

Доклады Российской академии наук. Науки о Земле, 2023, T. 510, № 1, стр. 61-65
Биота эдиакарского типа в верхнем докембрии Тиманского кряжа (возвышенность Джежим-Парма, республика Коми)
А. В. Колесников 1, *, И. В. Латышева 1, А. В. Шацилло 1, 2, член-корреспондент РАН Н. Б. Кузнецов 1, А. С. Колесников 1, В. Д. Десяткин 1, Т. В. Романюк 2
1 Геологический институт Российской академии наук
Москва, Россия
2 Институт физики Земли им. О.Ю. Шмидта Российской академии наук
Москва, Россия
* E-mail: kolesnikov@ginras.ru
Поступила в редакцию 27.12.2022
После доработки 09.01.2023
Принята к публикации 10.01.2023
- EDN: FFZWNN
- DOI: 10.31857/S2686739722602964
Аннотация
В джежимской свите Тиманского кряжа впервые обнаружены макроскопические палеонтологические остатки – объемные слепки и отпечатки мягкотелых организмов эдиакарского типа. Среди них диагностированы представители палеопасцихнид, фрондоморф, чуариоморфных колоний, арумбериеморфных текстур, а также ископаемые следы жизнедеятельности. Обнаружение большого количества разнообразных палеонтологических остатков эдиакарских организмов на Тиманском кряже не только расширяет представление об их палеогеографическом ареале расселения, но также уточняет временные рамки накопления джежимской свиты, положение которой в разрезе верхнего докембрия было спорным.
Тиманский кряж расположен на северо-востоке европейской̆ части России. Протягиваясь с северо-запада на юго-восток, он состоит из ряда возвышенностей высотой от 300 до 450 м, ограничивающих Печорскую низменность с юго-запада. По характеру строения и полноте разрезов верхнего докембрия в пределах Тиманского кряжа выделен ряд зон (с юго-запада на северо-восток): Обдырско-Нившерская, Четласско-Джежимпарминская, Цильменско-Ропчинская и Вымско-Вольская [1]. В Четласско-Джежимпарминской зоне верхний докембрий объединен в четласскую и быстринскую серии. Первая из них некоторыми авторами отнесена к среднему рифею [2], а другими авторами – к верхнему рифею–(?)венду [1]. В сводном разрезе четласской серии выделяют светлинскую, новобобровскую и визингскую свиты. Эта серия с несогласием перекрыта быстринской серией, расчлененной на устьпалегскую (анъюгскую, джежимскую), ворыквинскую, павъюгскую и паунскую свиты.
Возвышенность Джежим-Парма расположена на юго-востоке Тиманского кряжа. Здесь, в обрамлении палеозойских комплексов, развиты песчаники и алевролиты визингской свиты, песчаники джежимской свиты, известняки (местами доломитизированные) и алевро-аргиллиты павъюгской свиты [3]. Соотношения между свитами тектонические, характер разрывных нарушений неизвестен [4]. На протяжении долгого времени возраст и стратиграфическое положение этих свит были дискуссионны. Так, В.Г. Оловянишников считал, что доломиты павъюгской свиты представляют древнейшую часть разреза, и относил их к верхнему рифею, а джежимскую свиту сопоставлял с анъюгской свитой венда [3]. Однако в легенде второго издания Тиманской серии листов ГГК-200 РФ [5] джежимская свита помещена в основание разреза и отнесена к верхнему рифею.
В 2021–2022 гг. в карьере Асыввожского месторождения бутового камня (рис. 1) мы провели рекогносцировочные исследования красноцветных силикокластических пород джежимской свиты, где впервые обнаружили многочисленные остатки ископаемых организмов эдиакарского типа (рис. 2).
Рис. 1.
Схема расположения нового местонахождения остатков организмов эдиакарского типа на северо-восточном обрамлении Восточно-Европейской платформы.
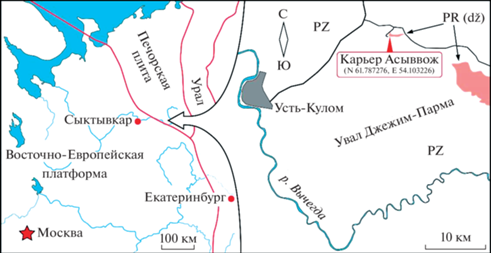
Рис. 2.
Представители ископаемой биоты эдиакарского типа из джежимской свиты Южного Тимана: а – обр. № ST22/1-55, Palaeopascichnus linearis; б – ST22/1-67, P. linearis; в – обр. № ST22/1-62, P. gracilis; г – обр. № ST22/1-64, P. delicatus; д – обр. № ST22/1-71, Aspidella sp.; е – обр. № ST22/1-42, Aspidella sp.; ж – обр. № ST22/1-72, Beltanelliformis minutae; з – обр. № ST22/1-34, Helminthoidichnites isp.; и – обр. № ST22/1-68, H. tenuis (?); й – обр. № ST22/1-73, Arumberia vindhyanensis; к – обр. № ST22/1-61, A. banksi. Коллекция лаборатории стратиграфии верхнего докембрия Геологического института РАН (г. Москва).

Общая видимая мощность свиты, вскрытая карьером, составляет ~70 м. Свита сложена красноцветными, преимущественно косо- и волнистослоистыми, песчаниками с редкими пакетами переслаивающихся алевролитов и песчаников мощностью до 50 см. Палеонтологические остатки приурочены к одному из таких пакетов на отметке 12 м от основания видимой части разреза джежимской свиты. Собранная нами коллекция состоит из 84 экземпляров представителей палеопасцихнид, органов прикрепления фрондоморфных организмов, чуариоморф, ископаемых следов жизнедеятельности и микробиальных колоний.
Таксономически наиболее разнообразные палеопасцихниды представлены в виде однорядных серий выпуклых слепков камер на подошвах алевролитов и мелкозернистых песчаников (позитивный гипорельеф), имеющих глобулярную, вытянутую или аллантоидную (выгнутую) форму, расположенных цепочкообразно друг за другом (рис. 2 а–д). В большинстве случаев палеопасцихнид можно идентифицировать только с помощью узконаправленного источника света, расположенного под низким углом к поверхности слоистости. Это обусловлено тем, что палеонтологические остатки низкорельефные и демонстрируют степень сохранности хуже, чем, например, схожая ассоциация из вилухинской подсвиты чернокаменской свиты венда Среднего Урала [6]. Среди палеопасцихнид диагностированы три вида: Palaeopascichnus linearis, состоящий из однорядных серий глобулярных или слегка вытянутых камер шириной от 3 до 6 мм (рис. 2 а, б); P. gracilis, состоящий из однорядных серий сильно вытянутых камер длиной до 1 мм и шириной до 25 мм (рис. 2 в); P. delicatus, состоящий из однорядных серий камер, имеющих аллантоидную форму и прогрессивно увеличивающуюся ширину (рис. 2 г).
Органы прикрепления представлены позитивными гипорельефными луковицевидными слепками или дисковидными отпечатками в алевролитах и мелкозернистых песчаниках (рис. 2 д, е). Диаметр луковицевидных тел и дисков варьирует от 1 до 3 см, а высота гипорельефа достигает 5 мм. Степень сохранности не позволяет однозначно установить видовую принадлежность остатков, однако, подобные формы широко распространены в синекаменской подсвите чернокаменской свиты верхнего венда Среднего Урала и определены как слепки органов прикрепления фрондоморфных организмов Aspidella Billings [7].
Остатки чуариоморфных колоний наиболее многочисленны в джежимской свите. Они представлены скоплениями небольших округлых или эллиптических слепков без каких-либо видимых концентрических или радиальных складок в позитивном гипорельефе алевролитов и мелкозернистых песчаников (рис. 2 ж). Диаметр отдельных экземпляров редко превышает 7–10 мм, а высота гипорельефа обычно менее 1 мм. Характер площадного распределения и морфология отдельных экземпляров идентичны чуариоморфным колониям Beltanelliformis konovalovi из чернокаменской свиты Среднего Урала [8], хотя последние демонстрируют существенное различие в размерах особей. Ископаемые формы, подобные асыввожским, ранее были описаны как остатки мягкотелых колониальных организмов Beltanelliformis minutae из серии лонгминдиан эдиакария Англии [9], однако позже другими исследователями была высказана точка зрения об их принадлежности к микробиально-индуцированным осадочным текстурам [10]. С другой стороны, недавно были получены убедительные доказательства цианобактериальной природы отдельных экземпляров Beltanelliformis из верхнего венда юго-восточного Беломорья [11], соответственно ничто не мешает рассматривать многочисленные скопления округлых слепков в джежимской свите как специфические остатки микробных (чуариоморфных) колоний.
Ископаемые следы жизнедеятельности представлены на поверхностях напластования мелкозернистых песчаников и алевролитов простыми горизонтальными неветвящимися меандрирующими валиками (позитивный гипорельеф) или канавками (негативный эпирельеф) шириной 1–3 мм (рис. 2 з, и), которые интерпретированы нами, как заполненные осадком остатки нор. Поверхность нор ровная, без каких-либо видимых пережимов и морфологических структур. Наблюдаемые признаки в достаточной степени весомы для определения этих остатков до ихнорода Helminthoidichnites isp., которые широко распространены в ископаемой летописи начиная с отложений позднего эдиакария [12].
Помимо всего, в джежимской свите распространены проблематики и различные микробиальные образования, среди которых встречены арумбериеморфные текстуры, также широко известные в литературе под родовым именем “Arumberia” (рис. 2 й, к). На протяжении длительного времени их принимали за эрозионные текстуры, однако, результаты последних исследований убедительно показали их биологическое происхождение [13–15]. В джежимской свите встречены две разновидности арумбериеморфных текстур: Arumberia vindhyanensis (рис. 2 й) – серии субпараллельных неглубоких канавок или невысоких валиков (до 0.7 мм), расположенных близко друг к другу, пространство между которыми рассечено более мелкими канавками/валиками; A. banksi (рис. 2 к) – серии относительно крупных субпараллельных и веерообразно расходящихся канавок (негативный эпирельеф) и валиков (позитивный гипорельеф) шириной до 2 мм и глубиной/высотой до 2 мм на подошве и внутри песчаных слоев.
До настоящего времени находки ископаемых организмов эдиакарского типа на северо-восточном обрамлении Восточно-Европейской платформы не были известны, а джежимскую свиту Южного Тимана относили к верхнему рифею [1, 2, 5]. Обнаружение остатков организмов эдиакарского типа однозначно указывает на вендский возраст свиты. Так, в пользу более молодого (пострифейского) возраста указывают находки ихнофоссилий Helminthoidichnites isp., известные в ископаемой летописи начиная с временной отметки ~ 560 млн лет [12], а также отпечатки микробиальных сообществ типа Arumberia banksi, время существования которых в ископаемой летописи ограничено интервалом 560–520 млн лет [16]. Исключение составляют современные проявления арумбериеморфных образований на поверхностях галотолерантных цианобактериальных матов в крайне мелководных водоемах на приливно-отливных равнинах [14]. Важными находками являются палеопасцихниды, стратиграфический диапазон которых соответствует всему объему вендской системы [17]. Более того, нам удалось обнаружить ассоциацию палеопасцихнид, включающую в себя виды Palaeopascichnus delicatus, P. gracilis и P. linearis, время существования которой ограничено интервалом 560–550 млн лет [18] и соответствует, в современном понимании, беломорскому ярусу верхнего венда Восточно-Европейской платформы [19].
Несмотря на то что качество сохранности макрофоссилий Тиманского кряжа уступает сохранности аналогичных организмов из крупнейших местонахождений мира, комплекс остатков эдиакарского типа из джежимской свиты по разнообразию таксонов и многочисленности можно сравнить с ископаемой чернокаменской биотой венда Среднего Урала [7]. Подобно фоссилиеносным уровням чернокаменской свиты Среднего Урала [6–8, 20], палеонтологические остатки из джежимской свиты приурочены к отложениям крайнемелководных обстановок приливно-отливных равнин с признаками периодического осушения (наличие арумбериеморфных текстур, знаков волновой ряби, трещин усыхания и слепков мелких кубических кристаллов соли).
В таксономическом смысле представленный материал пока недостаточно богат для детальных исследований в области морфологии, тафономии и палеоэкологии. Тем не менее обнаружение первых макрофоссилий в джежимской свите дает перспективу для поиска новых палеонтологических остатков и открытия полноценного местонахождения ископаемых организмов эдиакарского типа в регионе и указывает на то, что палеонтологический потенциал венда северо-востока Восточно-Европейской платформы еще не раскрыт. Таким образом, разрезы верхнего докембрия Тиманского кряжа обретают статус ключевых объектов для изучения функциональных особенностей палеоэкосистем на ранних этапах эволюции Metazoa и реконструкции их ареала расселения от юго-восточного Беломорья до Южного Урала.
Список литературы
Оловянишников В.Г. Верхний докембрий Тимана и полуострова Канин. 1998. Екатеринбург: УрО РАН, 1998. 164 с.
Рифей и венд Европейского Севера СССР. Гецен В.Г., Дедеев В.А. и др. (ред.) 1987. Сыктывкар: Коми ФАН СССР. 124 с.
Гниловская М.Б., Вейс А.Ф. и др. Доэдиакарская фауна Тимана (аннелидоморфы верхнего рифея) // Стратиграфия. Геологическая корреляция. 2000. Т. 8. № 4. С. 11–39.
Kuznetsov N.B., Natapov L.M., et al. Geochronological, geochemical and isotopic study of detrital zircon suites from Late Neoproterozoic clastic strata along the NE margin of the East European Craton: Implications for plate tectonic models // Gondwana Research. 2010. V. 17 (2–3). P. 583–601.
Опаренкова Л.И., Иванов Н.Ф. Легенда Тиманской серии листов Государственной геологической карты Российской Федерации масштаба 1: 200 000 (издание второе) / Ред. В.Г. Колокольцев. 1999. Ухта.
Десяткин В.Д., Колесников А.В. и др. Палеопасцихниды из чернокаменской свиты верхнего венда Среднего Урала (Пермский край) // ДАН. Науки о Земле. 2021. Т. 499 (2). С. 138–143.
Гражданкин Д.В., Маслов А.В. и др. Осадочные системы сылвицкой серии (верхний венд Среднего Урала). Екатеринбург: УрО РАН, 2010. 280 с.
Kolesnikov A. Beltanelliformis konovalovi sp. nov. From the Terminal Neoproterozoic of Central Urals: Taphonomic and Ecological Implications // Frontiers in Earth Science. 2022. 10: 875001.
McIlroy D., Crimes T.P., et al. Fossils and matgrounds from the Neoproterozoic Longmyndian Supergroup, Shropshire, UK // Geological Magazine. 2005. V. 142 (4). P. 441–455.
Menon L.R., McIlroy D., et al. The dynamic influence of microbial mats on sediments: fluid escape and pseudofossil formation in the Ediacaran Longmyndian Supergroup, UK // Journal of the Geological Society. 2016. V. 173. P. 177–185.
Bobrovskiy I., Hope J.M., et al. Molecular fossils from organically preserved Ediacara biota reveal cyanobacterial origin for Beltanelliformis // Nature Ecology & Evoltuion. 2018. V. 2. P. 437–440.
Jensen S. Trace fossils from the Lower Cambrian Mickwitzia sandstone, south-central Sweden // Fossils and Strata. 1997. № 42. 110 p.
Колесников А.В., Гражданкин Д.В. и др. Арумбериеморфные текстуры в верхнем венде Урала // ДАН. 2012. Т. 447 (1). С. 66–72.
Kolesnikov A.V., Danelian T., et al. Arumberiamorph structure in modern microbial mats: implications for Ediacaran palaeobiology // Bull. Soc. géol. Fr. 2017. V. 188 (5). P. 1–10.
Retallack G.J., Broz A.P. Arumberia and other Ediacaran–Cambrian fossils of central Australia // Historical Biology. 2021. V. 33 (10). P. 1964–1988.
McMahon W.J., Davies N.S., et al. Enigma variations: characteristics and likely origin of the problematic surface texture Arumberia, as recognized from an exceptional bedding plane exposure and the global record // Geological Magazine. 2022. V. 159 (1). P. 1–20.
Kolesnikov A.V. Stratigraphic correlation potential of the Ediacaran palaeopascichnids // Estudios Geologicos. 2019. V. 75 (2). e102.
Kolesnikov A., Desiatkin V. Taxonomy and palaeoenvironmental distribution of palaeopascichnids // Geological Magazine. 2022. V. 159. P. 1175–1191.
Гражданкин Д.В., Маслов А.В. Место венда в международной стратиграфической шкале // Геология и геофизика. 2015. Т. 56 (4). С. 703–717.
Sozonov N.G., Bobkov N.I., et al. The ecology of Dickinsonia on tidal flats // Estudios Geologicos. 2019. V. 75 (2). e116.
Дополнительные материалы отсутствуют.
Инструменты
Доклады Российской академии наук. Науки о Земле