

Физиология человека, 2021, T. 47, № 3, стр. 13-19
Влияние водной иммерсии обезьян на активность структур заднетеменной коры, участвующих в планировании и коррекции движений руки при выполнении моторной задачи
А. М. Бадаква 1, *, Н. В. Миллер 1, **, Л. Н. Зобова 1, В. Ю. Рощин 1, 2
1 ФГБУН ГНЦ РФ – Институт медико-биологических проблем РАН
Москва, Россия
2 ФГБУН ГНЦ РФ – Институт высшей нервной деятельности и нейрофизиологии РАН
Москва, Россия
* E-mail: ambadakva@mail.ru
** E-mail: nvmiller@mail.ru
Поступила в редакцию 02.12.2020
После доработки 12.12.2020
Принята к публикации 24.12.2020
Аннотация
Для оценки влияния опорной разгрузки на интенсивность нейрональной активности (НА) в областях заднетеменной коры (PPC), связанных с планированием и коррекцией целенаправленных движений руки (5d, PRR, 7a и LIP), у двух обезьян с фиксированной головой зарегистрирована НА исследуемых областей коры при выполнении задачи перемещения курсора на периферическую мишень с помощью джойстика до и после 3-часовой водной иммерсии. Фиксация головы у животных исключала влияние изменений вестибулярного входа на получаемые результаты. Полученные данные подтвердили предположение о влиянии опорной разгрузки на планирование и коррекцию движений руки при выполнении использованной моторной задачи, проявившиеся в снижении точности движения курсора и изменении интенсивности НА в трех областях PPC. Однако в связи с новыми данными о доминировании чувствительности области 7а (традиционно рассматриваемой как зрительной) к вестибулярному входу, вплоть до торможения зрительного входа, более важным является вывод о необходимости предварительного изучения различных вариантов взаимодействия вестибулярного, зрительного и проприоцептивного входов в областях коры, связанных с целенаправленными движениями руки – от их содействия до торможения одного сигнала другим.
Выраженное влияние гравитационной разгрузки на механизмы управления движениями, связанное с изменением состояния сенсорных систем и их участия в обеспечении активности центральных нервных механизмов, показано в многочисленных исследованиях, проведенных на человеке и животных как в космических полетах (КП), так и в модельных наземных исследованиях [1–4]. Ранее нами было обнаружено [4], что опорная разгрузка в условиях трехчасовой водной иммерсии не оказывает влияния на функциональное состояние нейронов представительства руки в первичной моторной коре (М1) у обезьян, оцениваемое с помощью измерения пороговых значений тока микростимуляции [5] (устраняющего и замещающего естественную активность и вызывающего движение руки с пространственно устойчивой конечной точкой [6]). Вместе с тем, опорная разгрузка изменяла паттерны движений курсора во время выполнения моторной задачи его перемещения с помощью джойстика, что свидетельствовало об изменении активности структур заднетеменной коры (PPC), участвующих в планировании и коррекции движений руки при выполнении моторной задачи [7]. Для более прямого подтверждения воздействия опорной разгрузки на функциональные субобласти PPC с различной ролью в сенсомоторной трансформации [7] необходимо провести регистрацию нейрональной активности (НА) в этих областях (5d, PRR, 7a и LIP) во время выполнения моторной задачи обезьянами до и после иммерсионного воздействия и оценить изменения в их взаимодействии.
МЕТОДИКА
Исследования проводили на двух обезьянах Macaca mulatta (А и Б) массой 4 и 6 кг. Предварительно обезьян готовили к иммерсионному исследованию, включая операцию установки технологического кольца на черепе животного и обучение их выполнению зрительно-двигательной задачи управления движением курсора на экране компьютерного дисплея к периферической мишени с помощью джойстика.
Зрительно-двигательная задача заключалась в попадании курсором, управляемым с помощью компьютерного двухкоординатного джойстика, в цель, предъявляемую в одном из 8 фиксированных мест на периферии экрана по случайному выбору, за ограниченное время (10 с). Обезьяна с жестко закрепенной за технологическое кольцо головой сидела в приматологическом кресле перед экраном компьютера. Каждое движение начиналось из центра экрана: цель предъявляли после того, как обезьяна приводила курсор к центру или отпускала его. При удачном попадании в цель обезьяна получала подкрепление в виде капли сока. Продолжительность экспериментальной сессии составляла 60 мин. Процедуры подготовки и тренировки обезьян описаны в работе [4]. Отражение положения курсора на экране производили каждые 40 мс (25 кадров в секунду) с латентностью не более 30 мс, что должно было обеспечивать ощущение адекватной управляемости и непрерывности движения курсора.
Опорную разгрузку в условиях невесомости моделировали оригинальным методом водной иммерсии обезьян, описанным в работе [4].
Для регистрации активности нейронов коры изготовляли и использовали вольфрамовые микроэлектроды в стеклянной изоляции с сопротивлением кончика в диапазоне от 0.5 до 2.0 МОм на частоте 1 кГц, помещенные в металлическую направляющую канюлю с внешним диаметром 0.6 мм. Для введения электрода в мозг в костях черепа сверлили вручную канал, направляя сверло диаметром 0.6 мм с помощью отверстий в съемной стереотаксической пластине. Электрод в направляющей канюле вводили через образовавшийся канал до твердой мозговой оболочки. Перемещение микроэлектрода в коре с помощью микроподачи и выбор позиции регистрации осуществляли при визуальном и аудио-контроле НА, а также наблюдении за движениями обезьяны.
Область 5d, которая расположена каудально по отношению к первичной соматосенсорной коре (S1) и медиально по отношению к интрапариетальной борозде (IPS), вовлечена в обработку пространственной информации о положении конечностей и в управление достигающими движениями руки [8]. Многие нейроны области 5d показывают выраженные паттерны разрядов, которые непрерывно изменяются согласованно с траекторией движения управляемого рукой курсора [9]. Существенно, что области 5d левого и правого полушарий имеют выраженные каллозальные связи, проявляющиеся в ответе нейронов 5d одного полушария на стимуляцию рецептивного поля другого полушария и облегчающие картирование области 5d [10].
Область PRR лежит медиально и дорсально по отношению к LIP и включает в себя структуры, клетки которых более активны при достигающих движениях руки, чем при саккадических движениях глаз [7]. При планировании достигающих движений руки в PRR учитываются не только взаимное расположение текущей позиции руки и цели, но и кинематические характеристики высокого уровня, такие как параметры траектории движения [4].
У обезьян область 7а расположена в выпуклости нижней париетальной извилины. Многочисленными исследованиями было установлено прямое участие этой области в обработке сигналов зрительного входа, в частности, представленных в них пространственных характеристик [10]. Оценка НА в области 7а может быть использована для определения позиций зрительных ориентиров и расположения, относительно них, наблюдателя. Такая обработка важна в навигации и пространственной ориентации [11].
Область LIP расположена непосредственно медиальнее области 7а в латеральном склоне IPS [12]. Обычно LIP рассматривают как единую область, но на самом деле она состоит из двух анатомически различных подразделений: дорсальной LIP (LIPd) и вентральной LIP (LIPv) [13]. LIPd в большей мере вовлечена в обработку зрительных сигналов, тогда как LIPv больше связана со зрительно вызванными саккадами [14].
Определение локализации областей 5d, PRR, 7a и LIP в соматосенсорной коре обезьяны предполагало предварительное нахождение представительства руки в М1 и центральной борозды. Картирование представительства руки в М1 осуществляли в области с соответствующими стереотаксическими координатами по атласу мозга обезьяны с помощью бифазной микростимуляции (с амплитудой до 110 мкА, длительностью импульса 0.2 мс, частотой повторения бифазной комбинации 200 Гц и общей длительностью 500 мс) и наблюдения за выраженными длительными движениями предплечья, запястья и пальцев обезьяны, возникающими при таких параметрах стимуляции [6]. Делая треки более каудально, находили точки, в которых моторные ответы руки на микростимуляцию наблюдали на глубине вплоть до 5–6 мм, что означало нахождение кончика электрода в переднем склоне центральной борозды (CS). Отсутствие моторных реакций на микростимуляцию при наличии НА на большой глубине в еще более каудальных точках означало нахождение трека в заднем склоне CS и переход в область первичной соматосенсорной коры (S1). Картирование представительства руки в S1 осуществляли, регистрируя ответы НА на стимуляцию кожи и шерсти плеча, предплечья, запястья, ладони и пальцев контрлатеральной руки обезьяны с помощью волосяной кисточки. Регистрируя НА по глубине трека прохождения электрода и смещаясь в каудальном направлении, выявляли поверхность зацентральной извилины, где активность прекращалась на глубине порядка 2–3 мм (достижение белого вещества) вплоть до начала переднего склона IPS (глубина регистрации 4–6 мм). При этом регистрировали НА во время стимуляции кожи и шерсти плеча, предплечья, запястья, ладони и пальцев обезьяны не только контрлатеральной руки, но и ипсилатеральной. Модуляция НА при стимуляции как левой, так и правой рук, означала нахождение электрода в области 5d. При более каудальном и глубоком погружении электрода вдоль переднего склона IPS находили область PRR, популяционная активность нейронов которой увеличивалась в связи с движением руки при перемещении курсора к мишени во время выполнения двигательной задачи.
После достижения переднего склона IPS делали также треки на 1–2 мм каудальнее, определяли трек, где активность нейронов реагировала на зрительный вход и прекращалась на глубине порядка 2–3 мм (достижение белого вещества), принимая этот трек как принадлежащий области 7а. Отступив на 1 мм медиальнее и пошагово возвращаясь в ростральном направлении, находили трек с выраженной реакцией нейронов на саккаду глаз при глубине погружения электрода 6–8 мм. Этот трек принимали как принадлежащий вентральному подразделению области LIP (LIPv).
Рис. 1 иллюстрирует динамику НА в исследуемых областях PPC у обеих обезьян во время выполнения моторной задачи. Для оценки текущего значения частоты высокоамплитудных спайков, их выделяли из сигнала НА с помощью пороговой детекции в программе Spike2, а затем подсчитывали их количество в скользящем окне длительностью 400 мс и делили на длительность окна. Последовательность полученных значений частоты спайков отражала участие нейронов исследуемой области в планировании и коррекции движений руки, управляющей курсором с помощью джойстика во время выполнения моторной задачи.
Рис. 1.
Фрагменты данных, полученных у обезьян (А) и (Б) в четырех областях PPC – 5d, PRR, 7a и LIP. На каждом из рисунков сверху вниз: текущая частота спайков; сигнал НА; метка событий (a – момент предъявления периферической мишени в различных случайных положениях; б – момент попадания курсора на мишень). Масштаб сигналов по осям показан масштабирующими отрезками.
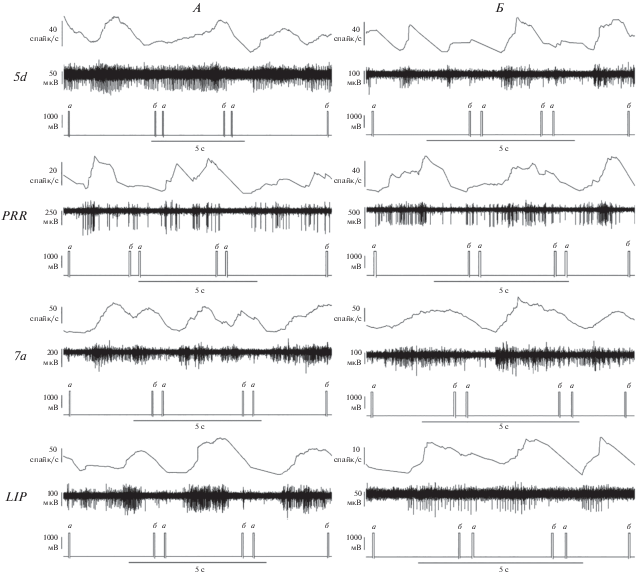
При обработке данных, полученных при выполнении обезьянами моторной задачи до и после иммерсии, для каждой удачной попытки попадания в цель производили визуализацию записей НА, привязанной к фазам движения курсора к цели, что позволяло исключить из анализа попытки, содержащие помехи в записях НА или сложные траектории движения к цели, свидетельствующие об отвлечении обезьяны от выполнения задачи.
В качестве индекса интенсивности НА при движении к цели для каждой анализируемой попытки вычислялось среднеквадратичное значение сигнала НА (НАСК) в полосе частот 250–600 Гц, усредненное по временному окну длительностью 500 мс, расположенному симметрично относительно точки траектории, в которой достигалась максимальная для данной попытки линейная скорость курсора.
Для оценки кривизны траектории движения курсора к цели для каждой анализируемой попытки вычисляли среднее по тому же временному окну значение косинуса угла между текущим направлением движения курсора и текущим направлением на мишень (cos α), рассчитываемого с временным шагом в 40 мс [4]. Эта косинусная мера наглядно иллюстрирует интегральное “качество” траекторий движения к цели: значение 1 соответствует прямолинейному движению на цель, значения, близкие к 0, свидетельствуют о хаотичности движений, в среднем не обеспечивающих приближение к цели. В процессе многомесячного обучения обезьян выполнению данной зрительно-двигательной задачи значение этого показателя для траекторий успешных попыток попадания в цель изменялось от 0.3–0.4, стабилизируясь на уровне 0.7–0.9. Последние значения соответствуют преимущественно устойчивым криволинейным траекториям, приводящим к цели, что являлось критерием достаточной тренированности обезьян. Криволинейность траекторий, специфическая как по отношению к цели, так и к конкретной обезьяне, не отражала поискового характера движений, о чем свидетельствовала одномодальность формы профиля скорости вдоль траектории. Более детальное изучение этой проблемы не входило в задачи данного исследования.
Далее вычислялись средние значения этих показателей (средних по окну cos α и НАСК) для отобранных движений к каждой цели и средние для всех целей. Достоверность анализируемых различий оценивали с помощью параметрического t-критерия Стьюдента, принимая критический уровень достоверности равным 0.01.
РЕЗУЛЬТАТЫ ИССЛЕДОВАНИЯ И ИХ ОБСУЖДЕНИЕ
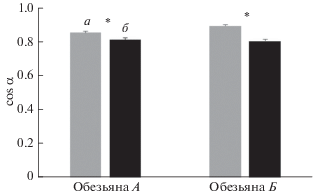
За одну экспериментальную сессию продолжительностью 60 мин тренированные обезьяны выполняли от 300 до 600 удачных попыток попадания в цель, из которых для дальнейшего анализа исключали около 30% попыток, содержащих помехи в записях НА или сложные траектории движения к цели, свидетельствующие об отвлечении обезьяны от выполнения задачи. На рис. 2 показаны изменения усредненных значений cos α (M ± sd) при движении к каждой из 8 мишеней за время сеанса выполнения моторной задачи после иммерсии у обезьяны А. Несмотря на то, что при движении к двум из восьми мишеней достоверных изменений не наблюдалось, для остальных мишеней изменения были однонаправлены и достоверны – значение показателя уменьшалось после воздействия иммерсии, что говорит об ухудшении точности движения руки к мишени. Об этом же свидетельствует (рис. 3) уменьшение усредненного для всех мишеней значения косинуса α после иммерсии как у обезьяны А с 0.851 ± 0.007 до 0.810 ± 0.008, так и у обезьяны Б с 0.890 ± 0.007 до 0.800 ± 0.010.
Рис. 2.
Изменения усредненных значений cos α (M ± sd) при движении курсора к каждой из 8 мишеней за время сеанса предъявления моторной задачи после иммерсии у обезьяны А. а – до иммерсии, б – после иммерсии. * – достоверное отличие (p < 0.01; количество степеней свободы n = 110, 104, 60, 109, 47, 85 для мишеней 1, 2, 3, 4, 7, 8 соответственно).

Рис. 3.
Изменения усредненных по 8 мишеням за время сеанса выполнения моторной задачи значений cos α (M ± sd), после иммерсии у обезьян А и Б. а – до иммерсии, б – после иммерсии. Звездочка обозначает достоверное отличие (p < 0.01; n = 687 и 752 для обезьян А и Б соответственно).
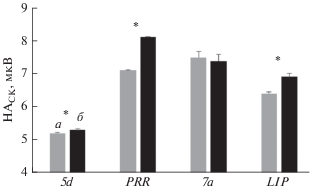
Подобным образом было оценено влияние иммерсии на изменение НАСК в четырех областях PPC: 5d, PRR, 7a и LIP. На рис. 4 показано изменение НАСК в области 5d у обезьяны А при движении к 8 мишеням. При движении в некоторых направлениях изменения были незначительны и не достоверны, однако при движении в других направлениях НАСК достоверно увеличилась. Усреднение показателя по всем мишеням за сеанс выполнения моторной задачи показал также достоверное его увеличение в области 5d после иммерсии; в областях PRR и LIP также наблюдали достоверное увеличение НАСК, в то время как в области 7а изменение НАСК не было достоверным (рис. 5). Аналогичные данные были получены на обезьяне Б.
Рис. 4.
Изменение усредненного значения НАСК (М ± sd) в области 5d при движении курсора к каждой из 8 мишеней за время сеанса предъявления моторной задачи после иммерсии у обезьяны А. а – до иммерсии, б – после иммерсии. * – достоверное отличие (p < 0.01; n = 110, 104, 60, 109, 47, 85 для мишеней 1, 2, 3, 4, 7, 8 соответственно).
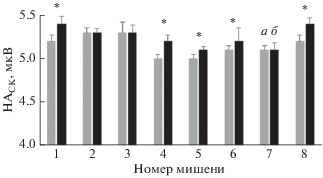
Рис. 5.
Изменения усредненного по 8 мишеням значения НАСК (М ± sd) за сеанс выполнения моторной задачи в четырех областях PPC до и после иммерсии у обезьяны А. а – до иммерсии, б – после иммерсии. * – достоверное отличие (p < 0.01; n = 687 и 752 для обезьян А и Б соответственно).
Таким образом, изменение точности траекторий движения курсора к мишеням под влиянием водной иммерсии свидетельствует о воздействии опорной разгрузки на планирование и коррекцию движений руки при выполнении моторной задачи, что предполагает изменение активности структур PPC. Вместе с тем, при запланированном анализе влияния иммерсии на взаимодействие исследованных структур PPC обезьян (5d, PRR, 7а и LIP) во время выполнения двигательной задачи возникли неожиданные трудности.
В исследовании, проведенном M. Gallaher et al. [3] на людях с участием 26-и молодых испытуемых, было показано, что смена ориентации тела испытуемых по отношению к вектору земной гравитации приводит к быстрым изменениям в стратегии двигательного поведения, что авторы исследования связывают с важной ролью гравитационной информации в планировании и выполнении движений. Поэтому использование фиксации головы обезьян во время наших исследований, исключающей влияние изменений вестибулярного входа на получаемые результаты, является, казалось бы, оправданным. Однако при углубленном (с использованием обзора литературных данных последних лет) анализе влияния иммерсии на НА исследованных нами структур PPC обезьян (5d, PRR, 7а и LIP) во время выполнения двигательной задачи мы столкнулись с совершенно неожиданным фактом, изменившим наше отношение к полученным данным. В недавно опубликованных результатах исследования E. Avila et al. на обезьянах Macaca mulatta [15] была обнаружена чувствительность нейронов области 7а к вестибулярным входам, которая никогда ранее не тестировалась. Отчасти исследование E. Avila et al. было вызвано устным сообщением K. Kawano, единственным, описавшим вестибулярные ответы в области 7a, об ошибочности его работы. Традиционно область 7a рассматривалась как зрительная и вообще считалась выходом дорсальной зрительной иерархии [15].
В исследовании E. Avila et al. была использована движущаяся платформа с линейной и вращательной степенями свободы. Кроме того, была применена одновременная стимуляция зрительного входа (используя оптический поток) и реальное движение (используя платформу), чтобы изучить мультисенсорную конвергенцию в области 7а во время линейного движения.
По признанию авторов работы [15], результаты исследования были совершенно неожиданными. Основное, что они обнаружили – существование в области 7а популяций нейронов, независимо кодирующих зрительный и связанные с движением вестибулярные стимулы. Ответы на комбинированный зрительно-вестибулярный стимул имели тенденцию к доминированию вестибулярного входа. В некоторых случаях нейроны, отвечающие на селективный зрительный стимул, даже подавлялись, когда визуальное предъявление движения сочеталось с движением платформы. Тот факт, что вестибулярные входы в большей мере определяют нейрональные ответы в области 7a, означает, что локальные механизмы могут угнетать зрительные входы, связанные с движением, если доступны вестибулярные сигналы. Из этого следует, что полученные нами результаты о влиянии иммерсии на НА исследованных, тесно связанных между собой структур PPC обезьян при жесткой фиксации головы могут, быть неполными из-за исключения учета влияния вестибулярного входа.
ЗАКЛЮЧЕНИЕ
Результаты исследования подтвердили влияние опорной разгрузки на планирование и коррекцию движений руки обезьяны при выполнении использованной моторной задачи. Однако, планируя и проводя исследование, мы не могли учитывать данные, опубликованные M. Gallaher et al. [3] и E. Avila et al. [15] о том, что как отсутствие вестибулярного входа, так и его наличие существенно влияет на совместную активность исследуемых структур PPC обезьян во время выполнения двигательной задачи. Хотя в настоящее время существуют общепринятые представления о распределении вестибулярных входов в коре головного мозга, основанные на многолетних исследованиях различных авторских коллективов [16], после публикации E. Avila et al. [15] наша оценка влияния иммерсии на НА структур PPC обезьян во время выполнения двигательной задачи требует, во-первых, более тщательного изучения имеющихся литературных данных об областях вестибулярного представительства в коре головного мозга и присутствии в них проекций других модальностей; во-вторых, учета различных вариантов взаимодействия вестибулярного и сенсорных входов других модальностей – от их конвергенции до угнетения одного сигнала другим.
Этические нормы. Все исследования проведены в соответствии с принципами биомедицинской этики, сформулированными в Хельсинкской декларации 1964 г. и ее последующих обновлениях, и одобрены Комиссией Института медико-биологических проблем РАН по биомедицинской этике (Москва), протокол № 476 от 28.06.2018 г.
Финансирование работы. Работа выполнена в рамках базовой темы РАН № 63.1.
Конфликт интересов. Авторы декларируют отсутствие явных и потенциальных конфликтов интересов, связанных с публикацией данной статьи.
Список литературы
Van Omberger A., Demertzi A., Tomilovskaya E. et al. The effect of spaceflight and microgravity on the human brain // J. Neurology. 2017. V. 264. Suppl 1. P. 18.
Tomilovskaya E., Shigueva T., Sayenko D. et al. Dry immersion as a ground-based model of microgravity physiological effects // Front. Physiol. 2019. V. 10. Article 284.
Gallagher M., Arshad I., Ferrè E.R. Gravity modulates behaviour control strategy // Exp. Brain Res. 2019. V. 237. № 4. P. 989.
Бадаква А.М., Миллер Н.В., Зобова Л.Н., Рощин В.Ю. Исследование влияния опорной разгрузки на корковые механизмы управления движениями руки в иммерсионных экспериментах на обезьянах // Авиакосмич. и экологич. мед. 2019. Т. 53. № 3. С. 33.
Бадаква А.М., Зобова Л.Н., Миллер Н.В., Рощин В.Ю. Воздействие длительной внутрикортикальной микростимуляции на моторную кору // Физиология человека. 2017. Т. 43. № 5. С. 129. Badakva A.M., Zobova L.N., Miller N.V., Roschin V.Yu. Influence of long-term intracortical microstimulation on the motor cortex // Human Physiology. 2017. V. 43. № 5. P. 601.
Graziano M.S.A. Ethological action maps: a paradigm shift for the motor cortex // Trends Cogn. Sci. 2016. V. 20. № 2. P. 121.
Cui H., Andersen R.A. Posterior parietal cortex encodes autonomously selected motor plans // Neuron. 2007. V. 56. № 3. P. 552.
Lacquaniti F., Guigon E., Bianchi L. et al. Representing spatial information for limb movement: role of area 5 in the monkey // Cereb. Cortex.1995. V. 5. № 5. P. 391.
Mulliken G.H., Musallam S., Andersen R.A. Forward estimation of movement state in posterior parietal cortex // PNAS. 2008. V. 105. № 24. P. 8170.
Krubitzer L., Disbrow E. The evolution of parietal areas involved in hand use in primates / The Senses. V. 6: Somatosensation. Eds. Gardner E., Kaas J. London: Elsevier, 2008. P. 183.
Torres E.B., Quian Quiroga R., Cui H., Buneo C. Neural correlates of learning and trajectory planning in the posterior parietal cortex // Front. Integr. Neurosci. 2013. V. 7. P. 39.
Chafee M.V., Crowe D.A. Thinking in spatial terms: decoupling spatial representation from sensorimotor control in monkey posterior parietal areas 7a and LIP // Front. Integr. Neurosci. 2013. V. 6. Article 112.
Patel G.H., Shulman G.L., Baker J.T. et al. Topographic organization of macaque area LIP // PNAS. 2010. V. 107. № 10. P. 4728.
Chen M., Li B., Guang J. et al. Two subdivisions of macaque LIP process visual-oculomotor information differently // PNAS. 2016. V. 113. № 41. P. E6263.
Avila E., Lakshminarasimhan K.J., DeAngelis G.C., Angelaki D.E. Visual and vestibular selectivity for self-motion in macaque posterior parietal area 7a // Cerebral Cortex. 2019. V. 29. № 9. P. 3932.
Cullen K.E. Vestibular processing during natural self-motion: implications for perception and action // Nat. Rev. Neurosci. 2019. V. 20. № 6. P. 346.
Дополнительные материалы отсутствуют.
Инструменты
Физиология человека