

Доклады Российской академии наук. Науки о жизни, 2020, T. 491, № 1, стр. 130-133
ПЕРВАЯ НАХОДКА ЭТРУССКОГО МЕДВЕДЯ (URSUS ETRUSCUS, URSIDAE, CARNIVORA) В КРЫМУ (ПЕЩЕРА ТАВРИДА, РАННИЙ ПЛЕЙСТОЦЕН)
Д. О. Гимранов 1, *, А. В. Лавров 2, Д. Б. Старцев 3, К. К. Тарасенко 2, академик РАН А. В. Лопатин 2, 4
1 Институт экологии растений и животных,
Уральское отделение Российской академии наук
Екатеринбург, Россия
2 Палеонтологический институт имени А.А. Борисяка Российской академии наук
Москва, Россия
3 Крымский федеральный университет
им. В.И. Вернадского
Симферополь, Россия
4 Московский государственный университет
имени М.В. Ломоносова
Москва, Россия
* E-mail: djulfa250@rambler.ru
Поступила в редакцию 26.11.2019
После доработки 26.11.2019
Принята к публикации 26.11.2019
Аннотация
В нижнеплейстоценовых отложениях пещеры Таврида (Крым) найден фрагмент черепа медведя с частично сохранившимися зубными рядами. Присутствие Р1–Р3, строение Р4 и размеры щечных зубов позволяют отнести находку к Ursus etruscus Cuvier, 1823. Новая находка является первой для территории Крыма и России в целом и представляет большой интерес в связи с редкостью этого вида в плейстоцене Восточной Европы.
В 2018 г. в Крыму, в 15 км к востоку от г. Симферополя, была открыта пещера Таврида с многочисленными остатками плейстоценовых животных. Таксономический состав комплекса позвоночных этого местонахождения соответствует позднему виллафранку Западной Европы (зоны MQ18–MQ19) и псекупскому (= одесскому) фаунистическому комплексу России, и имеет возраст около 1.8–1.5 млн. лет [1]. Остатки ископаемых медведей были обнаружены тремя первыми авторами в 2018 г. в приповерхностном слое грунта в небольшом зале пещеры, который называется “Логово гиен”.
В статье описывается фрагмент черепа этрусского медведя Ursus etruscus Cuvier, 1823, хранящийся в Палеонтологическом институте им. А.А. Борисяка РАН (ПИН) в Москве. Образец (экз. ПИН, № 5644/65) был реставрирован из шести крупных и нескольких мелких обломков (рис. 1).
Рис. 1.
Ursus etruscus Cuvier, 1823, экз. ПИН, № 5644/65, фрагмент черепа; вид с вентральной (сверху), левой латеральной (в центре) и правой латеральной (внизу) сторон; пещера Таврида, Крым; нижний плейстоцен.

Экз. ПИН, № 5644/65 представляет собой лицевую часть черепа с обломанными альвеолами резцов и частично разрушенными альвеолами клыков. Каудально он обломан на уровне переднего края глазниц и заднего края носовых костей. Лицевой отдел черепа удлиненной формы, характерной для U. etruscus и U. arctos L., 1758. Носовые кости не сохранились, но судя по величине пространства на их месте, они были короткими и узкими. Носовое отверстие сравнительно небольшое, овальное. Подглазничное отверстие маленькое, лежит над задней частью М1. Передний край глазницы расположен на уровне передней части М2.
В зубных рядах сохранились правый Р4, сильно стертые левые М1 и М2, а также альвеолы правых и левых Р1, Р2, Р3, левого P4 и правых M1–M2. Р4 крупный, метастиль отсутствует, протокон четко выражен. Отчетливый цингулюм окаймляет коронку Р4 по всему периметру зуба, слабее всего развит в антеролабиальной части. Талон М2 относительно длинный. Наличие передних премоляров P1–P3 и строение верхних щечных зубов соответствует признакам U. etruscus [2].
Размеры верхних щечных зубов медведя из пещеры Таврида (экз. ПИН, № 5644/65) и других ископаемых медведей представлены в табл. 1. В связи с тем, что у экз. ПИН, № 5644/65 длина M1 и М2 является реконструируемой (зубы сильно стерты), графики со сравнением размеров построены с использованием ширины М1 и М2 (рис. 2, 3). По средним значениям длины ряда Р4–М2 и ширины М1 и М2 экз. ПИН, № 5644/65 наиболее близок к образцам U. etruscus из местонахождений Вальдарно и Сен-Валье [2].
Таблица 1.
Средние размеры (L – длина, W – ширина) верхних щечных зубов и длина ряда P4–M2 у представителей рода Ursus (N – число образцов, M – среднее значение, в мм)
| Таксон, местонахождение | LP4–M2 | P4 | M1 | M2 | Источник | ||||
|---|---|---|---|---|---|---|---|---|---|
| L | W | L | W | L | W | ||||
| U. etruscus (Вальдарно) | N | 15 | 18 | 18 | 22 | 22 | 20 | 20 | [2] |
| M | 68.4 | 16.4 | 11.6 | 21.3 | 16.5 | 31.5 | 17.7 | ||
| U. etruscus (Сен-Валье) | N | 7 | 7 | 7 | 7 | 7 | 7 | 7 | |
| M | 65.0 | 15.8 | 11.2 | 20.2 | 16.1 | 29.7 | 17.0 | ||
| U. etruscus (Таврида) | N | 1 | 1 | 1 | 1 | 1 | 1 | 1 | Данная статья |
| 70.1 | 15.0 | 10.2 | 24.0* | 16.4* | 34.0* | 17.7* | |||
| U. thibetanus | N | 5 | 10 | 10 | 16 | 17 | 5 | 5 | [2–4] |
| M | 58.2 | 14.1 | 10.1 | 19.0 | 14.9 | 26.5 | 15.1 | ||
| U. arctos suessenbornensis (Дойч-Альтенбург 4) | N | – | 3 | 3 | 6 | 6 | 4 | 4 | [5] |
| M | – | 17.9 | 12.2 | 23.6 | 17.1 | 40.0 | 20.9 | ||
| U. arctos priscus (Торньютон, Таубах) | N | 11 | 30 | 30 | 49 | 49 | 44 | 44 | [2] |
| M | 82.3 | 17.6 | 13.1 | 24.3 | 18.1 | 40.7 | 20.7 | ||
| U. deningeri (Мосбах, Гаргас, Штайнхайм, Пещера Единорога) | N | 6 | 37 | 36 | 47 | 47 | 52 | 53 | [2] |
| M | 89.0 | 18.5 | 13.1 | 25.4 | 18.4 | 40.8 | 20.8 | ||
| U. deningeri (Унтермасфельд) | N | 1 | 1 | 1 | 1 | 1 | 1 | 1 | |
| 84.0 | 18.7 | 13.1 | 27.1 | 20.6 | 40.2 | 20.4 | |||
| U. kudarensis Baryshnikov, 1985 (Кударо 1) | N | 8 | 55 | 58 | 52 | 58 | 33 | 42 | |
| M | 90.7 | 19.3 | 13.6 | 26.1 | 18.9 | 43.3 | 21.9 | ||
| Группа U. savini (Бактон, Кавказ, Урал, Сибирь) | N | 11 | 31 | 24 | 25 | 25 | 22 | 22 | |
| M | 77.6 | 18.0 | 12.5 | 24.8 | 17.4 | 37.5 | 18.7 | ||
| U. spelaeus Rosenmüller, 1794 (Золитен) | N | 19 | 62 | 62 | 62 | 62 | 65 | 65 | |
| M | 92.2 | 20.0 | 13.7 | 28.6 | 19.7 | 44.5 | 22.9 | ||
| U. kanivetz (Vereschagin, 1973)** (= U. ingressus Ra-beder et al., 2004) (Урал) | N | 9 | 19 | 19 | 20 | 20 | 83 | 83 | |
| M | 87.6 | 19.7 | 14.0 | 27.9 | 19.2 | 44.5 | 22.7 | ||
** – Название вида Ursus kanivetz (Vereschagin, 1973) основано на подвидовом наименовании Ursus (Spelaearctos) spelaeus kanivetz Verestchagin, 1973 [6].
Рис. 2.
Сравнение средних значений длины ряда Р4–М2 и ширины M1 у представителей рода Ursus. Значения на осях координат в мм. Обозначения: L – длина; W – ширина; 1 – U. thibetanus, 2 – U. etruscus (Вальдарно), 3 – U. etruscus (Сен-Валье), 4 – U. etruscus (пещера Таврида, показано звездой), 5 – группа форм U. savini, 6 – U. arctos priscus, 7 – U. deningeri (Мосбах), 8 – U. kudarensis (Кударо 1), 9 – U. kanivetz (Урал), 10 – U. spelaeus (Золитен), 11 – U. deningeri (Унтермасфельд).
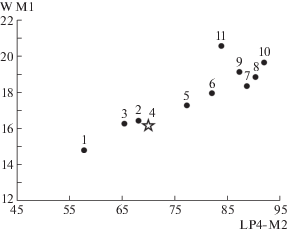
Рис. 3.
Сравнение средних значений длины ряда Р4–М2 и ширины М2 у представителей рода Ursus. Обозначения см. рис. 2.
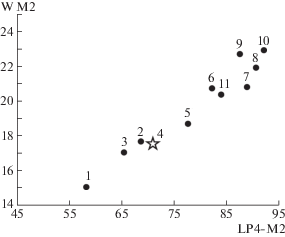
По длине ряда Р4–М2 экземпляр из Крыма заметно мельче, чем позднеплейстоценовые U. arctos priscus Goldfuss, 1818 и медведи группы U. savini Andrews, 1922 [2], но крупнее, чем U. thibetanus Cuvier, 1823. Размеры М1 у экз. ПИН, № 5644/65 находятся в пределах изменчивости U. etruscus, U. arctos suessenbornensis Soergel, 1926 (местонахождение Дойч-Альтенбург 4), U. dolinensis García et Arsuaga, 2001 (местонахождение Атапуэрка TD 4), U. arctos priscus и медведей из группы U. savini [2, 5, 7]. Длина и ширина М2 у экз. ПИН, № 5644/65 лежат в пределах изменчивости U. etruscus, U. arctos и медведей из группы U. savini [2].
Медведь из пещеры Таврида отличается от гималайского медведя U. thibetanus более крупными зубами и строением Р4 (хорошо развитый паракон, значительно превосходящий по величине метакон, и относительно крупный протокон). Экз. ПИН, № 5644/65 не имеет выраженного антеролингвального угла коронки М1, который характерен для U. thibetanus. Лицевая часть черепа экз. ПИН, № 5644/65 удлинена сильнее, чем у гималайского медведя.
Описываемый экземпляр отличается от медведей группы U. savini наличием альвеол Р1–Р2, которые крайне редко встречаются у последних [8], отсутствием дополнительных бугорков на Р4 [2] и меньшими размерами зубов (табл. 1, рис. 2, 3).
От плейстоценовых представителей U. arctos описываемый экземпляр отличается намного меньшими размерами зубов (табл. 1, рис. 2, 3) и наличием P1–P2. По строению паракона и метакона Р4 экз. ПИН, № 5644/65 сходен с морфотипом В1, выделенным у современного бурого медведя [9]; однако данный морфотип у U. arctos крайне редок, его частота встречаемости – 0.1% [9].
Нужно отметить, что у позднеплейстоценовых бурых медведей Р4 усложнен дополнительными бугорками [9, 10]. Кроме того, медведь из пещеры Таврида отличается от U. arctos укороченными носовыми костями.
Различия с U. deningeri Reichenau, 1904 состоят в более мелких размерах, наличии P1–P3 (почти всегда отсутствующих у всех медведей спелеоидной линии [8]), строении Р4 (у медведя Денингера на P4 почти всегда присутствует метастиль), короткой ростральной части черепа, меньших размерах носового отверстия и более переднем положении подглазничного отверстия.
Этрусский медведь был широко распространен в раннем плейстоцене и входил в состав многих фаун позднего виллафранка [2]. U. etruscus описан из двух местонахождений Украины, а также с территории Кавказа [2]. Предполагается, что распространение этрусского медведя было ограничено южными широтами Палеарктики. Первое появление U. etruscus датируется средним виллафранком (нижняя часть MN17). Самые поздние находки вида относятся к позднему виллафранку – MQ18–MQ20 [2, 11, 12]. В настоящее время нет однозначного мнения о таксономическом положении большинства находок медведей спелеоидной и арктоидной линий раннего плейстоцена Европы (поздний виллафранк, MQ19–MQ20) [2, 12–14]. Некоторые исследователи рассматривают ранних спелеоидных медведей, таких как U. dolinensis, в качестве переходных форм от U. etruscus к U. deningeri [7]. Также в раннем и среднем плейстоцене на территории Европы обитали U. thibetanus и U. savini.
Большинство находок U. etruscus сделаны на территории Западной и Юго-Восточной Европы – в Испании (Эль-Ринкон, Ла-Пуэбла-де-Вальверде, Вента-Мисена), Франции (Сен-Валье, Шайак, Сенез, Сесаге), Нидерландах (Тегелен), Италии (Оливола, Вальдарно, Кростоло-Модолена, Пьетрафитта, Колле-Курти, Монте-Арджентарио, Пирро-Норд), Германии (Эрпфинген, Шернфельд), Румынии (Граунчанулуй), Болгарии (Варшетс), Греции (Вассилоуди 1, Аполлония 1, Дафнеро 1, Тсиотра-Врисси), а также на Украине (Горишняя Выгнанка, Котловина 1). В Азии остатки U. etruscus описаны из Израиля (Убейдия), Грузии (Дманиси), Азербайджана (Палан-Тюкан), Таджикистана (Куруксай, Оби-Гарм, Тутак), Китая (Нихэвань, Чжоукоудянь 18, пещера Цзиньюань) [14]. Одна находка известна из Северной Африки – из Марокко (Ahl al Oughlam) [2].
Этрусский медведь рассматривается как всеядная форма, по характеру питания и общим размерам сходная с некрупными современными бурыми медведями U. arctos [15].
Описанная находка является первым свидетельством обитания этрусского медведя на территории Крыма и России в целом и, таким образом, дополняет представления о распространении этого вида в раннем плейстоцене на территории Восточной Европы.
БЛАГОДАРНОСТИ
Авторы благодарят сотрудников Крымского федерального университета им. В.И. Вернадского, помогавших в организации и проведении полевых работ в пещере Таврида. Авторы особенно признательны Я. Вагнеру (Национальный музей, Прага, Чехия) за ценные советы и замечания в процессе подготовке публикации.
ИСТОЧНИКИ ФИНАНСИРОВАНИЯ
Работа выполнена в рамках государственных заданий Института экологии растений и животных УрО РАН и ПИН, при частичной поддержке гранта Президента Российской Федерации для молодых российских ученых (МК-1130.2019.4), а также проекта РФФИ № 19-05-00982, Программы фундаментальных исследований Президиума РАН “Эволюция органического мира. Роль и влияние планетарных процессов” и проекта программы УрО РАН № 18-4-4-3.
Список литературы
Лопатин А.В., Вислобокова И.А., Лавров А.В. и др. // ДАН. 2019. Т. 485. № 3. С. 40–43.
Барышников Г.Ф. Семейство медвежьих (Carnivora, Ursidae). СПб.: Наука, 2007. 542 с.
Baryshnikov G.F. // Proceeding of the Zoological Institute RAS, 2010. V. 314. № 1. P. 67–79.
Baryshnikov G.F., Zakharov D.S. // Proceedings of the Zoological Institute RAS. 2013. V. 317. № 1. P. 3–10.
Rabeder G., Pacher M., Withalm G. // Mitt. Komm. Quartärforsch. Österr. Akad. Wiss. 2010. Bd 17. P. 1–135.
Верещагин Н.К. // Зоол. журн. 1973. Т. 52. № 6. С. 920–930.
García N., Arsuaga J.L. // Earth and Planetary Sci. 2001. V. 332. P. 717–725.
Pacher M. // Fossil Imprint. 2017. V. 73. № 3–4. P. 482–494.
Гимранов Д.О. // Зоол. журн. 2018. Т. 97. № 2. С. 205–223.
Marciszak A., Schouwenburg C., Lipecki G. et al. // Quater. Int. 2019. V. 534. P. 158–170.
Mazza P., Rustioni M. // Palaeontogr. Ital. 1992. V. 79. P. 101–146.
Wagner J., Cermak S. // Bull. Geosci. 2012. V. 87. № 3. P. 461–496.
Agadzhanyan A.K., Vislobokova I.A., Shunkov M.V., Ulyanov V.A. // Fossil Imprint. 2017. V. 73. P. 93–114.
Jiangzuo Q., Liu J., Wang Y. et al. // Quatern. Sci. 2017. V. 37. № 4. P. 828–837.
Medin T., Martínez-Navarro B., Rivals F. et al. // Quatern. Int. 2017. V. 431. Pt B. P. 20–41.
Дополнительные материалы отсутствуют.
Инструменты
Доклады Российской академии наук. Науки о жизни