

Доклады Российской академии наук. Науки о Земле, 2020, T. 493, № 2, стр. 88-92
СообществА Portlandia арктических морей: таксономические и структурные различия на географическом градиенте
В. О. Мокиевский 1, *, А. А. Удалов 1, А. И. Чава 1, У. В. Симакова 1, В. Н. Кокарев 2, Т. В. Неретина 3
1 Институт океанологии им П.П. Ширшова
Российской академии наук
Москва, Россия
2 Факультет биологических наук и аквакультуры, Университет Норд
Буде, Норвегия
3 Беломорская биологическая станция им. Н. А. Перцова биологического факультета МГУ им. М. В. Ломоносова
Москва, Россия
* E-mail: vadim@ocean.ru
Поступила в редакцию 25.05.2020
После доработки 01.06.2020
Принята к публикации 02.06.2020
Аннотация
Проанализирован видовой состав и структура донных сообществ с преобладанием двустворчатых моллюсков рода Portlandia, широко распространенных на шельфе Российской Арктики. Молекулярно-генетический анализ подтвердил самостоятельность видов Portlandia aestuariorum и P. arctica и показал, что все образцы P. arctica из разных регионов принадлежат к одному виду независимо от морфологических вариаций. Описано 10 различных вариантов бентосных сообществ из Белого, Карского, Лаптевых и Восточно-Сибирского морей, в которых P. aestuariorum и P. arctica играют доминирующую роль. Для видов Portlandia не выявлено стабильных сопутствующих видовых комплексов. Сообщества практически не имеют общих видов, их состав и структура определяются подбором видов со сходными экологическими предпочтениями из состава локальной фауны. Различия сообществ определяются как географическими, так и экологическими причинами.
Одним из широко распространенных и традиционных подходов выделения сообществ организмов является объединение описаний (станций) по доминирующему виду [1]. Все описания с одним и тем же доминантом считаются относящимися к одному сообществу. В случае видов с широким распространением этот подход может приводить к объединению ассоциаций видов, различных по видовому составу и происхождению. В то же время анализ состава и структуры сообществ с одним и тем же доминантом служит хорошим объектом для изучения проблемы сопряженности видов в донных сообществах и оценки роли межвидовых взаимодействий в организации бентоса.
Преобладающий по биомассе вид может выступать в качестве вида-эдификатора, трансформирующего среду обитания для сопутствующих видов, однако это происходит далеко не всегда и не во всех сообществах. Вопрос о степени сопряженности видов и о регулирующей роли вида-доминанта слабо изучен для сообществ инфауны [2]. Исследование географически удаленных сообществ с одним и тем же доминантом позволяет оценить его роль в качестве эдификатора и выявить набор сопряженных видов, если такие существуют.
Сообщества с доминированием двустворчатого моллюска Portlandia arctica могут служить хорошим объектом для количественной оценки географических различий в однотипных сообществах и анализа связи между доминирующим видом и его окружением.
P. arctica имеет циркумарктическое распространение, она обычна во всех морях арктического шельфа Евразии и часто выступает в роли доминирующего вида в сообществах илистых грунтов [3–7, 11].
Материалом для анализа послужили количественные дночерпательные сборы экспедиций на судах ИО РАН “Академик Мстислав Келдыш” и “Профессор Штокман” из Карского, Лаптевых и Восточно-Сибирского морей, и сборы Беломорской прибрежно-морской экспедиции ИО РАН в сотрудничестве с Беломорской биостанцией МГУ из Белого моря (2007–2017 гг.). В анализ включены данные об обилии макробентоса для 61 станции, на которых виды рода портландия входили в число доминирующих или характерных (рис. 1).
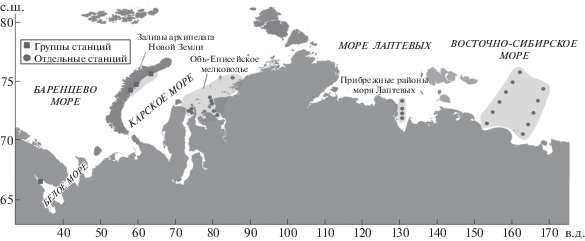
Таксономический состав рода Portlandia до сих пор служит предметом дискуссий. Морфологическая изменчивость раковин портландий в разных частях ареала привела к выделению большого числа морфологических форм разного таксономического ранга [4]. Прежде чем решать задачу о составе и структуре однотипных сообществ на протяженном географическом градиенте, следовало убедиться в том, что во всех случаях доминирующая форма относится к одному и тому же виду.
Для обоих морфовидов портландий (P. aestua-riorum, 10 образцов, эстуарии Оби, Енисея и Хатанги, и P. arctica, отнесенных к трем варьететам: var. typica, var. portlandica, var. siliqua, – 12 образцов из Карского, Лаптевых, Белого и восточно-Сибирского морей) был проведен анализ последовательностей участка митохондриального гена COI. Для сравнения использовали данные, опубликованные в базе данных BOLD [8], о моллюсках из Канадской Арктики (P. arctica, Yoldia hyperborea). Результаты филогенетической реконструкции (RAxML, 1000 bootstrap [9]) говорят о дивергенции двух видов независимо от географического положения образцов. Средние эволюционные дистанции [10] между морфовидами (0.089 ± 0.0160, M ± S.E.) заметно превышали внутригрупповые дистанции (P. aestuariorum – 0.001 ± 0.0008, P. arctica – 0.007 ± 0.002). Из чего можно сделать вывод, что выделенная первоначально в статусе подвида, а затем переведенная в ранг вида форма P. aestuariorum Mosevich, 1924 является самостоятельным видом. Все образцы морфовида P. arctica из разных регионов, в том числе и из Канадской Арктики принадлежат к одному виду независимо от характерных для них морфологических вари-аций.
Анализ структуры сообществ (сходство видового состава, сходство структуры выборок по соотношениям видов, состав доминирующей группы) позволяет разделить всю нашу выборку на несколько самостоятельных комплексов. Отчетливо отделяются все станции с доминированием Portlandia aestuariorum. Сообщества с доминированием этого вида описаны на нашем материале из опресненных районов эстуариев Лены и Енисея при солености 18–26 е.п.с. В их составе преобладают виды солоноватоводного комлекса, такие как Marenzelleria arctia и Mesidotea sabini в эстуарии Енисея, Prionospio cirrifera в приустьевом районе р. Лена (табл. 1). Состав и структура сообществ P. aestuariorum в двух исследованных районах сильно различаются, хотя их объединяет низкое видовое богатство (табл. 1). Из руководящих видов только Mesidotea sibirica присутствует в обоих районах.
Таблица 1.
Характеристики сообществ Portlandia arctica в разных районах
| Район | Глубина, м | N, экз/м2 | B, г/м2 | Число видов | Доминирующий видовой комплекс | ||
|---|---|---|---|---|---|---|---|
| всего | на пробу 0.1 м2 | ES(100) | |||||
| Енисейский залив, опресненная часть (Карское море) | 13–14 | 2388 | 14 | 23 | 17 | 9 | Marenzelleria arctia, Portlandia aestuariorum, Mesidotea sabini |
| Придельтовый район р. Лена, опресненная часть (море Лаптевых) | 10–12 | 872 | 30 | 27 | 17 | 9 | Portlandia aestuariorum, Prionospio cirrifera |
| Пролив Великая Салма, Кандалакшский залив (Белое море) | 69–133 | 1949 | 106 | 75 | 29 | 15 | Portlandia arctica, Galathowenia oculata, Aricidea nolani, Scoloplos sp., Pectinaria hyperborea, Maldane sarsi, Nuculana pernula |
| Полуизолированная обедненная лагуна Бабье море (Белое море) | 11–31 | 1117 | 25 | 38 | 12 | 8 | Portlandia arctica, Prionospio cirrifera, Aricidea nolani |
| Обская губа, морская часть (Карское море) | 15–21 | 1196 | 17 | 32 | 20 | 12 | Portlandia arctica, Trochochaeta carica, Micronephthys minuta, Terebellides stroemii, Cossura spp. |
| Енисейский залив и Енисейское мелководье (Карское море) | 30–53 | 1086 | 58 | 69 | 36 | 15 | Portlandia arctica, Mesidotea sabini, Cirratulidae gen. sp., Pectinaria hyperborea, Micronephthys minuta, Pandora glacialis, Macoma calcarea |
| Внутренняя часть заливов архипелага Новая Земля (Карское море) | 56–138 | 1393 | 55 | 56 | 22 | 14 | Ennucula tenuis, Portlandia arctica, Scoletoma fragilis, Mendicula ferruginosa, Cirratulidae gen. sp. |
| Заливы архипелага Новая Земля, терминальные станции (Карское море) | 21–109 | 402 | 22 | 13 | 8 | 7 | Portlandia arctica |
| Прибрежные районы Лаптевых и Восточно-Сибирского морей | 17–32 | 2937 | 81 | 72 | 36 | 17 | Portlandia arctica, Mesidotea sabini, Artacama proboscidea, Cirratulidae gen. sp., Praxillella praetermissa, Aglaophamus malmgremi, Thyasiragouldi, Lyonsia arenosa |
| Восточно-Сибирское море, основная часть | 13–50 | 1024 | 45 | 68 | 19 | 9 | Portlandia arctica, Mesidotea sabini |
Сообщества с доминированием Portlandia arctica представлены в нашем материале сборами из двух районов Белого моря, двух районов Карского моря, из моря Лаптевых и из Восточно-Сибирского моря. Все исследованные сообщества формируются в достаточно широком диапазоне температуры (от +5 до –1.5°С) и солености (24–34 е.п.с.), на мягких илистых грунтах в диапазоне глубин от 10 до 140 м. Тем не менее, следует заметить, что большинство из них обитают в условиях пониженной солености и на небольших глубинах, и только немногие варианты формируются в глубоких (до 150 м) котловинах заливов восточного побережья Новой Земли с высокой (34.5 е.п.с.) соленостью. На нашем материале выделяется 8 основных комплексов, различающихся структурными характеристиками и наборами руководящих видов (табл. 1). В их видовом составе практически нет общих видов, из 250 найденных нами видов лишь несколько (менее 10) (Micronephthys minuta, Terebellides stroemii, Macoma calcarea, Cirratulidae gen. sp., Cossura spp.) отмечены во всех районах, но далеко не всегда – в составе руководящего комплекса. Различия в видовом составе сообществ из разных морей и районов моря определяются как географическими, так и экологическими причинами. Ряд видов (Golfingia margaritacea, Ophiopleura borealis, Yoldiella lenticula, Mendicula ferruginosa, Yoldiella solidula, Bathyarca glacialis) характерны только для глубоководных вариантов сообщества, описанных из заливов архипелага Новая Земля. В районах с пониженной соленостью в состав сообщества P. arctica входит ряд мезогалинных видов (Trochochaeta carica, Aulodrilus pluriseta, Halicryptus spinulosus, Aulodrilus pigueti, Marenzelleria arctia), часть из них встречается также и в составе сообществ Portlandia aestuariorum.
Географические различия в составе сообществ проявляются не менее отчетливо. В сообществах Белого моря происходит обеднение состава за счет выпадения ряда арктических форм, широко распространенных в остальных морях – Bathyarca glacialis, Mendicula ferruginosa, Mesidotea sabini, Ophiocten sericeum, Ophiopleura borealis. Ряд видов характерен только для западных вариантов сообщества портландии и отсутствует в Восточно-Сибирском море (Maldane sarsi, Scoletoma fragilis, Pectinaria hyperborea, Aricidea nolani). В Белом море в составе сообществ портландии многочисленны мелководные бореальные виды (Galathowenia oculata, Nuculana pernula, Yoldia hyperborea).
Структурные характеристики сообществ с участием P. arctica также отличаются от района к району (табл. 1). Средние значения биомассы и численности различаются в различных вариантах сообществ в два–три раза, видовое богатство – втрое. При этом наиболее низкое разнообразие (7–9 видов на 100 найденных экземпляров) отмечено в краевых сообществах – в полуизолированных и кутовых частях заливов, а также в наиболее географически изолированном восточном варианте сообщества – в Восточно-Сибирском море (табл. 1).
Мы не видим в составе сообществ портландии характерных видов, которые были бы тесно связаны по своей встречаемости с этими моллюсками и уникальных именно для данных сообществ. Напротив, состав субдоминантов отличается в каждом море и биотопе, формируясь из числа широко распространенных эврибионтных видов, распространенных в соседних сообществах.
Все сообщества портландии приурочены к биотопам, появившимся в голоцене. На пике последнего оледенения районы современного распространения сообществ портландии были покрыты льдом (Белое море, заливы архипелага Новая Земля) или находились выше уровня моря (Объ-Енисейское мелководье, мелководья морей Восточно-Сибирского и Лаптевых). Набор видов при формировании сообщества в каждом случае определялся составом региональной фауны и локальными условиями биотопа.
Сравнительный анализ сообществ Portlandia arctica из разных районов Арктики и широкого спектра биотопов показал, что наличие постоянного характерного или доминирующего вида остается единственной общей чертой, объединяющей группировки видов, различные по таксономическому составу и структуре. Даже те немногие виды, которые входят во все исследованные комплексы P. arctica (около 10 видов), оказываются эврибионтными видами циркумарктического распространения, характерными для целого ряда соседних сообществ. Роль Portlandia arctica в качестве эдификатора, определяющего облик сообщества и подбор видов, не выявляется, состав и структура сообществ во всех биотопах формируется подбором видов со сходными экологическими предпочтениями из состава локальной фауны.
Список литературы
Воробьев В.П. Бентос Азовского моря // Тр. АзЧерНИРО. 1949. Т. 13. С. 1–193.
Мокиевский В.О., Цетлин А.Б. Многолетняя динамика донных сообществ // Зоологический журнал. 2020. Т. 99. № 7.
Наумов А.Д., Мокиевский В.О., Исаченко А.И., Савченко О.Н., Биягов К.Л., Аристов Д.А. Сублиторальные сообщества макробентоса Бабьего моря. В кн.: Комплексные исследования Бабьего моря, полу-изолированной беломорской лагуны: геология, гидрология, биота – изменения на фоне трансгрессии берегов (Труды Беломорской биостанции МГУ, т. 12) / В.О. Мокиевский, А.И. Исаченко, П.Ю. Дгебуадзе, А.Б. Цетлин (ред.). М.: Т-во научных изданий КМК. 2016. С. 122–143.
Филатова З.А. Некоторые зоогеографические особенности двустворчатых моллюсков рода Portlandia // Тр. ИО АН СССР. 1951. Т. VI. С. 117–131.
Филатова З.А., Зенкевич Л.А. Количественное распределение донной фауны Карского моря // Тр. Всесоюзного гидробиологического общества. 1957. Т. VIII. С. 3–67.
Aitken A.E., Fournier J. Macrobenthic Communities of Cambridge, McBeth and Itirbilung Fjords, Baffin Island, Northwest Territories, Canada // Arctic. 1993. V. 46. № 1. P. 60–71.
Naumov A.D., Fedyakov V.V. New Results of the Macro-zoobenthos of the White Sea Deep Basin // Berichte zur Polarforshung. 2000. Bd. 359. P. 54–96.
Ratnasingham S., Hebert P. D. N. The Barcode of Life Data System (www. barcodinglife. org). Molecular Ecology Notes. 2007. V. 7. № 3. P. 355–364.
Stamatakis A. RAxML Version 8: A Tool for Phylogenetic Analysis and Post-analysis of Large Phylogenies // Bioinformatics. 2014. V. 30. № 9. P. 1312–1313.
Tamura K., Kumar S. Evolutionary Distance Estimation under Heterogeneous Substitution Pattern among Lineages // Molecular Biology and Evolution. 2002. V. 19. P. 1727–1736.
Thorson G. Investigations on Shallow Water Animal Communities in the Franz Joseph Fjord (East Greenland) and Adjacent Waters // Meddelelser om Grønland. 1933. V. 100. № 2. P. 1–70.
Дополнительные материалы отсутствуют.
Инструменты
Доклады Российской академии наук. Науки о Земле