

Физиология человека, 2022, T. 48, № 3, стр. 14-23
Окуломоторная активность у детей 4–6 лет при рассматривании изображения
Е. А. Буденкова 1, *, Д. А. Швайко 1
1 ФГАОУ ВО Балтийский федеральный университет имени И. Канта
Калининград, Россия
* E-mail: KBudenkova@gmail.com
Поступила в редакцию 17.03.2021
После доработки 02.12.2021
Принята к публикации 09.02.2022
- EDN: XRXQKU
- DOI: 10.31857/S0131164622030043
Аннотация
Движения глаз взаимосвязаны с когнитивным процессом, в связи с чем анализ окуломоторной активности представляется полезным инструментом в области изучения зрительного внимания, рабочей памяти и анализа зрительной информации в общем. В данном исследовании получена характеристика саккадического и фиксационного компонентов движений глаз у детей дошкольного возраста (от 4 до 6 лет) с разным восприятием статического изображения. Выделены четыре группы детей на основе качества ответа на вопрос о том, что изображено на картинке (правильный ответ, неправильный ответ, нет ответа, правильный ответ после подсказки). Установлены окуломоторные особенности для этих четырех групп. Полученные результаты вносят вклад в понимание разных типов когнитивных стратегий, что представляется полезным для построения индивидуальных траекторий обучения в школе, формирования оптимальной когнитивной стратегии анализа информации, необходимой для успешной жизнедеятельности.
Исследованию окуломоторной активности у детей школьного возраста посвящено значительное количество работ в рамках специальных стимульных парадигм (Step, Gap, Overlap и др.) и при чтении текстов разной сложности [1, 2]. Работ же, посвященных изучению окуломоторной активности при свободном рассматривании крайне мало. Однако представляется полезным использование изображений в качестве стимульного материала для изучения нормальной динамики окуломоторных параметров в онтогенезе в силу большей экологической валидности.
В области исследования движений глаз при рассматривании изображений к настоящему времени накоплен ряд сведений: окуломоторный паттерн зависит от свойств зрительного стимула, типа задания, физиологического состояния и индивидуальных особенностей обследуемого [3–6]. Фиксации при рассматривании изображения распределяются по зонам интереса, на одних элементах изображения взгляд задерживается дольше, на других меньше, часть элементов не рассматривается вовсе. Значительное внимание уделялось изучению взаимосвязи между движениями глаз и когнитивными способностями [7–9]. Согласно немногочисленным исследованиям известно, что окуломоторный паттерн при просмотре изображения преобразуется по мере взросления: амплитуда саккад достигает значений, наблюдаемых у взрослых (2–4 угл. град) к 4–6 годам, продолжительность фиксаций сокращается (200–300 мс) к 6–8 годам [9].
Система регуляции окуломоторной активности задействует структуры разных отделов головного мозга, которые, хоть и менее интенсивно, продолжают развитие постнатально [10]. Структуры, вовлеченные в контроль саккад, связаны с обеспечением когнитивных функций [10, 11]. В связи с чем, саккады представляют интерес в качестве косвенного нейрофизиологического маркера сформированности внимания, памяти, восприятия и более сложных интегративных процессов. Активность корковых глазодвигательных полей связана как с выполнением саккад, так и с обеспечением ряда когнитивных процессов (внимание, рабочая память, категоризация, селекция мишени, зрительный поиск, мониторинг ошибок, принятие решения, вознаграждение и прогнозирование).
К 3–4 годам жизни масса головного мозга увеличивается втрое и продолжает расти до 6–7 лет, после чего объем возрастает медленно, достигая максимального значения к 20–29 годам. В 2–3 года большинство ядер ствола мозга хорошо развито, однако ряд структур среднего (красное ядро, черное вещество) и промежуточного мозга созревают в постнатальный период примерно к 13–16 годам [12]. К 7 годам масса мозжечка достигает массы мозжечка взрослого человека. Интенсивное развитие структур мозжечка происходит в период полового созревания. В течение первых трех лет происходит интенсивный рост и миелинизация отростков нейронов, дифференцирование нейронов в слоях коры. В период от 3 до 10 лет увеличивается количество ассоциативных волокон, завершается процесс развития корковых формаций, но тонкая дифференцировка продолжается до 16–18 лет [12, 13]. Обобщая вышеизложенную информацию можно сказать, что постнатальное развитие головного мозга в целом и структур, опосредующих как движения глаз, так и когнитивные функции, протекает гетерохронно. Такое нелинейное “созревание” головного мозга, в свою очередь, может отражаться в нелинейном изменении окуломоторных параметров [14].
В настоящем исследовании мы изучаем произвольную глазодвигательную активность при просмотре изображения детьми от 4 до 6 лет. У детей без диагностированных заболеваний, на момент участия в исследовании, каких-либо патологий не выявлено, и развитие характеризуется как нормальное. В работе преследуются две основные цели. Во-первых, изучить возрастную динамику параметров окуломоторной активности в группах 4, 5, 6 лет. Во-вторых, исследовать взаимосвязь между произвольной окуломоторной активностью у детей при просмотре изображения и степенью понимания увиденного. Для достижения поставленных целей зарегистрирована окуломоторная активность у детей от 4 до 6 лет, проанализированы полученные данные погодово и с учетом когнитивного разделения.
МЕТОДИКА
В исследовании принимали участие 160 воспитанников детских садов № 132 и 45 (г. Калининград). Участники исследования не имели отклонений в развитии. Часть участников была не в состоянии выполнить калибровку с необходимым уровнем точности, поэтому к статистическому анализу допущены данные 135 детей (табл. 1).
Таблица 1.
Половозрастной состав исследуемой группы
| Группы, возраст | 4 года 7 мес. ± 3 мес. | 5 лет 6 мес. ± 6 мес. | 6 лет 3 мес. ± 1 мес. |
|---|---|---|---|
| Количество (м; ж) | 34 (15; 19) | 78 (46; 32) | 23 (10; 13) |
Регистрацию окуломоторной активности проводили в помещении детского сада, соответствующем техническим и санитарным нормам, с 9 до 12 ч утра. Участники исследования располагались за столом на расстоянии 60 ± 5 см от монитора ноутбука, по центру. Положение головы во время эксперимента не фиксировали. Участники получали инструкцию сидеть ровно, стараясь не двигаться, внимательно отслеживать перемещения стимульного объекта по экрану только взглядом.
Участникам было предложено к произвольному рассматриванию цветное фото с изображением птиц, сидящих в ряд на ветке, развернутое на весь экран ноутбука. Изображение было подобрано с учетом вероятности неоднозначности восприятия (ряд птиц можно принять за гусеницу). Разрешение изображения 1024 × 768 пикселей, масштаб и качество оригинального фото при расширении на весь экран не пострадали (рис. 1), экспозиция составляла 10 с. Перед просмотром целевого изображения предъявляли ряд адаптационных изображений, не учитываемых при дальнейшем анализе. Чтобы оценить фоновое состояние зрительного внимания и рабочей памяти, участников исследования не предупреждали заранее о предстоящей проверке: после рассматривания изображения задавали вопрос о том, что участники увидели (“Что было на изображении?”). В случае неверного ответа или при возникновении затруднений участнику исследования задавали дополнительные наводящие вопросы, например, “оно живое или неживое?”, “видел(а) ли ты это на улице?”.

Для оценки когнитивных функций участников исследования разделяли на семантические группы по типу ответа на вопрос после просмотра изображения: R – правильно поняли и описали изображение (за правильный ответ принимали – “птицы”, “птицы на ветке”), W – неправильно описали изображение (самый популярный неверный ответ – “гусеница”), NoA – не смогли описать изображение даже после наводящих вопросов, RwP – правильно описали изображение только после наводящих вопросов. Группа R включала детей 4–6 лет, группа W – 5 лет, NoA – 5–6 лет, RwP – 4–5 лет. Соотношение по полу равномерно в трех группах из четырех, а группа W на три четверти состояла из мальчиков.
Движения глаз регистрировали с помощью аппарата RED250 mobile “SensoMotoric Instruments” (Германия). Частота регистрации 120 Гц, пространственное разрешение 0.1° с погрешностью 0.5°. При регистрации аппарат определяет расположение головы испытуемого перед монитором и относительно положения головы определяет движения зрачков. Мы решили не фиксировать положение головы участников исследования при выполнении ими задания по ряду причин: приблизить условия к естественным (в работе R. Krauzlis et al. [15] упоминается предположение о возможном видоизменении движений глаз при фиксации головы), исключить негативную эмоциональную реакцию и отвлечение внимания на наше приспособление для фиксации. Перед началом выполнения задания для каждого участника проводили калибровку айтрекера по пяти точкам (точность (X/Y) не менее 0.2°/0.2°). Сырые данные обработаны при помощи программы BeGaze 3.0 “SensoMotoric Instruments” (Германия). Для подачи стимульного материала использовали ноутбук Acer Aspire E5-551G-F63G с матовым экраном (диагональ 15.6'', разрешение 1920 × 1080, 60 Гц). На расстоянии 60 ± 5 см от монитора охват экрана составляет 29° ± 1° по горизонтали и 14° ± 2° по вертикали, изображение ограничено – составляет 25° ± 1° по горизонтали и 9° ± 1° по вертикали. Для оценки глазодвигательной активности при просмотре изображения определяли параметры: количество и среднюю продолжительность фиксаций (в анализ не включали крайние значения, выходящие за пределы М ± 3σ), амплитуду, среднюю и пиковую скорость саккад, диаметр зрачка при фиксациях характер распределения фиксаций. Анализировали данные, зарегистрированные по правому глазу.
Статистическую обработку проводили с помощью SPSS 21.0 и Exсel, оценку распределений на нормальность – с использованием критерия Колмогорова-Смирнова (приводится стандартизированное значение критерия, z). Принимали гипотезу о нормальности распределения при p ≤ 0.05. В качестве меры центральной тенденции использовали медиану (М), как менее чувствительный к выбросам показатель. Для оценки отклонения значений от медианы рассчитывали среднеквадратическое отклонение (σ). С помощью однофакторного дисперсионного анализа (ANOVA) проанализировали влияние фактора “возраст” (три уровня: 4, 5 и 6 лет) и фактора “ответ на вопрос после просмотра изображения” (четыре уровня: правильный ответ (R), правильный ответ после подсказки (RwP), неправильный ответ (W) и без ответа (NoA)) на окуломоторные переменные. Уровень значимости равен, с учетом поправки Бонферрони, p ≤ 0.017 и p ≤ 0.008 для первого и второго фактора, соответственно. Для проверки наличия корреляции между возрастом и количественными показателями саккад использовали коэффициент корреляции Пирсона (достоверность различий при p ≤ 0.05). Для межгруппового сравнения независимых выборок использовали t-критерий с учетом поправки Бонферрони, где p ≤ 0.017 для сравнения групп по возрасту (4, 5 и 6 лет) и p ≤ 0.008 – для семантических групп (R, W, NoA, RwP).
РЕЗУЛЬТАТЫ ИССЛЕДОВАНИЯ И ИХ ОБСУЖДЕНИЕ
В настоящей работе авторы рассматривают окуломоторную активность детей дошкольного возраста, разделяя участников на возрастные группы (4 года, 5 и 6 лет) и на семантические группы (правильный ответ (R), неправильный ответ (W), без ответа (NoA), правильный ответ после подсказки (RwP)). В состав каждой семантической группы входят участники 4, 5 и 6 лет. Полученные данные в общем виде представлены в табл. 2.
Таблица 2.
Показатели окуломоторной активности разных групп
| Группы | Возрастные группы | Семантические группы | |||||
|---|---|---|---|---|---|---|---|
| 4 года | 5 лет | 6 лет | R | W | NoA | RwP | |
| Саккады | |||||||
| Амплитуда, град | 1.8 ± 0.7 | 1.6 ± 0.7 | 2.0 ± 1.4 | 1.5 ± 0.4 | 1.5 ± 0.3 | 1.2 ± 0.1 | 1.8 ± 0.7 |
| Скорость ср.1 | 69 ± 13 | 65 ± 17 | 68 ± 19 | 62 ± 13 | 65 ± 4 | 62 ± 6 | 70 ± 19 |
| Скорость пик.1 | 186 ± 37 | 168 ± 53 | 175 ± 55 | 164 ± 38 | 148 ± 15 | 176 ± 23 | 179 ± 46 |
| Фиксации | |||||||
| Количество, ед. | 21 ± 6 | 23 ± 6 | 20 ± 6 | 23 ± 6 | 19 ± 2 | 28 ± 3 | 22 ± 6 |
| Длительность, мс | 334 ± 79 | 316 ± 87 | 387 ± 114 | 353 ± 91 | 243 ± 32 | 339 ± 65 | 308 ± 61 |
| Диаметр зрачка, мм | 4.3 ± 0.7 | 4.4 ± 0.6 | 4.3 ± 0.5 | 4.3 ± 0.5 | 4.0 ± 0.5 | 4.4 ± 0.6 | 4.3 ± 0.6 |
Проверка данных на нормальность распределения (табл. 3) показала, что большинство окуломоторных параметров подчиняются нормальному распределению. Исключение составляет распределение амплитуды саккад в группах детей 5 и 6 лет.
Таблица 3.
Результаты проверки на нормальность распределения
| Группы | Возрастные группы | Семантические группы | |||||
|---|---|---|---|---|---|---|---|
| 4 года | 5 лет | 6 лет | R | W | NoA | RwP | |
| Саккады | |||||||
| Амплитуда, град | 0.4; 0.9 | 1.4; 0.02* | 1.4; 0.02* | 0.7; 0.6 | 0.4; 0.9 | 0.8; 0.4 | 0.6; 0.8 |
| Скорость ср. | 0.5; 0.8 | 1.0; 0.2 | 0.8; 0.4 | 0.7; 0.6 | 0.5; 0.9 | 0.7; 0.6 | 0.6; 0.7 |
| Скорость пик. | 0.6; 0.7 | 0.7; 0.6 | 0.7; 0.5 | 0.6; 0.7 | 0.4; 0.9 | 0.4; 0.9 | 0.6; 0.7 |
| Фиксации | |||||||
| Количество, ед. | 0.5; 0.8 | 0.9; 0.3 | 0.3; 0.9 | 0.6; 0.7 | 0.9; 0.3 | 0.5; 0.9 | 0.8; 0.4 |
| Длительность, мс | 0.4; 0.9 | 1.0; 0.2 | 0.7; 0.7 | 0.9; 0.3 | 0.4 ;0.9 | 0.5; 0.9 | 0.4; 0.9 |
| Диаметр зрачка, мм | 0.6; 0.8 | 0.9; 0.3 | 0.5; 0.9 | 0.6; 0.7 | 0.9; 0.3 | 0.6; 0.7 | 0.7; 0.5 |
Примечание: приведены значения критерия Колмогорова–Смирнова (z; p); * – распределение не нормально, при р ≥ 0.05. Остальные обозначения см. табл. 2.
В результате дисперсионного анализа обнаружено, что влияние фактора “ответ на вопрос после просмотра изображения” статистически значимо лишь в отношении параметра длительность фиксаций (F(3, 128) = 3.76, p ≤ 0.001). По остальным параметрам статистически значимых различий не обнаружено: количество фиксаций (F(3, 128) = = 2.36, p ≥ 0.08), диаметр зрачка при фиксациях (F(3, 128) = 0.44, p ≥ 0.72), амплитуда саккад (F(3, 128) = 2.61, p ≥ 0.06), средняя (F(3, 128) = = 1.31, p ≥ 0.27) и пиковая (F(3, 128) = 1.15, p ≥ 0.33) скорость саккад. Фактор “возраст” оказался статистически не значимым в отношении окуломоторных параметров: длительность (F(2, 129) = 2.56, p ≥ 0.08) и количество фиксаций (F(2, 129) = 2.71, p ≥ 0.07), диаметр зрачка при фиксациях (F(2, 129) = = 0.42, p ≥ 0.65), амплитуда саккад (F(2, 129) = = 2.25, p ≥ 0.11), средняя (F(2, 129) = 1.54, p ≥ 0.22) и пиковая (F(2, 129) = 1.28, p ≥ 0.28) скорость саккад.
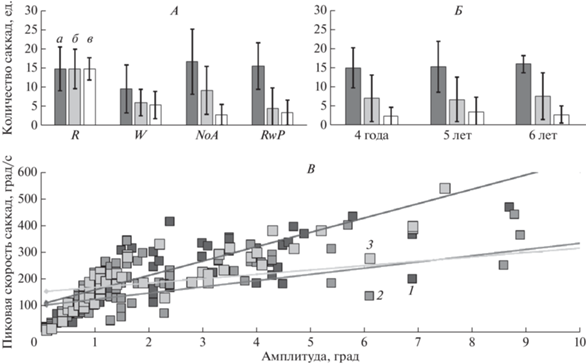
Стоит отметить, что средние значения амплитуды саккад, не превышают размера 2°. Релевантными принято считать те саккады, амплитуда которых превышает размер фовеа, т.е. 2°. Так, в работе А. Helo et al. средние значения амплитуд саккад всех возрастов превышают 3 град [10]. При этом в исследовании A. Helo et al. участники также располагались на расстоянии 60 см от экрана, в качестве стимульного материала использован ряд цветных фото (впрочем, не указано каких именно), использовался более мощный айтрекер. С другой стороны, диаграмма распределения соотношений между пиковой скоростью и амплитудой саккад в данном исследовании (рис. 2) имеет схожий вид с таковым у взрослых людей в норме [6]. Это, в совокупности с заявленными функциональными характеристиками REDm, позволяет допустить подлинность значений амплитуды зарегистрированных саккад.
Рис. 2.
Характеристика саккад при рассматривании изображения. А – количество саккад в семантических группах, Б – количество саккад в возрастных группах, В – соотношение пиковой скорости и амплитуды саккад; а – амплитуда саккад ≤1 град, б – амплитуда саккад 2−10 град, в – амплитуда саккад >10 град. R – правильный ответ, W – неправильный ответ, NoA – без ответа, RwP – правильный ответ после подсказки. 1 – 4 года, 2 – 5 лет, 3 – 6 лет.
Среднее значение диаметра зрачка при рассматривании изображения в среднем не отличается между группами, однако корреляционный анализ показал, что по мере рассматривания изображения диаметр зрачка увеличивается в группах R, W и RwP (r = 0.406; p ≤ 0.001; r = 0.740 p ≤ 0.01; r = 0.720; p ≤ 0.001 соответственно). В группе NoA корреляции между временем выполнения задания и диаметром зрачка не обнаружено (r = –0.024; p ≥ 0.500).
Для оценки динамики продолжительности зрительных фиксации по мере рассматривания изображения было проведено сравнение средней продолжительности фиксаций в начале просмотра (первые пять фиксаций) и по завершению (последние пять фиксаций) просмотра (рис. 3).
Рис. 3.
Средняя продолжительность фиксаций в начале и конце просмотра изображения. А – возрастные группы, Б – семантические группы. а – начало просмотра, б – конец просмотра изображения. * – статистически значимые различия. Остальные обозначения см. рис. 2.

Относительно групп 4, 5 и 6 лет никаких различий по длительности фиксаций между началом и концом просмотра изображения не обнаружено. В пределах семантических групп обнаружено, что продолжительность фиксаций по завершению просмотра снижается в группе R (t (128) = 24.14; p ≤ 0.001) и увеличивается в группе RwP (t (128) = = –7.03; p ≤ 0.002).
При рассматривании зрительные фиксации участников исследования располагались одиночно или группами (рис. 4).
Рис. 4.
Распределение фиксаций при рассматривании изображения. А – правильный ответ (R), Б – неправильный ответ (W), В – без ответа (NoA), Г – правильный ответ после подсказки (RwP). Первые изображения в рядах – усредненная тепловая карта группы, вторые и третьи изображения в рядах – частные примеры распределения фиксаций без учета их продолжительности (окружности – места фиксаций).

Фиксации объединяются в группы при перекрытии окружностей, которыми отображаются фиксации (рис. 4). В пределах каждой группы можно условно выделить доминантную зону фиксирования взора (на которую приходится максимальное время фиксаций) или доминанту в сочетании с субдоминантой(ми). Во всех группах доминанта расположена в верхне-центральной зоне изображения, во всех группах кроме NoA субдоминанты развиты слабо. Стоит отметить наличие в группе W горизонтальной ориентированности окуломоторной активности. Для удобства характеристики распределения зрительных фиксаций можно использовать условные формулы, описанные в табл. 4.
Таблица 4.
Характеристика распределения фиксаций при рассматривании изображения
| Группы участников | R | W | NoA | RwP |
|---|---|---|---|---|
| Общее число групп | 2–3 | 4–6 | 5–7 | 3–5 |
| Малая группа (2–5 фиксаций), А | 1–2 | 3–6 | 4–7 | 1–3 |
| Средняя группа (6–9 фиксаций), В | 0 | 0–2 | 0–1 | 0–2 |
| Крупная группа (от 10 фиксаций), С | 1 | 0–1 | 0–1 | 0–3 |
| Количество одиночных фиксаций | 1–8 | 2–5 | 3–7 | 3–6 |
| Общая формула фиксаций | А + С | А; А + В; А + С | А; А + В; А + С | А; А + В; А + С |
| Частные примеры формул | 2А + С | 6А; 3А + 2В; 3А + С | 7А; 4А + В; 4А + С | 3А; 3А + 2В; 3А + 2С |
Примечание: обозначения см. табл. 2.
Группы фиксаций можно разделить на малые (2–5 фиксации, А), средние (6–9 фиксаций, В) и крупные (от 10 и более фиксаций, С). Обнаружено, что для группы R, в отличие от остальных, характерно наличие одной крупной группы фиксаций и пары малых.
Сравнение полученных значений скорости саккад и длительности фиксаций с литературными данными затруднительно, в силу различных условий проведения исследований, т.к. это было бы некорректно. К тому же исследований, подходящих для сравнения полученных результатов, крайне мало.
При произвольном рассматривании статических изображений взрослыми амплитуда саккад уменьшается, а продолжительность фиксаций растет со временем [16]. Такие изменения могут быть отражением смены режима “общего” восприятия на “детальный” [17]. В данной работе такая зависимость не обнаружена. У детей, успешно ответивших на вопрос, напротив, средняя длительность фиксаций снижается, а значение амплитуды саккад остается неизменным. В группе RwP амплитуда саккад также не различается между началом и окончанием просмотра, но продолжительность фиксации заметно увеличивается. В группе W амплитуда саккад увеличивается, а в NoA – уменьшается. Неоднородность результатов позволяет предположить либо преобладание “общего”, либо переходного между “общим” и “детальным” режимами визуального восприятия у детей. В пользу данного предположения можно привести то, что при задействовании рабочей памяти у детей и взрослых наблюдается отличие в активации зон головного мозга. Например, в исследовании V.M. Vogan et al. показано, что, несмотря на общую схожесть паттерна нейронной активации у взрослых людей при увеличении нагрузки на вербальную рабочую память отмечено значительное снижение активности в двусторонней передней поясной извилине, передней медиальной префронтальной извилине, правой верхней боковой височной извилине и левой задней поясной извилине [18]. Результаты эксперимента по влиянию на качество рабочей памяти удаления части префронтальной коры у обезьян разного возраста показали, что процесс регуляции рабочей памяти может сравнительно отличаться на разных этапах онтогенеза [19]. Таким образом, исследования когнитивных процессов у взрослых нельзя с полной уверенностью экстраполировать на детей.
При интерпретации изображения немаловажен процесс разделения фоновой и целевой информации, который во многом определяется качеством предварительной оценки релевантности информации на уровне периферического зрения [17]. Показателем дефицита периферического анализа может служить точность саккад, которую довольно сложно оценить при выполнении произвольного рассматривания. В результатах данного исследования саккад, обращенных на фон, крайне мало, что, возможно, объясняется особенностями самого изображения, которые не предполагают сдвиг внимания на фоновую часть. Однако наличие в W, NoA и RwP большего количества малых групп фиксаций, по сравнению с группой R, позволяет предположить, что при неоптимальном окуломоторном паттерне дети совершают больше возвратных (возвращающихся к уже просмотренной зоне) саккад, и возможно вследствие недостаточной точности саккад [20]. Такая особенность (множество небольших групп фиксаций) может отражать адаптивный процесс, направленный на интерпретацию недопонятых частей изображения.
При свободном рассматривании средняя продолжительность фиксаций коррелирует с объемом обрабатываемой информации [21]. Исходя из этого, можно предположить, что снижение продолжительности фиксаций в группах R и W может свидетельствовать о завершении процесса анализа информации, увеличение времени фиксаций в группе RwP может отражать мобилизацию когнитивных ресурсов, а отсутствие изменений в группе NoA – о продолжающемся процессе обработки информации, который, впрочем, малоэффективен. Предполагая, что функциональная асимметрия между правой (детальный анализ) и левой (общий анализ) частями поля зрения носит врожденный характер, отсутствие статистически значимых различий во времени фиксации в пределах разных частей поля зрения может отражать несформированность управляющих функций.
Среди прочих моделей регуляции движений глаз, перспективной представляется та, согласно которой характер движений глаз определяется взаимодействием между процессами фокальной и периферической обработки зрительной информации [22–24]. В работе A. Helo et al. [10] формирование двух стратегий обработки зрительной информации отмечается к 2–3 годам. Первая стратегия в большей степени связана с периферическим зрением и процессами пространственного ориентирования, общим анализом информации. Для такой стратегии характерны зрительные фиксации с продолжительностью менее 180 мс и саккады с амплитудой менее 5 угл. град. Вторая стратегия связана с реализацией локального анализа информации и категоризации. При этом процесс общего анализа оптимальнее осуществляется в правой части поля зрения, а локальный анализ – в левой части поля зрения [22]. Наблюдаемая асимметрия, вероятно, может подвергаться воздействию ряда внешних факторов и приобретенного опыта.
Опираясь на полученные данные сложно утверждать о преобладании какой-либо из описанных стратегий. Можно с осторожностью предположить, что в группах R и W преобладает стратегия анализа зрительной информации по локальному типу, т.к. зрительные фиксации распределены более “скученно” по сравнению с группами RwP и NoA. Примечательная для группы W горизонтальная ориентация окуломоторного паттерна может свидетельствовать о нарушении внимания, так как подобный паттерн был обнаружен в группе детей с синдромом дефицита внимания и гиперактивности [3]. Формы окуломоторных паттернов в группах R, RwP и NoA малоразличимы на первый взгляд. Однако анализ состава фиксационных групп показал явное отличие группы R от W, RwP, NoA. Помимо этого, для групп обнаружена различная динамика продолжительности фиксаций (отрицательная в R и положительная в RwP).
Влияние утомления мы исключаем ввиду малой продолжительности предъявления стимульного материала и соответствия проведения исследования периоду дневной активности детей. С другой стороны, на движениях глаз участников могло повлиять беспокойство или ситуативная тревожность при выполнении задания [25]. Изменение диаметра зрачка при фиксациях не коррелирует с состоянием рабочей памяти в случае, если отведенное на рассматривание время больше пяти секунд, но, вероятно, отражает лишь общее влияние когнитивной нагрузки [21]. Положительная динамика размера зрачка при рассматривании в группах R, W и RwP показывает, что участники этих групп, по сравнению с группой NoA, могут иметь более высокие когнитивные способности в отношении объема рабочей памяти и интеллектуальной гибкости [26]. С другой стороны, увеличение диаметра зрачка может отражать степень прилагаемых усилий при осуществлении когнитивного контроля [27]. Для подтверждения либо опровержения этих предположений необходимо дополнительное исследование.
ЗАКЛЮЧЕНИЕ
В результате проведенного исследования показано, что наблюдается тенденция к снижению скорости саккад и увеличению продолжительности зрительных фиксаций с возрастом, что предположительно может характеризовать процесс созревания коры больших полушарий, а также ствола головного мозга и мозжечка. Динамика значений по различным окуломоторным параметрам имеет нелинейный характер, что с высокой вероятностью обусловлено гетерохронностью развития головного мозга. Выявленные группы детей, разделенных по их способности ответить на вопрос о просмотренном изображении (что косвенно свидетельствует об уровне сформированности когнитивных функций) имеют характерный окуломоторный профиль. Результаты данного исследования дают возможность более точно характеризовать когнитивные функции в “фоновом режиме”. Результаты проведенного исследования могут внести вклад в понимание возрастной динамики саккад и послужить для создания базы контрольных значений окуломоторной активности в норме, что может оказаться полезным в области клинических исследований нейропатологий. При дальнейшей работе по данной теме планируется проведение лонгитюдного исследования с привлечением методов нейровизуализации с целью развития нейропедагогического направления в обучении.
Этические нормы. Все исследования проведены в соответствии с принципами биомедицинской этики и одобрены независимым этическим комитетом при Национальном медицинском исследовательском центре психиатрии и неврологии им. В.М. Бехтерева (Санкт-Петербург).
Информированное согласие. Перед началом исследования каждый родитель участника представил добровольное письменное информированное согласие, подписанное им после разъяснения ему потенциальных рисков и преимуществ, а также характера предстоящего исследования.
Конфликт интересов. Авторы декларируют отсутствие явных и потенциальных конфликтов интересов, связанных с публикацией данной статьи.
Вклад авторов в публикацию. Е.А. Буденкова – проведение эксперимента, обработка данных, написание статьи. Д.А. Швайко – написание статьи.
Список литературы
Rommelse N.N.J., Van der Stigchel S., Sergeant J.A. A review on eye movement studies in childhood and adolescent psychiatry // Brain Cogn. 2008. V. 68. № 3. P. 391.
Frey A. Eye movements in children during reading: a review // Actes du Symposium international sur la litéracie à l’école / International Symposium for Educational Literacy (SILE/ISEL), 2015. P. 316.
Hayes T.R., Henderson J.M. Scan patterns during scene viewing predict individual differences in clinical traits in a normative sample // PLoS One. 2018. V. 13. № 5. P. e0196654.
Henderson J.M. Gaze control as prediction // Trends Cogn. Sci. 2017. V. 21. № 1. P. 15.
Krasich K., Huffman G., Faber M., Brockmole J.R. Where the eyes wander: The relationship between mind wandering and fixation allocation to visually salient and semantically informative static scene content // J. Vis. 2020. V. 20. № 9. P. 10.
Leigh R.J., Zee D.S. The neurology of eye movements. OUP USA, 2015. 1136 p.
Liu Z., Yang Z., Gu Y. et al. The effectiveness of eye tracking in the diagnosis of cognitive disorders: A systematic review and meta-analysis // PLoS One. 2021. V. 16. № 7. P. e0254059.
Eckstein M.K., Guerra-Carrillo B., Singley A.T.M. et al. Beyond eye gaze: What else can eyetracking reveal about cognition and cognitive development // Dev. Cogn. Neurosci. 2017. V. 25. P. 69.
Walter K., Bex P. Cognitive load influences oculomotor behavior in natural scenes // Sci. Rep. 2021. V. 11. № 1. P. 12405.
Helo A., Pannasch S., Sirri L. et al. The maturation of eye movement behavior: scene-viewing characteristics in children and adults // Vision Res. 2014. V. 103. P. 83.
Le Meur O., Coutrot A., Liu Z. et al. Visual attention saccadic models learn to emulate gaze patterns from childhood to adulthood // IEEE Trans. Image Process. 2017. V. 26. № 10. P. 4777.
Хрипкова А.Г. Возрастная физиология и школьная гигиена. М.: Просвещение, 1990. 319 с.
Безруких М.М., Фарбер Д.А. Физиология развития ребенка. Руководство по возрастной физиологии. М.: Изд-во Моск. психол.-соц. ин-та, 2010. 768 с.
Luna B., Velanova K., Geier C.F. Development of eye-movement control // Brain Cogn. 2008. V. 68. № 3. P. 293.
Krauzlis R.J., Goffart L., Hafed Z.M. Neuronal control of fixation and fixational eye movements // Philos. Trans. R. Soc. B: Biol. Sci. 2017. V. 372. № 1718. P. 20 160 205.
Pannasch S., Helmert J.R., Roth K. et al. Visual fixation durations and saccade amplitudes: Shifting relationship in a variety of conditions // J. Eye Mov. Res. 2008. V. 2. № 2. P. 1.
Ito J., Yamane Y., Suzuki M. et al. Switch from ambient to focal processing mode explains the dynamics of free viewing eye movements // Sci. Rep. 2017. V. 7. № 1. P. 1082.
Vogan V.M., Morgan B.R., Powell T.L. et al. The neurodevelopmental differences of increasing verbal working memory demand in children and adults // Dev. Cogn. Neurosci. 2016. V. 17. P. 19.
Goldman P.S., Rosvold H.E., Vest B. et al. Analysis of the delayed-alternation deficit produced by dorsolateral pre-frontal lesions in the rhesus monkey // J. Comp. Physiol. Psychol. 1971. V. 77. № 2. P. 212.
Chow-Wing-Bom H., Dekker T.M., Jones P.R. The worse eye revisited: evaluating the impact of asymmetric peripheral vision loss on everyday function // Vision Res. 2020. V. 169. P. 49.
Meghanathan R.N., van Leeuwen C., Nikolaev A.R. Fixation duration surpasses pupil size as a measure of memory load in free viewing // Front. Hum. Neurosce. 2015. V. 8. P. 1063.
Karim A.K.M.R., Kojima H. The what and why of perceptual asymmetries in the visual domain // Adv. Cogn. Psychol. 2010. V. 6. P. 103.
Torralba A., Oliva A., Castelhano M.S. et al. Contextual guidance of eye movements and attention in real-world scenes: the role of global features in object search // Psychol. Rev. 2006. V. 113. № 4. P. 766.
Kanan C., Tong M.H., Zhang L. et al. SUN: top-down saliency using natural statistics // Vis. Cogn. 2009. V. 17. № 6–7. P. 979.
Lisk S., Vaswani A., Linetzky M. et al. Systematic review and meta-analysis: eye-tracking of attention to threat in child and adolescent anxiety // J. Am. Acad. Child Adolesc. Psychiatry. 2020. V. 59. № 1. P. 88.
Tsukahara J.S., Harrison T.L., Engle R.W. The relationship between baseline pupil size and intelligence // Cogn. Psycho. 2016. V. 91. P. 109.
van der Wel P., van Steenbergen H. Pupil dilation as an index of effort in cognitive control tasks: A review // Psychon. Bull. Rev. 2018. V. 25. № 6. P. 2005.
Дополнительные материалы отсутствуют.
Инструменты
Физиология человека