЕ играет важную роль при быстрых сокращениях или в начале любого сокращения, так как влияет на скорость развития силы. Естественно, что, чем больше совпадают сократительные циклы разных ДЕ в начале сокращения мышцы, тем быстрее нарастает сила [42].
В нашем исследовании в эксцентрическом режиме синхронность была несколько выше, чем в концентрическом в диапазоне исследуемых интенсивностей. Так как закономерной связи между генерацией силы и синхронностью ДЕ в предыдущих исследованиях обнаружено не было, можно поставить под сомнение гипотезу о том, что синхронность может как-то влиять на суммарный механический эффект и бо́льшее производство силы, неоднократно задокументированное во время эксцентрических сокращений. Вполне логично предположить, что, если в эксцентрическом режиме работает меньшее совокупное количество волокон и в подавляющем большинстве это быстрые ДЕ, обладающие сходным порогом рекрутирования и, следовательно, частотой импульсации, то данное “подобие” может увеличивать случайную синхронизацию импульсной активности ДЕ.
ЗАКЛЮЧЕНИЕ
Технологически получить биосигнал поверхностной ЭМГ просто, но интерференционная электромиограмма часто не имеет специфических признаков, напоминая хаотическую кривую. Однако применение серьезного математического аппарата для анализа ЭМГ позволяет делать серьезные выводы о состоянии и функционировании двигательной системы. В нашей работе впервые применен метод рекуррентного анализа для косвенной оценки степени синхронизации работы ДЕ в эксцентрическом и концентрическом режимах сокращения с интенсивностью от 25 до 100% от МПС. Полученные данные согласуются с работами последних лет по изучению механизмов управления движениями и подтверждают тот факт, что с увеличением интенсивности сокращения синхронизация ДЕ во времени снижается. В эксцентрическом режиме синхронность была несколько выше, чем в концентрическом в диапазоне исследуемых интенсивностей. Тем не менее, более высокая степень синхронизации, вероятно, не вносит вклад в высокое производство силы во время эксцентрических сокращений, поскольку предыдущие исследования не подтвердили связи между величиной синхронизации ДЕ и силой сокращения. Как справедливо заметили C.J. DeLuca и J.C. Kline [17, 43], синхронизация, вероятно, не несет конкретного физиологического смысла и является не более чем эпифеноменом паттерна импульсации ДЕ, зависящая от бесчисленных и до сих пор неизвестных нейронных взаимодействий.
Необходимо проведение дальнейших исследований с применением метода декомпозиции ЭМГ-сигнала в эксцентрическом режиме мышечного сокращения в широком диапазоне интенсивностей для понимания изучаемых физиологических процессов.
Этические нормы. Все исследования проведены в соответствии с принципами биомедицинской этики, сформулированными в Хельсинкской декларации 1964 г. и ее последующих обновлениях, и одобрены локальным биоэтическим комитетом Российского государственного университета физической культуры, спорта, молодежи и туризма “РГУФКСМиТ” (Москва).
Информированное согласие. Каждый участник исследования представил добровольное письменное информированное согласие, подписанное им после разъяснения ему потенциальных рисков и преимуществ, а также характера предстоящего исследования.
Финансирование работы. Авторы сообщают, что не получали никакого финансирования и все затраты на исследование были личными вкладами авторов.
Конфликт интересов. Авторы декларируют отсутствие явных и потенциальных конфликтов интересов, связанных с публикацией данной статьи.
Список литературы
Hahn D. Stretching the limits of maximal voluntary eccentric force production in vivo // J. Sport Health Sci. 2018. V. 7. № 3. P. 275.
Herzog W., Schappacher G., DuVall M. et al. Residual force enhancement following eccentric contractions: a new mechanism involving titin // Physiology. 2016. V. 31. № 4. P. 300.
Altenburg T.M., de Ruiter C.J., Verdijk P.W. et al. Vastuslateralis surface and single motor unit electromyography during shortening, lengthening and isometric contractions corrected for mode-dependent differences in force-generating capacity // Acta Physiologica. 2009. V. 196. № 3. P. 315.
Del Valle A., Thomas C.K. Firing rates of motor units during strong dynamic contractions // Muscle Nerve. 2005. V. 32. № 3. P. 316.
Friedmann-Bette B., Bauer T., Kinscherf R. et al. Effects of strength training with eccentric overload on muscle adaptation in male athletes // Eur. J. Appl. Physiol. 2010. V. 108. № 4. P. 821.
agaard P., Simonse E.B., Andersen J.K. et al. Neural inhibition during maximal eccentric and concentric quadriceps contraction: effects of resistance training // J. Appl. Physiol. V. 89. № 6. P. 2249
Semmler J.G., Kornatz K.W., Dinenno D.V. et al. Motor unit synchronisation is enhanced during slow lengthening contractions of a hand muscle // J. Physiol. 2002. V. 545. № 2. P. 681.
Del Santo F., Gellia F., Schmied A. Motor unit synchronous firing as revealed by determinism of surface myoelectric signal // J. Neurosci. Meth. 2006. V. 155. № 1. P. 116.
Мейгал А.Ю., Риссанен С.М., Зарипова Ю.Р. и др. Возможности, открываемые использованием нелинейных параметров поверхностной электромиограммы в диагностике заболеваний и состояний двигательной системы человека // Физиология человека. 2015. Т. 41. № 6. С. 119. Meigal A.Yu., Rissanenb S.M., Zaripova Yu.R. et al. Nonlinear parameters of surface electromyogram for diagnostics of neuromuscular disorders and normal conditions of the human motor system // Human Physiology. 2015. V. 41. № 6. P. 672.
Farina D., Fattorini L., Felici F. et al. Nonlinear surface EMG analysis to detect changes of motor unit conduction velocity and synchronization // J. Appl. Physiol. 2002. V. 93. № 5. P. 1753.
Fattorini L., Felici F., Filligoi G.C. et al. Influence of high motor unit synchronization levels on non-linear and spectral variables of the surface EMG // J. Neuroscience Methods. 2005. V. 143. № 2. P. 133.
Webber C.L., Marwan N. Recurrence Quantification Analysis: Theory and Best Practices / Understanding Complex Systems. Switzerland: Springer Series, 2015. 421 p.
Zbilut J.P., Webber C.L., Jr. Embeddings and delays as derived from quantification of recurrence plots // Physics Letters A. 1992. V. 171. № 3–4. P. 199.
Del Santo F., Gelli F., Mazzocchio R. et al. Recurrence quantification analysis of surface EMG detects changes in motor unit synchronization induced by recurrent inhibition // Exp. Brain Res. 2007. V. 178. № 3. P. 308.
Hollander D.B., Kraemer R.R., Kilpatric M.W. et al. Maximal eccentric and concentric strength discrepancies between young men and women for dynamic resistance exercise // J. Strength Cond. Res. 2007. V. 21. № 1. P. 34.
De Luca C.J., Roy A.M., Erim Z. Synchronization of motor-unit firings in several human muscles // J. Neurophysiol. 1993. V. 70. № 5. P. 2010.
Kline J.C., De Luca C.J. Synchronization of motor unit firings: an epiphenomenon of firing rate characteristics not common inputs // J. Neurophysiol. 2016. V. 115. № 1. P. 178.
Petersen E., Rostalski P. A comprehensive mathematical model of motor unit pool organization, surface electromyography and force generation // Front. Physiol. 2019. V. 10. P. 176.
Eccles J.C., Eccles R.M., lundberg A. The action potentials of the alpha motoneurones supplying fast and slow muscles // J. Physiol. 1958. V. 142. № 2. P. 275.
Tracy B.L., Maluf K.S., Stephenson J.L. et al. Variability of motor unit discharge and force fluctuations across a range of muscle forces in older adults // Muscle Nerve. 2005. V. 32. № 4. P. 533.
Barry B.K., Pascoe M.A., Jesunathadas M. et al. Rate coding is compressed but variability is unaltered for motor units in a hand muscle of old adults // J. Neurophysiol. 2007. V. 97. № 5. P. 3206.
Moritz C.T., Barry B.K., Pascoe M.A. et al. Discharge rate variability influences the variation in force fluctuations across the working range of a hand muscle // J. Neurophysiol. 2005. V. 93. № 5. P. 2449.
Seyffarth H. The Behavior of Motor-Units in Voluntary Contraction. University of Oslo; Jacob DybWads, Forlag Oslo, Norway, 1940. № 4. P. 1.
Person R.S., Kudina L.P. Discharge frequency and discharge pattern of human motor units during voluntary contraction of muscle // Electroencephalogr. Clin. Neurophysiol. 1972. V. 32. № 5. P. 471.
Tanji J., Kato M. Firing rate of individual motor units in voluntary contraction of abductor digitiminimi muscle in man // Exp. Neurol. 1973. V. 40. № 3. P. 771.
De Luca C.J., Contessa P.J. Hierarchical control of motor units in voluntary contractions // Neurophysiol. 2012. V. 107. № 1. P. 178.
De Luca C.J., Hostage E.C. Relationship between firing rate and recruitment threshold of motoneurons in voluntary isometric contractions // J. Neurophysiol. 2010. V. 104. № 2. P. 1034.
De Luca C.J., Le Fever R.S., Mc Cue M.P. et al. Behaviour of human motor units in different muscles during linearly varying contractions // J. Physiol. 1982. V. 329. P. 113.
Freund H.J., Büdingen H.J., Dietz V. et al. Activity of single motor units from human forearm muscles during voluntary isometric contractions // J. Neurophysiol. 1975. V. 38. № 4. P. 933.
Holobar A., Farina D., Gazzoni M. et al. Estimating motor unit discharge patterns from high-density surface electromyogram // Clin. Neurophysiol. 2009. V. 120. № 3. P. 551.
Kamen G., Sison S.V., Du C.C. et al. Motor unit discharge behavior in older adults during maximal-effort contractions // J. Appl. Physiol. 1995. V. 79. № 6. P. 1908.
Masakado Y. The firing pattern of motor units in the mono- and multidirectional muscle // Jpn. Rehabil. Med. 1991. V. 28. P. 703.
Kanosue K., Yoshida M., Akazawa K. et al. The number of active motor units and their firing rates in voluntary contraction of human brachialis muscle // J. Physiol. 1979. V. 29. № 4. P. 427.
Masakado Y., Akaboshi K., Nagata M. Motor unit firing behavior in slow and fast contractions of the first dorsal interosseous muscle of healthy men // Electroencephalogr. Clin. Neurophysiol. 1995. V. 97. № 6. P. 290.
McGill K.C., Lateva Z.C., Marateb H.R. EMGLAB: an interactive EMG decomposition program // J. Neurosci. Methods. 2005. V. 149. № 2. P. 121.
Monster A.W., Chan H. Isometric force production by motor units of extensor digitorumcommunis muscle in man // J. Neurophysiol. 1977. V. 40. № 6. P. 1432.
Stashuk D., de Bruin H. Automatic decomposition of selective needle-detected myoelectric signals // IEEE Trans. Biomed. Eng. 1988. V. 35. № 1. P. 1.
Westgaard R.H., De Luca C.J. Motor control of low-threshold motor units in the human trapezius muscle // J. Neurophysiol. 2001. V. 85. № 4. P. 1777.
Semmler J.G., Nordstrom M.A. Motor unit discharge and force tremor in skill- and strength-trained individuals // Exp. Brain Res. 1998. V. 119. № 1. P. 27.
Milner-Brown H.S., Stein R.B., Lee R.G. Synchronization of human motor units: possible roles of exercise and supraspinal reflexes // Electroencephalogr. Clin. Neurophysiol. 1975. V. 38. № 3. P. 245.
Contessa P., Adam A., De Luca C.J. Motor unit control and force fluctuation during fatigue // J. Appl. Physiol. 2009. V. 107. № 1. P. 235.
Yao W., Fuglevand R.J., Enoka R.M. Motor-unit synchronization increases EMG amplitude and decreases force steadiness of simulated contractions // J. Neurophysiol. 2000. V. 83. № 1. P. 441.
De Luca C.J., Kline J.C. Statistically rigorous calculations do not support common input and long-term synchronization of motor-unit firings // J. Neurophysiol. 2014. V. 112. № 11. P. 2729.
Дополнительные материалы отсутствуют.
Инструменты
Физиология человека