

Физиология человека, 2022, T. 48, № 2, стр. 86-98
Применение методов вейвлетного и рекуррентного анализа к исследованиям биоэлектрической активности мозга при умеренных когнитивных нарушениях
О. Е. Дик 1, *, А. Д. Ноздрачев 1
1 ФГБУН Институт физиологии имени И.П. Павлова РАН
Санкт-Петербург, Россия
* E-mail: dickviola@gmail.com
Поступила в редакцию 27.04.2021
После доработки 04.10.2021
Принята к публикации 27.10.2021
- EDN: QJPFDW
- DOI: 10.31857/S0131164622020059
Аннотация
Исследованы динамические изменения, происходящие в паттернах электроэнцефалограмм (ЭЭГ) пациентов с умеренными когнитивными нарушениями и количественные показатели этих паттернов, определенные с помощью методов вейвлетного и рекуррентного анализа до и после лечебных ритмических воздействий. Показано, что вычисленные показатели вейвлетных спектров и рекуррентных диаграмм (значения коэффициента усвоения ритма, процента рекуррентных точек, расположенных на диагональных линиях и коэффициента взаимной корреляции между вероятностями рекуррентностей светового сигнала и ответа мозга) очень чувствительны к изменениям в реактивных паттернах ЭЭГ в ответ на стимулирующие воздействия.
Сосудистые заболевания головного мозга являются частой причиной когнитивных расстройств у лиц старшего возраста [1–3]. При хронической цереброваскулярной недостаточности состояние когнитивных функций коррелирует с выраженностью поражения головного мозга [4]. Проявления когнитивных нарушений могут протекать от минимальной дисфункции до деменции [5]. Высокий уровень тревожности, стрессовые расстройства повышают риск развития когнитивных нарушений [6–9]. Значительная распространенность этих нарушений (более 40%) среди неврологических пациентов и сложность медикаметозного лечения обуславливает необходимость поиска безопасных немедикаметозных способов коррекции [7]. Одним из методов, основанных на неспецифических сенсорных импульсных воздействиях на организм человека с лечебно-диагностическими целями, является метод формирования стабильных функциональных связей мозга человека [10].
Следует отметить, что в современной нейрофизиологии выделяют три вида связей между структурами мозга: структурные (анатомические нейрональные сети), функциональные (обуславливающие корреляции между различными ансамблями нейронов) и эффективные (информационные потоки между ансамблями) [11]. Исследования паттернов функциональных связей, основанные на когерентности или корреляции между различными областями коры, продемонстрировали, что эти связи отражают основную структурную организацию анатомических связей [12]. Согласно работе [13], два основных принципа, сегрегация и интеграция, связывают эти различные способы взаимодействия мозга. Сегрегация относится к существованию специализированных областей мозга, организованных в отдельные нейрональные популяции, а интеграция порождает скоординированную активацию распределенных популяций нейронов, что способствует возникновению когерентных когнитивных и поведенческих состояний [13]. В настоящей работе под формированием функциональных связей мозга понимается создание функциональной связности в нейрональных структурах мозга в условиях ритмических световых воздействий, приводящих к переходу от патологического к оптимальному функциональному состоянию мозга [14].
Такие направленные воздействия играют важную роль для коррекции психофизиологической деятельности человека при различных заболеваниях органического и функционального генеза, а также для здоровых лиц в условиях эмоциональных перегрузок [15, 16]. Способность восстанавливать утраченные резервы психических процессов позволяет использовать этот метод в ситуациях, когда фармакотерапия имеет побочные действия на организм [15–17].
Известно, что восстановление функционального состояния мозга происходит у больных с отчетливой реакцией усвоения ритмической фотостимуляции в исходной электроэнцефалограмме (ЭЭГ) [18]. При этом выраженность перестроек электрической активности мозга в соответствии с частотой световых импульсов, определяется возбудимостью нейронов коры головного мозга [19]. Нейрофизиологические механизмы влияния ритмической фотостимуляции на мозг состоят в том, что такая стимуляция, по сути, манипулирует колебаниями мозга, захватывая эти колебания и обеспечивая фазовую синхронизацию ритмического стимула и отклика мозга на него [20, 21]. В связи с этим для объективного подтверждения положительной динамики функционального состояния в процессе лечения методом активации функциональных связей необходимо использовать анализ реактивных паттернов ЭЭГ как ответа мозга на разные частоты световых импульсов до и после курса лечения.
Современная математика предлагает множество методов линейного и нелинейного анализа сложных нестационарных сигналов, среди которых можно выделить методы вейвлетного и рекуррентного анализа, позволяющие оценить количественные характеристики ЭЭГ мозга [22, 23]. Ранее мы показали возможность применения методов вейвлетного и рекуррентного анализа к оценке изменения степени невротизации у пациентов с неврологическими нарушениями в виде панических атак после лечебных воздействий, направленных на формирование функциональных связей мозга во время ритмической фотостимуляции [24, 25].
Целью настоящей работы является исследование динамики паттернов ЭЭГ мозга человека при умеренных когнитивных нарушениях до и после сеансов активации функциональных связей мозга и определение динамических характеристик, которые могут служить объективными показателями улучшения функционального состояния нервной системы после этих сеансов. Для этого, при анализе совместных рекуррентных диаграмм ритмических световых сигналов и ответов мозга на эти сигналы, в данной работе используется дополнительный, кроме рассмотренных ранее, показатель этих диаграмм, такой как коэффициент взаимной корреляции между вероятностями рекуррентностей светового сигнала и ответа мозга на определенные частоты.
МЕТОДИКА
Исследовали характеристики ЭЭГ мозга 30 женщин в возрасте от 65 до 75 лет (в среднем 69 лет), обратившихся в связи с ухудшением памяти в клинику Института мозга человека имени Н.П. Бехтеревой РАН (г. Санкт-Петербург). По краткой шкале оценки психического статуса MMSE (27.5 ± ± 0.4 балла) у этих женщин наблюдали умеренные когнитивные нарушения, а по данным магнитно-резонансной томографии (МРТ) отмечали умеренные нарушения функционального состояния мозга, соответствующие сосудистой энцефалопатии. ЭЭГ регистрировали до и после лечебных сеансов, проводимых в клинике Института мозга человека имени Н.П. Бехтеревой РАН. Курс 11 лечебных сеансов состоял в 1 сеансе формирования и последующих 10 сеансах активации функциональных связей мозга во время ритмической фотостимуляции. Сеанс их формирования включал в себя фотостимуляцию длительностью 10 с на фоне действия однократного приема фармакологического препарата этимизола в дозировке 0.01 мг. Однократный прием данного препарата в такой дозировке не может оказывать значимых лечебных когнитивно-положительных эффектов [14]. При таком использовании этимизол выполняет функцию неспецифического коннектора, способствующего фиксации следов памяти [10]. На следующие сутки проводили сеанс активации, состоящий из 6 серий фотостимуляций тех же частот без дополнительных фармакологических воздействий. Длительность 1 серии составляла 10 с. Интервалы между сериями – 1 мин. Во время проведения сеансов больные находились в комфортной позе, лежа с закрытыми глазами. Фотостимуляции проводили с помощью “тренажера функциональной активности мозга ТММ Мираж”, который является представителем линии приборов световой стимуляции (ООО НПП “МедПАСС”, Россия). Данный прибор активно используется в программах немедикаментозной коррекции при различных неврологических нарушениях [15–17].
ЭЭГ регистрировали до начала курса и через неделю после него с помощью 21-канального электроэнцефаллографа в состоянии покоя и при ритмической фотостимуляции. Активные электроды располагали по стандартной схеме 10 /20. В качестве индифферентного использовали усредненный электрод (Av). Ритмическая фотостимуляция представляла собой последовательность двенадцатисекундных серий световых импульсов с разной частотой с интервалом 30 с между сериями. Ответы мозга регистрировали в затылочных локусах O1, O2, где реакции мозга на световой стимул имеют наибольшую амплитуду. Частота дискретизации составляла 256 Гц.
Предоставленные клиникой Института мозга человека имени Н.П. Бехтеревой РАН данные, отфильтровывали в диапазоне 1.5–25 Гц и после удаления артефактов анализировали с помощью метода непрерывного вейвлет-преобразования [22] и метода анализа совместных рекуррентностей ЭЭГ и светового сигнала [23]. Применяли разработанное собственное матобеспечение и пакеты программ Wavelet Toolbox MATLAB, а также процедуры CRP Toolbox, доступные на сайте tocsy.pik-potsdam.de/crp.php.
В качестве базисного вейвлета использовали комплексный вейвлет Морле:
(1)
${{\psi }_{0}}(t) = {{\pi }^{{{{ - 1} \mathord{\left/ {\vphantom {{ - 1} 4}} \right. \kern-0em} 4}}}}\exp ( - 0.5{\kern 1pt} {{t}^{2}})\exp (i{{\omega }_{0}}t){\text{,}}$(2)
$f = {{\left( {{{\omega }_{0}} + \sqrt {2 + {{\omega }_{0}}} } \right)} \mathord{\left/ {\vphantom {{\left( {{{\omega }_{0}} + \sqrt {2 + {{\omega }_{0}}} } \right)} {4\pi a}}} \right. \kern-0em} {4\pi a}} \approx {1 \mathord{\left/ {\vphantom {1 {a.}}} \right. \kern-0em} {a.}}$(3)
$\begin{gathered} W(f,{{t}_{0}}) = {{\pi }^{{{{ - 1} \mathord{\left/ {\vphantom {{ - 1} 4}} \right. \kern-0em} 4}}}}\sqrt f \int\limits_{ - \infty }^{ + \infty } {x(t){\text{ exp}}} {{\left( { - 0.5(t - {{t}_{0}}} \right)}^{2}}{{f}^{2}}) \times \\ \times \,\,\exp ( - 2\pi i(t - {{t}_{0}})f)dt, \\ \end{gathered} $Величина квадрата модуля вейвлет-преобразования |W(f, t0)|2 определяет мгновенное распределение энергии паттерна ЭЭГ по частотам f (локальный вейвлетный спектр в момент времени t0). Интеграл от локального вейвлетного спектра
задает глобальный вейвлетный спектр (интегральное распределение энергии вейвлетного спектра сигнала по частотам в интервале времени [t1, t2]).Интегрирование по частоте
(5)
$E({{t}_{0}}) = \int\limits_{{{f}_{{C{\kern 1pt} }}} - {\kern 1pt} \Delta {\kern 1pt} f}^{{{f}_{C}}{\kern 1pt} + {\kern 1pt} \Delta {\kern 1pt} f} {{{{\left| {W(f,{{t}_{0}})} \right|}}^{2}}df} $Световой сигнал задавался последовательностью k импульсов, следующих друг за другом с частотой fC:
(6)
$p(t) = \sum\limits_{j{\kern 1pt} = {\kern 1pt} 0}^{k{\kern 1pt} - {\kern 1pt} 1} {\frac{{0.5}}{{{{r}_{0}}\sqrt \pi }}} \exp \left( { - \frac{{{{{(t - {{t}_{j}})}}^{2}}}}{{4r_{0}^{2}}}} \right),$Реакцию усвоения частоты фотостимуляции в паттернах ЭЭГ оценивали по величине коэффициента усвоения (kR) в исследуемом частотном диапазоне [fС – Δ f, fС + Δ f], где Δ f = 0.5 Гц, и по нарастанию энергии вейвлетного спектра непосредственно в период фотостимуляции [27].
Коэффициент усвоения ритма (kR) определяли как отношение максимумов глобальных вейвлетных спектров во время и до действия фотостимула:
(7)
${{k}_{R}} = {{{{E}_{{\max }}}{{{(f)}}_{{{\text{during}}}}}} \mathord{\left/ {\vphantom {{{{E}_{{\max }}}{{{(f)}}_{{{\text{during}}}}}} {{{E}_{{\max }}}{{{(f)}}_{{{\text{before}}}}}}}} \right. \kern-0em} {{{E}_{{\max }}}{{{(f)}}_{{{\text{before}}}}}}}{\text{.}}$Метод анализа совместных рекуррентностей ЭЭГ и светового сигнала основан на оценке совместных рекуррентных диаграмм паттерна ЭЭГ и светового сигнала.
Совместная рекуррентная диаграмма представляет собой графическое представление матрицы:
(8)
${{R}_{{i,{\kern 1pt} j}}}(\varepsilon ) = \left\{ \begin{gathered} 1,\,\,\,\,{{y}_{i}} \approx {{y}_{j}},\,\,\,{{z}_{i}} \approx {{z}_{j}} \hfill \\ 0,\,\,\,\,{{y}_{i}} \ne {{y}_{j}},\,\,\,\,{{z}_{i}} \ne {{z}_{j}}, \hfill \\ \end{gathered} \right.$Фазовые траектории состояний z(t) и y(t) были получены из исходных временны́х рядов {x(t)} и {p(t)} методом временны́х задержек [29]:
где d – временнáя задержка, m – размерность вложения (минимальная размерность пространства, в котором восстановленная траектория воспроизводит свойства исходной траектории). Оптимальная временнáя задержка d находилась на основании поиска первого минимума функции взаимной информации [30]. Оптимальную размерность вложения m определяли методом поиска минимума ближайших ложных соседей [31]. Величину ε-ошибки выбирали равной 1% от величины стандартного отклонения анализируемого паттерна ЭЭГ.Реакцию усвоения ритма в паттернах ЭЭГ методом анализа совместных рекуррентностей оценивали по показателю рекуррентной диаграммы, называемому показателем детерминизма и определяемому как отношение числа рекуррентных точек, составляющих диагональные структуры, к общему числу рекуррентных точек:
(10)
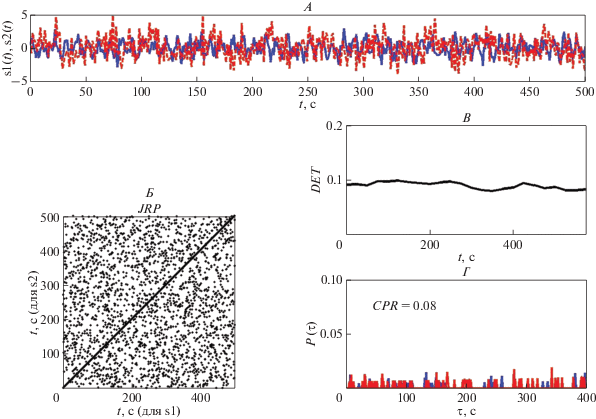
$DET = {{\sum\limits_{l{\kern 1pt} = {\kern 1pt} {{l}_{{\min }}}}^N {lP(\varepsilon ,l)} } \mathord{\left/ {\vphantom {{\sum\limits_{l{\kern 1pt} = {\kern 1pt} {{l}_{{\min }}}}^N {lP(\varepsilon ,l)} } {\sum\limits_{i,{\kern 1pt} j}^N {{{R}_{{i,{\kern 1pt} j}}}(m,\varepsilon )} }}} \right. \kern-0em} {\sum\limits_{i,{\kern 1pt} j}^N {{{R}_{{i,{\kern 1pt} j}}}(m,\varepsilon )} }},$Длинные диагонали выявляют аналогичную эволюцию во времени в двух колебательных процессах, а короткие показывают уменьшение сходства динамики процессов. Поэтому для двух полностью стохастических процессов величина показателя DET будет близка к нулю, а для сходных детерминированных процессов эта величина будет близка к единице. На рис. 1 представлены периодические сигналы с одинаковым периодом, равным 50 с (рис. 1, А), построенная для этих сигналов рекуррентная диаграмма (рис. 1, Б) содержит длинные диагонали, находящиеся друг от друга на расстоянии, равном периоду колебаний, величина DET = 1 (рис. 1, В). Для двух полностью стохастических сигналов (рис. 2, А) совместная рекуррентная диаграмма не содержит длинных диагоналей, а имеет практически только изолированные точки (рис. 2, Б), величина DET = 0.1 (рис. 2, В). Таким образом, значение показателя DET, определяющего процент рекуррентных точек, расположенных на диагональных линиях совместной рекуррентной диаграммы, связано со сходством динамического поведения двух колебательных процессов (или отсутствием такого сходства). Однако для выяснения сходства динамического поведения реальных сложных сигналов этого показателя недостаточно. Тем более, что практически этот показатель показывает наличие некоторого сходства для значений DET, превышающих значение 0.8 [21, 23]. Кроме того, этот показатель не определяет такое явление, как фазовая синхронизация двух сигналов.
Рис. 1.
Периодические сигналы s1(t) и s2(t) с одинаковым периодом и фазой (А), совместная рекуррентная диаграмм этих сигналов (Б), изменения во времени показателя детерминизма (DET) (В) и полностью совпадающие вероятности рекуррентностей ${{P}_{1}}(\tau )$ и ${{P}_{2}}(\tau )$ этих сигналов (Г).
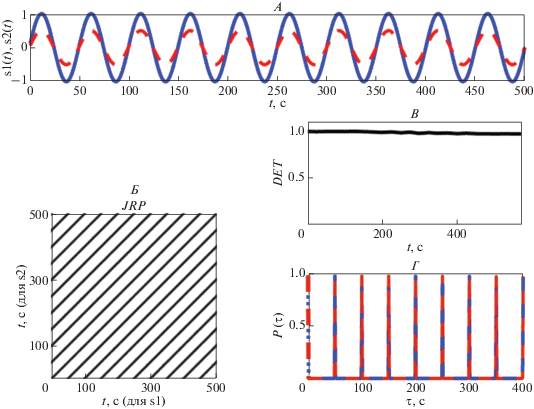
Рис. 2.
Совместная рекуррентная диаграмма (Б) для двух полностью стохастических сигналов (А), изменения во времени показателя детерминизма (DET) (В) и отсутствие совпадений вероятностей рекуррентностей ${{P}_{1}}(\tau )$ и ${{P}_{2}}(\tau )$ (Г).
В связи с этим для выявления отклика мозга на ритмическую фотостимуляцию в качестве показателя фазовой синхронизации между исходными временны́ми рядами {x(t)} и {p(t)} использовался коэффициент взаимной корреляции между вероятностями рекуррентностей ${{P}_{1}}(\tau )$ и ${{P}_{2}}(\tau )$ [32]:
(11)
$CPR = \frac{1}{{{{\sigma }_{1}}{{\sigma }_{2}}}}\sum\limits_{\tau {\kern 1pt} = {\kern 1pt} {{\tau }_{e}}}^{{N \mathord{\left/ {\vphantom {N 2}} \right. \kern-0em} 2}} {\left( {{{P}_{1}}(\tau ) - \overline {{{P}_{1}}(\tau )} } \right)\left( {{{P}_{2}}(\tau ) - \overline {{{P}_{2}}(\tau )} } \right)} ,$(12)
${{P}_{1}}(\tau ) = \frac{1}{{N - \tau }}\sum\limits_{i{\kern 1pt} = {\kern 1pt} 1}^{N{\kern 1pt} - {\kern 1pt} \tau } {{{R}_{{i,{\kern 1pt} j{\kern 1pt} + {\kern 1pt} \tau }}}} (\tau )$Совпадение максимумов вероятностей рекуррентностей ${{P}_{1}}(\tau )$ и ${{P}_{2}}(\tau )$ для двух фазовых траекторий z(t) и y(t) при одинаковых временах kτ, где k – целое число, означает наличие фазовой синхронизации двух исходных временны́х рядов {x(t)} и {p(t)} [33]. В этом случае значение коэффициента взаимной корреляции CPR близко к 1. Напротив, если два временны́х ряда не находятся в фазовой синхронизации, максимумы вероятностей рекуррентностей возникают неодновременно, и значение CPR близко к 0. Так, для полностью синхронизованных сигналов, представленных на рис. 1, А, значение CPR = 1, а для полностью стохастических сигналов, представленных на рис. 2, А, значение CPR = 0.08.
Для определения статистически значимых различий между средними значениями параметров, полученными до и после сеансов активации функциональных связей мозга, применяли однофакторный дисперсионный анализ ANOVA.
РЕЗУЛЬТАТЫ ИССЛЕДОВАНИЯ И ИХ ОБСУЖДЕНИЕ
В фоновых паттернах ЭЭГ большинства тестируемых пациентов преобладала низкоамплитудная полиморфная активность, свидетельствующая о выраженном снижении возбудимости корковых нейронов вследствие сосудистых нарушений головного мозга [34].
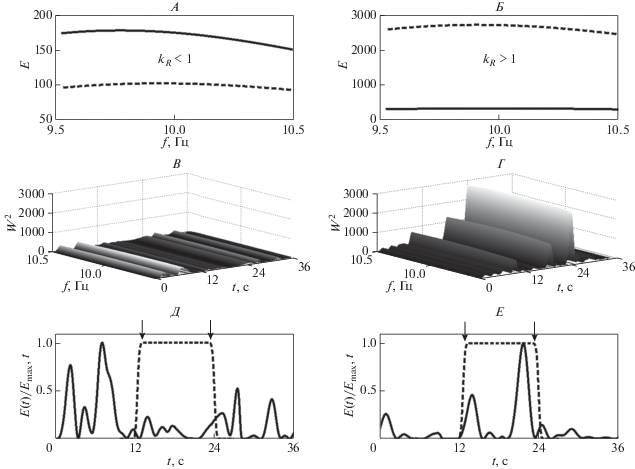
Примеры вейвлетных спектров реактивных паттернов ЭЭГ в ответ на фотостимуляцию частотой 10 Гц, полученных до и после сеансов активации функциональных связей мозга, представлены на рис. 3. Для паттерна ЭЭГ, зарегистрированного до этих сеансов, характерно следующее. Локальный вейвлетный спектр демонстрирует наличие максимумов на частотах анализируемого диапазона 10 ± 0.5 Гц в интервале времени, предшествующем времени фотостимуляции (рис. 3, Д). Нормированные интегральные распределения энергий вейвлетных спектров ${{E(t)} \mathord{\left/ {\vphantom {{E(t)} {{{E}_{{\max }}}(t)}}} \right. \kern-0em} {{{E}_{{\max }}}(t)}}$ паттерна ЭЭГ и светового сигнала (сплошная и штрихпунктирная линии, соответственно) показывают отсутствие нарастания энергии вейвлетного спектра ЭЭГ во время фотостимуляции (рис. 3, Д). Глобальный вейвлетный спектр E(f) этого паттерна во время фотостимуляции также имеет значения, меньшие, чем до фотостимуляции (рис. 3, А). Соответственно, значение коэффициента усвоения ритма предложенной частоты kR < 1. В отличие от этого после сеансов активации функциональных связей мозга нарастание энергии вейвлетного спектра ЭЭГ происходит непосредственно в период действия фотостимула (рис. 3, Е), поэтому максимумы энергии локального вейвлетного спектра находятся в интервале времени, соответствующем времени действия стимула (рис. 3, Г). Отношение максимумов глобальных вейвлетных спектров во время и до действия фотостимула определяет значение коэффициента усвоения ритма kR > 1 (рис. 3, Б). Таким образом, в анализируемых паттернах ЭЭГ, соответствующих состоянию до активации функциональных связей мозга, усвоение частоты 10 Гц отсутствует, а после сеансов активации наблюдается реакция усвоения этой частоты.
Рис. 3.
Oтсутствие реакции усвоения частоты 10 Гц в паттерне ЭЭГ (отведение О1) до сеансов активации функциональных связей мозга (А, В, Д) и возникновение реакции усвоения после сеансов (Б, Г, Е). Глобальные вейвлетные спектры паттернов ЭЭГ (А, Б) до фотостимуляции (сплошная линия), во время фотостимуляции (штрихпунктирная линия). Нормированные интегральные распределения вейвлетных спектров ЭЭГ и световых стимулов (сплошные и штрихпунктирные линии) (Д, Е). Начало и конец фотостимуляции показаны вертикальными стрелками.
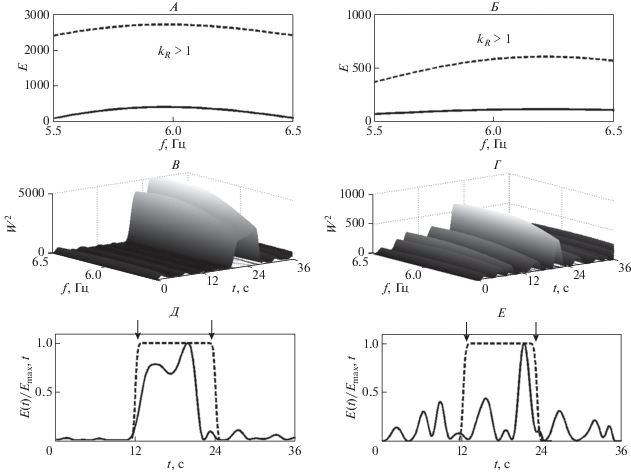
Для реактивных паттернов ЭЭГ в ответ на фотостимуляцию частотой 6 Гц, полученных до и после лечебных сеансов, характерно наличие реакции усвоения (рис. 4). Об этом свидетельствует превышение максимумов глобальных вейвлетных спектров во время действия фотостимула, по сравнению с величинами этих максимумов до фотостимуляции, kR > 1 в обоих случаях (рис. 4, А, Б), а также нарастание энергии вейвлетных спектров ЭЭГ именно во время фотостимуляции (рис. 4, В–Е).
Рис. 4.
Наличие реакции усвоения частоты 6 Гц в паттерне ЭЭГ (отведение О2) до сеансов активации функциональных связей мозга (А, В, Д) и после сеансов (Б, Г, Е). Обозначения см. рис. 3.
Динамика изменения усвоения ритма в паттернах ЭЭГ у пациентов с умеренными когнитивными нарушениями после проводимых сеансов подтверждалась также изменением одновременных рекуррентностей в совместных рекуррентных диаграммах ЭЭГ и ритмических световых сигналов определенных частот.
На рис. 5, А, Б представлены фотостимул частотой 10 Гц и фрагменты паттерна ЭЭГ длительностью 3 с во время фотостимуляции. Совместные рекуррентные диаграммы для светового сигнала и фрагментов ответов мозга до и после сеансов активации функциональных связей приведены на рис. 5, В, Г, соответственно. Эти диаграммы построены при величине временнóй задержки d = 3 и размерности вложения m = 3. Совместная рекуррентная диаграмма, полученная до сеансов (рис. 5, В), содержит очень короткие диагональные линии, локализующиеся в окрестности всегда существующей линии, проходящей под углом в 45°. Это соответствует близкому к нулю значению показателя DET и свидетельствует об отсутствии сходства динамики данного светового сигнала и анализируемого ответа.
Рис. 5.
Фрагменты ЭЭГ длительностью 3 с во время фотостимуляции и световой стимул частотой 10 Гц до и после сеансов активации функциональных связей мозга (А, Б). Совместные рекуррентные диаграммы этих сигналов (В, Г). Вероятности ${{P}_{1}}(\tau )$ и ${{P}_{2}}(\tau )$ рекуррентностей для ЭЭГ (сплошная кривая) и светового сигнала (штрихпунктирная кривая) (Д, Е).
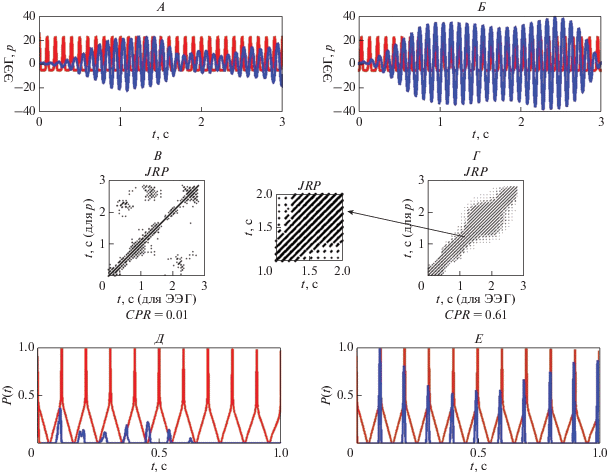
В отличие от этого, совместная рекуррентная диаграмма после сеансов (рис. 5, Г) имеет рекуррентные структуры, cодержащие длинные диагональные линии. Этой диаграмме соответствует значение показателя DET близкое к 0.9, что указывает на значительное сходство динамики сигнала возбуждения и отклика мозга. Стрелка на рис. 5 указывает на увеличенный фрагмент совместной рекуррентной диаграммы, построенной для интервала времени, равного 1 с, для того чтобы показать детально диагональные линии, равноотстоящие друг от друга на время, равное периоду колебаний светового сигнала.
Рис. 5, Д иллюстрирует отсутствие совпадения максимумов вероятностей рекуррентностей ${{P}_{1}}(\tau )$ и ${{P}_{2}}(\tau )$для фазовых траекторий светового сигнала и ответа мозга, а рис. 5, Е – наличие такого совпадения в моменты времени kτ, где k – целое число, τ = 0.1 с (соответствует периоду колебаний светового сигнала частотой 10 Гц). Совпадение положений максимумов вероятностей рекуррентностей ${{P}_{1}}(\tau )$ и ${{P}_{2}}(\tau )$ означает возникновение одновременных рекуррентностей в анализируемом фрагменте ЭЭГ и световом сигнале заданной частоты. В этом случае значение коэффициента взаимной корреляции CPR = 0.61 (рис. 5, Е). В случае отсутствия совпадения максимумов вероятностей рекуррентностей ${{P}_{1}}(\tau )$ и ${{P}_{2}}(\tau )$ для фазовых траекторий светового сигнала и ответа мозга значение коэффициента взаимной корреляции CPR = 0.01 (рис. 5, Д). Таким образом, рис. 5 представляет пример возникновения фазовой синхронизации во фрагменте ЭЭГ и световом стимуле частотой 10 Гц после примененных воздействий.
На рис. 6, А, Б представлены фрагменты ЭЭГ длительностью 6 с во время фотостимуляции и фотостимул частотой 6 Гц до и после сеансов активации функциональных связей мозга. Совместные рекуррентные диаграммы для сигнала светового возбуждения и фрагментов отклика мозга до и после этих сеансов показаны на рис. 6, В, Г. Параметры d и m такие же, как для рис. 5. Совместная рекуррентная диаграмма, полученная до сеансов (рис. 6, В), содержит длинные диагональные линии, соответствующие близкому к 0.95 значению показателя DET и свидетельствующие о значительном сходстве динамики сигнала возбуждения и ответа мозга. Положения максимумов вероятностей рекуррентностей ${{P}_{1}}(\tau )$ и ${{P}_{2}}(\tau )$ для фазовых траекторий светового сигнала и ответа мозга при этом совпадают (рис. 6, Д) в моменты времени kτ, где τ = 0.17 с, что соответствует периоду колебаний сигнала фотостимуляции частотой 6 Гц. Значение коэффициента взаимной корреляции CPR = 0.69 (рис. 6, Д). Это свидетельствует о наличии фазовой синхронизации в анализируемом фрагменте ЭЭГ и световом сигнале заданной частоты.
Рис. 6.
Фрагменты ЭЭГ длительностью 6 с во время фотостимуляции и световой стимул частотой 6 Гц до и после сеансов активации функциональных связей мозга (А, Б). Совместные рекуррентные диаграммы этих сигналов (В, Г) и вероятности ${{P}_{1}}(\tau )$ и ${{P}_{2}}(\tau )$ рекуррентностей (Д, Е). Остальные обозначения см. рис. 5.
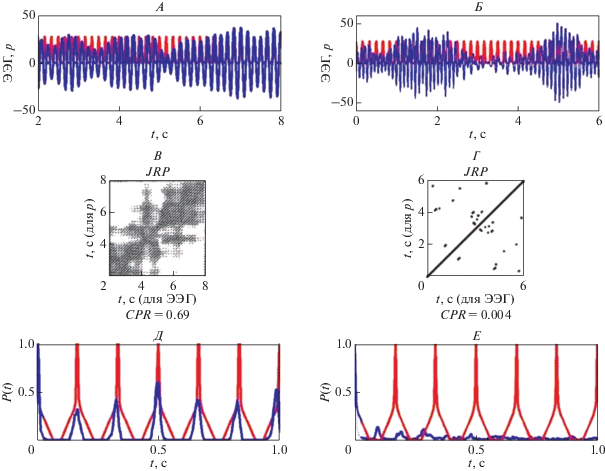
Для фрагмента ЭЭГ, полученного после сеансов активации функциональных связей мозга и светового сигнала частотой 6 Гц (рис. 6, Б), совместная рекуррентная диаграмма содержит только изолированные точки (рис. 6, Г), значение коэффициента взаимной корреляции CPR = 0.004 (рис. 6, Е). Это говорит о том, что в анализируемом фрагменте фазовая синхронизация между сигналом возбуждения и откликом отсутствует. Таким образом, рис. 6 представляет пример отсутствия фазовой синхронизации во фрагменте паттерна ЭЭГ и световом стимуле частотой 6 Гц после проведенных сеансов активации функциональных связей мозга.
Средние значения показателей реакции усвоения до и после сеансов активации функциональных связей мозга приведены в табл. 1. Усреднение проводилось по значениям, полученным для реактивных паттернов ЭЭГ 24 тестируемых лиц, т.е. 80% от общего числа пациентов, показавших наличие изменений в реактивных паттернах. В связи с отсутствием статистически значимых отличий в значениях показателей для разных затылочных локусов ЭЭГ, в табл. 1 приводятся результаты, полученные для затылочного локуса O1.
Таблица 1.
Усредненные значения коэффициентов усвоения (kR) ритма заданной частоты, показателя детерминизма (DET) в совместных рекуррентных диаграммах паттернов ЭЭГ и световых сигналов и показателя фазовой синхронизации (CPR), значения критерия Фишера (F1,47) и уровня значимости критерия (p)
| Частота, Гц | ЭЭГ пациентов до сеансов активации | ЭЭГ пациентов после сеансов | F1,47 | p |
|---|---|---|---|---|
| Коэффициент усвоения ритма заданной частоты (kR) | ||||
| 6 | 43 ± 3.7 | 7 ± 0.6 | 467 | 0.005 |
| 10 | <1 | 28 ± 2.6 | 609 | 0.0004 |
| 16 | <1 | <1 | 2.6 | 0.87 |
| Показатель детерминизма (DET) | ||||
| 6 | 0.87 ± 0.07 | 0.51 ± 0.05 | 197 | 0.01 |
| 10 | 0.53 ± 0.06 | 0.97 ± 0.09 | 208 | 0.01 |
| 16 | 0.67 ± 0.07 | 0.56 ± 0.05 | 3.9 | 0.75 |
| Коэффициент взаимной корреляции между вероятностями рекуррентностей (CPR) | ||||
| 6 | 0.49 ± 0.05 | 0.08 ± 0.009 | 401 | 0.003 |
| 10 | 0.05 ± 0.005 | 0.64 ± 0.06 | 377 | 0.002 |
| 16 | 0.06 ± 0.006 | 0.05 ± 0.006 | 2.8 | 0.91 |
При использовании дисперсионного анализа ANOVA нулевая гипотеза о равенстве средних значений показателей усвоения для двух рассмотренных групп отвергалась в случае, если статистика, полученная по F-критерию Фишера, превышала критическое значение Fcrit = F1,47 = 4.1. Значения 1 и 47 связаны с тем, что число групп k = 2, число усредняемых значений в каждой группе – 24, общее число наблюдений N = 2 × 24 = = 48, k – 1 = 1, N – k = 47. Статистически значимые отличия между группами определялись на основании значений p < 0.05 в силу того, что n = = k(k – 1)/2 = 1 и 1 – 0.951/n = 0.05.
Данные табл. 1 показывают, что усвоение частоты фотостимула 16 Гц отсутствует как до, так и после проводимых сеансов (значение коэффициента усвоения ритма kR < 1, процент рекуррентных точек, расположенных на диагональных линиях DET < 0.7, коэффициент взаимной корреляции между вероятностями рекуррентностей светового сигнала и ответа мозга CPR < 0.1, значение F-критерия Фишера F < Fcrit = 4.1 и уровня значимости этого критерия p > 0.05 для всех рассмотренных показателей).
Для частоты светового возбуждения 6 Гц до проводимых сеансов среднее значение коэффициента усвоения ритма kR = 43 ± 3.7, процент рекуррентных точек, расположенных на диагональных линиях DET = 0.87 ± 0.07, среднее значение коэффициента взаимной корреляции между вероятностями рекуррентностей светового сигнала и ответа мозга CPR = 0.49 ± 0.05, а после сеансов средние значения этих показателей уменьшаются до kR = 7 ± 0.6, DET = 0.51 ± 0.05 и CPR = 0.08 ± ± 0.009. Попарное сравнение средних значений анализируемых показателей вейвлетных спектров и совместных рекуррентных диаграмм до и после сеансов показывает, что значения статистики Фишера F > Fcrit = 4.1 и соответствуют малым значениям уровня значимости p (p < 0.02). Это свидетельствует о наличии достоверных различий между средними значениями рассмотренных показателей для частоты фотостимула 6 Гц до и после проводимых сеансов.
Для частоты светового возбуждения 10 Гц, наоборот, после проводимых сеансов, средние значения коэффициента усвоения ритма, процента рекуррентных точек, расположенных на диагональных линиях и коэффициента взаимной корреляции между вероятностями рекуррентностей светового сигнала и ответа мозга увеличиваются до величин kR = 28 ± 2.6, DET = 0.97 ± 0.09 и CPR = 0.64 ± 0.06, по сравнению со средними значениями до этих сеансов kR < 1, DET = 0.53 ± ± 0.06 и CPR = 0.05 ± 0.005. Значения критерия Фишера F > Fcrit = 4.1 и уровня значимости p < 0.02 демонстрируют наличие достоверных различий между средними значениями рассмотренных показателей для частоты фотостимуляции 10 Гц до и после проводимых сеансов.
Таким образом, оба метода, используемые в работе, указывают на то, что для пациентов с умеренными когнитивными нарушениями после сеансов, направленных на формирование функциональных связей мозга человека, отмечается повышение показателей реакции усвоения ритма частоты возбуждения α-диапазона и уменьшается значение этих показателей для частоты возбуждения θ-диапазона.
Уменьшение значений рассмотренных показателей свидетельствует о снижении или даже исчезновении сходства динамического поведения стимула и ответа мозга на него. При этом фазовая синхронизация между откликом мозга и фотостимулом до сеансов наблюдается только для частоты возбуждения θ-диапазона, а после этих сеансов – для частоты возбуждения α-диапазона.
Усиление реакции усвоения в α-диапазоне и ослабление в θ-диапазоне указывает на улучшение функционального состояния коры больших полушарий и улучшение ее регуляторных механизмов. Следует отметить, что наши данные корреллируют с результатами психофизиологического тестирования, проведенного для этих же групп пациентов в работе [16], где для них было показано почти трехкратное повышение индекса кратковременной памяти после примененного курса воздействий.
Известно, что динамика перестроек электрической активности мозга в соответствии с частотой световых импульсов связана с возбудимостью нейронов коры головного мозга [19]. Как показано в наших предыдущих работах, изменение показателей реакции усвоения ритмов фотостимуляции может быть использовано для оценки изменения функционального состояния пациентов с сосудистой патологией [27, 28], с нарушениями сердечного ритма [35], а также с невротическими расстройствами [24, 25]. При этом повышение показателей реакции усвоения ритма θ- и (или) β-диапазонов связаны с изменением уровня психоэмоциональной возбудимости пациента [27, 28]. Увеличение показателей реакции усвоения частот θ-диапазона у больных с нарушениями сердечного ритма в форме фибрилляции предсердий по мере нарастания проявлений этих нарушений, т.е. при переходе от пароксизмальной формы аритмии в постоянную, связано с преобладанием процессов торможения в нервной системе этих пациентов [35]. Снижение степени невротизации пациентов с невротическими расстройствами в виде панических атак после сеансов, направленных на формирование функциональных связей мозга, отражается в уменьшении количественных показателей реакции усвоения ритмов фотостимуляции β-диапазона и связано с понижением возбудимости нейронов коры больших полушарий [24, 25]. В случае умеренных когнитивных нарушений повышение количественных показателей реакции усвоения ритма частоты α-диапазона, вероятно, связано с повышением возбудимости и лабильности корковых нейронов после проведенных сеансов.
ЗАКЛЮЧЕНИЕ
Проанализированы изменения реакции усвоения ритмов фотостимуляции и определены количественные характеристики этой реакции, вычисленные до и после сеансов терапии, связанных c воздействием, направленным на активацию функциональных связей мозга у лиц с умеренными когнитивными нарушениями.
Полученные результаты являются еще одним доказательством высокой чувствительности примененных методов анализа вейвлетных спектров и показателей совместных рекуррентных диаграмм к выявлению динамических изменений, происходящих в реактивных паттернах биоэлектрической активности мозга в ответ на стимулирующие воздействия. Эти результаты могут быть использованы в клинике для достоверного нахождения улучшения функционального состояния головного мозга пациентов с умеренными когнитивными нарушениями.
Этические нормы. Все исследования проведены в соответствии с принципами биомедицинской этики, сформулированными в Хельсинкской декларации 1964 г. и ее последующих обновлениях, и одобрены локальным биоэтическим комитетом Института мозга человека им. Н.П. Бехтеревой (протокол КПЭ ИМЧ РАН от 18 апреля 2013 г.) (Санкт-Петербург).
Информированное согласие. Каждый участник исследования представил добровольное письменное информированное согласие, подписанное им после разъяснения ему потенциальных рисков и преимуществ, а также характера предстоящего исследования.
Благодарности. Авторы выражают благодарность Татьяне Николаевне Резниковой за предоставленные экспериментальные записи ЭЭГ.
Финансирование работы. Работа выполнена при финансовой поддержке ПФНИ государственных академий на 2013–2020 гг. (ГП-14, раздел 64).
Конфликт интересов. Авторы декларируют отсутствие явных и потенциальных конфликтов интересов, связанных с публикацией данной статьи.
Список литературы
Дробижев М.Ю., Макух Е.А., Дзантиева А.И. Когнитивные расстройства в общей медицине // Лечащий врач. 2007. № 8. С. 35. Drobizhev M.Yu., Makukh E.A., Dzantieva A.I. [Cognitive disorders in general medicine] // Lechashiy Vrach. 2007. № 8. P. 35.
Парфенов В.А. Лечение постинсультных когнитивных нарушений // Русский медицинский журнал. 2010. № 16. С. 986. Parfenov V.A. [Treatment of post-stroke cognitive impairment] // Medical J. Russian Federation. 2010. № 16. P. 986.
Новикова И.А., Соловьев А.Г., Местечко В.В. Особенности эмоциональной сферы лиц пожилого возраста с пограничными психическими расстройствами // Медицинская психология в России: электрон. науч. журн. 2014. № 4. С. 27. Novikova I.A., Soloviev A.G., Mestechko V.V. [Features of the emotional sphere of elderly people with borderline mental disorders] // Medicinskaâ Psihologiâ v Rossii. 2014. № 4. P. 27.
Антипенко Е.А. Антиооксидантная терапия при дисциркуляторной энцефалопатии // Ж. неврологии и психиатрии им. С.С. Корсакова. 2010. № 110. С. 53. Antipenko E.A. [Antioxidant therapy for discirculatory encephalopathy] // Zh. Nevrol. Psikhiatr. Im. S.S. Korsakova. 2010. № 110. P. 53.
Шарашкина Н.В., Остапенко В.С., Ткачева О.Н. и др. Когнитивные нарушения у пациентов с артериальной гипертонией пожилого и старческого возраста // Кардиоваскулярная терапия и профилактика. 2017. № 16. С. 34. Sharashkina N.V., Ostapenko V.S., Tkacheva О.N. et al. [Cognition disorders in hypertensive patients of older age and senile] // Kardiovaskulyarnaya Terapiya i Profilaktika. 2017. № 16. P. 34.
Qureshi S.U., Long M.E., Bradshaw M.R. et al. Does PTSD impair cognition beyond the effect of trauma? // J. Neuropsychiatry Clin. Neurosci. 2011. V. 23. № 1. P. 16.
Bierman E.J.M., Comijs H.C., Rijmen F. et al. Anxiety symptoms and cognitive performancein later life: results from the longitudinal aging study // Aging Ment. Health. 2008. V. 12. № 4. P. 517.
Gallacher R. Early stage dementia // Nurs. Standard. 2014. V. 28. № 20. P. 61.
Табеева Г.Р. Когнитивные и некогнитивные расстройства у пациентов пожилого возраста, ассоциированные со стрессом // Неврология, нейропсихиатрия, психосоматика. 2015. № 1. С. 87. Tabeeva G.R. [Cognitive and non-cognitive disorders in elderly patients associated with stress] // Nevrologiya, Neyropsikhiatriya, Psikhosomatika. 2015. № 1. P. 87.
Смирнов В.М., Бородкин Ю.С., Резникова Т.Н. Свойство мозга человека в условиях активации подкорковых структур формировать комплекс устойчивых внутримозговых функциональных связей / Научные открытия (Сборник кратких описаний). СПб., 1999. Вып. 1. С. 23. Smirnov V.M., Borodkin Yu.S., Reznikova T.N. [The property of the human brain in conditions of activation of subcortical structures to form a complex of stable intracerebral functional connections]. In: Nauchnyye otkrytiya (Sbornik kratkikh opisaniy). SPb., 1999. Issue. 1. P. 23.
Friston K.J. Functional and effective connectivity in neuroimaging: A synthesis // Hum. Brain Mapping. 1994. V. 2. P. 56.
Achard S., Salvador R., Whitcher B. et al. A resilient, low-frequency, small-world human brain functional network with highly connected association cortical hubs // J. Neurosci. 2006. V. 26. № 1. P. 63.
Tononi G., Sporns O. Edelman G.M. A measure for brain complexity: relating functional segregation and integration in the nervous system // Proc. Natl. Acad. Sci. U.S.A. 1994. V. 91. № 11. P. 5033.
Резникова Т.Н., Краснов А.А., Селиверстова Н.А. Исследование “внутренней картины болезни” у больных с органической и функциональной патологией ЦНС в процессе лечебных активаций методом артифициальных стабильных функциональных связей мозга человека // Вестник клинической психологии. 2004. Т. 2. № 1. С. 84. Reznikova T.N., Krasnov A.A., Seliverstova N.A., Terent’eva I.Yu. [The study of the “internal picture of the disease” in patients with organic and functional pathology of the central nervous system during therapeutic activations by the artificially stable functional connections of the human brain] // Vestn. Klin. Psikhol. 2004. V. 2. № 1. P. 84.
Резникова Т.Н., Федоряка Д.А., Селиверстова Н.А., Моховикова И.А. Опыт использования сенсорной импульсной стимуляции у больных с паническими атаками // Вестник психотерапии. 2018. № 68. С. 47. Reznikova T.N., Fedoryaka D.A., Seliverstova N.A., Mokhovikova I.A. [Experience of using sensory impulse stimulation in patients with panic attacks] // Vestnik Psikhoterapii. 2018. № 68. P. 47.
Резникова Т.Н., Селиверстова Н.А. О коррекции психического состояния немедикаментозным способом при рассеянном склерозе // Клиническая и специальная психология. 2019. Т. 8. № 1. С. 177. Reznikova T.N. Seliverstova N.A. [On the correction of the mental state in a non-drug way in multiple sclerosis] // Clinical Psychology and Special Education. 2019. V. 8. № 1. P. 177.
Резникова Т.Н., Селиверстова Н.А., Дик О.Е. и др. Оценка психофизиологического состояния у пожилых лиц с умеренными когнитивными нарушениями при сенсорных импульсных стимуляциях // Психическое здоровье. 2020. № 9. С. 12. Reznikova T.N., Seliverstova N.A., Dick O.E. et al. [Assessment of the psychophysiological state in the elderly with moderate cognitive impairment during sensory impulse stimulation] // Psikhicheskoye Zdorov’ye. 2020. № 9. P. 12.
Святогор И.А., Резникова Т.Н., Петухова Н.В. Динамика психофизиологического состояния в процессе лечения методом артифициальных стабильных функциональных связей // Биологическая обратная связь. 2001. № 4. С. 24. Svyatogor I.A., Reznikova T.N., Petukhova N.V. [Dynamics of the psychophysiological state in the process of treatment by the method of artifactual stable functional connections] // Biologicheskaya Obratnaya Svyaz. 2001. № 4. P. 24.
Федотчев А.И., Бондарь А.Т., Акоев И.Г Динамические характеристики резонансных ЭЭГ реакций на ритмическую фотостимуляцию // Физиология человека. 2000. Т. 26. № 2. С. 64. Fedotchev I., Bondar A.T., Akoev I.G. Dynamic characteristics of the human resonance EEG responses to rhythmic photostimulation // Human Physiology. 2000. V. 26. № 2. P. 179.
Notbohm A., Kurths J., Herrmann C.S. Modification of brain oscillations via rhythmic light stimulation provides evidence for entrainment but not for superposition of event-related responses // Front. Hum. Neurosci. 2016. V. 10. P. 10.
Dik O.E., Glazov A.L. Parameters of phase synchronization in electroencephalographic patterns as markers of cognitive impairment // Technical Physics. 2021. V. 66. № 4. P. 661. https://doi.org/10.1134/S1063784221040058
Hramov A.E., Koronovsky A.A., Makarov V.A. et al. Wavelets in neuroscience. Springer Series in Synergetcs. Berlin: Springer, 2015. 314 p.
Marwan N., Romano M.C., Thiel M. et al. Recurrence plots for the analysis of complex systems // Phys. Rep. 2007. V. 438. № 5. P. 237.
Дик О.Е., Святогор И.А., Резникова Т.Н. и др. Анализ паттернов ЭЭГ у лиц с паническими атаками // Физиология человека. 2020. Т. 46. № 2. С. 63. Dick O.E., Svyatogor I.A., Reznikova T.N. et al. Analysis of EEG Patterns in Subjects with Panic Attacks // Human Physiology. 2020. V. 46. № 2. P. 163.
Dick O.E. Wavelet and recurrence analysis of EEG patterns of subjects with panic attacks // Studies in Computational Intelligence. 2020. V. 856. P. 172.
Torrence C., Compo G.P. A practical guide to wavelet analysis // Bull. Am. Meteorol. Soc. 1998. V. 79. № 1. P. 61.
Dick O.E., Svyatogor I.A. Wavelet and multifractal estimation of the intermittent photic stimulation response in the electroencephalogram of patients with dyscirculatory encephalopathy // Neurocomputing. 2015. V. 165. № 1. P. 361.
Святогор И.А., Дик О.Е., Ноздрачев А.Д., Гусева Н.Л. Анализ изменений ЭЭГ-паттернов в ответ на ритмическую фотостимуляцию при различных нарушениях функционального состояния ЦНС // Физиология человека. 2015. Т. 41. № 3. С. 41. Svyatogor I.A., Dick O.E., Nozdrachev A.D., Guseva N.L. Analysis of changes in EEG patterns in response to rhythmic photic stimulation under various disruptions of the functional state of the central nervous system // Human Physiology. 2015. V. 41. № 3. P. 261.
Takens F. Detecting strange attractors in turbulence / Dynamical Systems and Turbulence, Lecture Notes in Mathematics // Eds. Rand D., Young L.S. Berlin: Springer-Verlag. 1981. V. 898. P. 366.
Fraser A.M., Swinney H.L. Independent coordinates for strange attractors from mutual information // Phys. Rev. A. 1986. V. 33. № 2. P. 1134.
Kennel M.B., Brown R., Abarbanel H.D. Determining embedding dimension for phase-space reconstruction using a geometrical construction // Phys. Rev. A. 1992. V. 45. № 6. P. 3403.
Romano M.C., Thiel M., Kurths J. et al. Detection of synchronization for non-phase-coherent and non-stationary data // Europhys. Lett. 2005. V. 71. № 3. P. 466.
Kurths J., Romano M.C., Thiel M. et al. Synchronization analysis of coupled noncoherent oscillators // Nonlinear Dynam. 2006. V. 44. № 1. P. 135.
Русинов В.С., Гриндель О.М., Болдырева Г.Н. и др. Биопотенциалы мозга человека. Математический анализ. М.: Медицина, 1987. 254 с.
Дик О.Е., Святогор И.А., Ноздрачев А.Д., Гусева Н.Л. Анализ реактивных паттернов ЭЭГ у лиц с фибрилляцией предсердий // Физиология человека. 2019. Т. 45. № 1. С. 49. Dick O.E., Svyatogor I.A., Nozdrachev A.D., Guseva N.L. Analysis of the reactive patterns of EEGs under atrial fibrillation // Human Physiology. 2019. V. 45. № 1. P. 40.
Дополнительные материалы отсутствуют.
Инструменты
Физиология человека