

Доклады Российской академии наук. Науки о жизни, 2020, T. 491, № 1, стр. 134-136
ИЗОТОПНЫЙ СОСТАВ АЗОТА И СТЕХИОМЕТРИЧЕСКИЕ СООТНОШЕНИЯ ЭЛЕМЕНТОВ В БИОМАССЕ SPIROGYRA В ОЗЕРЕ БАЙКАЛ
Член-корреспондент РАН М. И. Гладышев 1, 2, *, Л. А. Глущенко 2, Е. С. Кравчук 1, О. В. Анищенко 1, А. А. Колмакова 1, М. Ю. Колобов 3, В. А. Разгоняева 3, Н. Н. Сущик 1, 2
1 Институт биофизики Сибирского отделения Российской академии наук, Федеральный исследовательский центр “Красноярский научный центр СО РАН”
Красноярск, Россия
2 Сибирский федеральный университет
Красноярск, Россия
3 Московский государственный университет
имени М.В. Ломоносова
Москва, Россия
* E-mail: glad@ibp.ru
Поступила в редакцию 16.12.2019
После доработки 16.12.2019
Принята к публикации 16.12.2019
Аннотация
На основе анализа изотопного состава азота была впервые подтверждена гипотеза о разных источниках элементов минерального питания, потребляемых двумя группами водорослей литорали оз. Байкал. Сильно прикрепленные нитчатые водоросли рода Ulothrix, развивающиеся в прибойной зоне на глубине 0.5 м и слабо прикрепленные нитчатые водоросли рода Spirogyra, обитающие на большей глубине 3.0–5.0 м в зоне с пониженной турбулентностью, получают элементы минерального питания предположительно из поверхностного стока и грунтовых вод соответственно. Кроме того, стехиометрические соотношения C:N и N:P в биомассе водорослей свидетельствовали о том, что рост Spirogyra в оз. Байкал вероятнее всего был лимитирован азотом, тогда как рост Ulothrix был лимитирован фосфором.
В последние десятилетия глобальный характер приобрело массовое развитие водорослей в прибрежье озер и морей, получившее название “зеленых приливов” [1]. “Зеленые приливы”, вызываемые водорослями рода Spirogyra, возникли даже в олиготрофном оз. Байкал, причиняя ему значительный экологический ущерб [2]. Очевидно, что основной причиной “зеленых приливов” является эвтрофирование, то есть, увеличение потоков элементов минерального питания в водоемы, способствующее росту биомассы водорослей [1]. Тем не менее, для массового развития водорослей, как планктонных, так и бентосных, важна не только величина, но и локализация потока элементов, поступающих либо непосредственно в пелагиаль (эстуарии крупных рек или внутренняя биогенная нагрузка из анаэробного гиполимниона), либо в прибойную зону (поверхностный сток малых рек и ручьев), либо в приглубую часть литорали (разгрузка грунтовых вод) [3]. Например, для оз. Байкал были получены данные об усиливающемся антропогенном загрязнении (эвтрофировании) грунтовых вод в прибрежье и утверждалось, что именно эвтрофирование грунтовых вод и является причиной развития Spirogyra [2, 4]. Однако, в этих работах [2, 4] не объяснялось, почему именно Spirogyra, а не другие байкальские бентосные нитчатые водоросли (Cladophora, Ulothrix и др.) получили преимущества от этого эвтрофирования. Нами была выдвинута гипотеза о различных источниках элементов минерального питания для нитчатых водорослей прибойной зоны, способных сильно прикрепляться к твердому субстрату (Cladophora, Ulothrix и др.) и слабо прикрепленной Spirogyra, обитающей в основном на больших глубинах, в зоне с низкой турбулентностью (0 < Re < 7500) [3].
Для проверки гипотезы о различных источниках элементов минерального питания для сильно прикрепленных нитчатых водорослей прибойной зоны и слабо прикрепленной Spirogyra, в 2019 г. были проведены экспедиционные исследования на оз. Байкал. В период максимального развития Spirogyra, в августе–сентябре, пробы отбирались на о. Ольхон, рядом с поселком Хужир (53°11′37.73″ С 107°19′48.45″ В и 53°11′44.13″ С 107°19′50.47″ В; точка 3: 53°12′04.54″ С 107°20′22.78″ В) и заливе Загли (53°02′14.69″ С 106°57′29.07″ В и 53°02′06.41″ С 106°57′01.54″ В), а также рядом с поселками Листвянка (51°50′34.10″ С 104°52′39.97″ В), Большое Голоустное (52°01′34.03″ С 105°23′49.80″ В), Култук (51°43′23.93″ С 103°43′00.95′′ В) и Слюдянка (51°39′47.85″ С 103°43′25.41″ В).
Пробы Spirogyra брались водолазами на глубине 3.0–5.0 м, в отличие от других исследований, в которых ограничивались глубинами 0.5–1.5 м [2]. Напротив глубинных точек отбора Spirogyra, в прибойной зоне на глубине менее 0.5 м также брались пробы эпилитных нитчатых водорослей, представленных в основном Ulothrix. Нити Spirogyra и Ulothrix, очищенные от детрита и песка под бинокулярным микроскопом, замораживались и хранились при температуре –18°C до последующих анализов в лаборатории. Для анализов по ранее описанным методикам использовался элементный анализатор, эмиссионный спектрометр с индуктивно-связанной плазмой и изотопный масс-спектрометр [5–7].
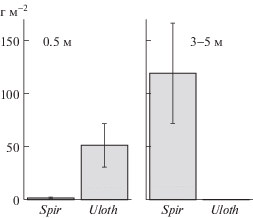
На глубинах 0.5 м доминировал Ulothrix, биомасса которого значительно превышала таковую Spirogyra и составляла в среднем 74.5 ± 11.8% от общей биомассы всех макро- и микроводорослей (рис. 1). В то же время на глубинах 3.0–5.0 м доминировала Spirogyra, биомасса которой составляла в среднем 93.6 ± 2.2% от общей биомассы всех водорослей (рис. 1).
Рис. 1.
Средняя биомасса Spirogyra и Ulothrix на глубинах 0.5 и 3.0–5.0 м в оз. Байкал в августе–сентябре 2019. Отрезками обозначены стандартные ошибки.
Стехиометрическое соотношение углерода и азота (C:N) в биомассе Spirogyra в среднем было достоверно выше, чем таковое Ulothrix (критерий Стьюдента t = 2.61, p < 0.05, число степеней свободы df = 17; рис. 2).
Рис. 2.
Среднее соотношение стабильных изотопов азота (δ15N, ‰) и стехиометрические соотношения углерода и азота (C:N, моль : моль) и углерода и фосфора (N:P, моль : моль) в биомассе Spirogyra на глубине 3.0–5.0 м и в биомассе Ulothrix на глубине 0.5 м в оз. Байкал в августе–сентябре 2019. Отрезками обозначены стандартные ошибки.
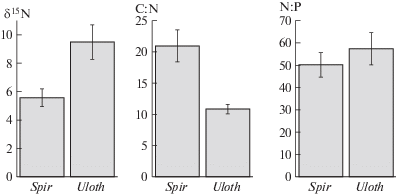
Напротив, стехиометрическое соотношение азота и фосфора (N:P) в биомассе Spirogyra было в среднем ниже, чем таковое Ulothrix, хотя эта тенденция и не была статистически достоверной (t = 0.76, p > 0.05, df = 17; рис. 2).
Обнаруженные стехиометрические соотношения элементов в биомассе свидетельствуют о том, что рост Spirogyra, в отличие от Ulothrix, был лимитирован азотом, а не фосфором [8]. Полученные результаты согласуются с литературными данными о том, что рост Spirogyra, в отличие от других макроводорослей, лимитируется азотом, а не фосфором [3]. Однако, следует отметить, что стехиометрические соотношения C:N и N:P являются лишь косвенными признаками лимитирования. Для прямого определения лимитирующего элемента необходимы специальные эксперименты [9].
Соотношение стабильных изотопов азота δ15N в биомассе Spirogyra в среднем было достоверно ниже, чем таковое для Ulothrix (t = 3.16, p < 0.01, df = 19; рис. 2). Данные различия свидетельствуют о том, что Spirogyra на глубине 3.0–5.0 м и Ulothrix на глубине 0.5 м получают элементы минерального питания из разных источников. Вероятно, Spirogyra получала азот из просачивающихся грунтовых вод, тогда как Ulothrix получал этот и другие элементы из поверхностного стока.
Таким образом, получены свидетельства о том, что рост Spirogyra в оз. Байкал лимитируется азотом, а не фосфором. Получено подтверждение гипотезы о различных источниках элементов минерального питания для сильно прикрепленных нитчатых водорослей прибойной зоны (Ulothrix) и слабо прикрепленной Spirogyra, развивающейся на глубинах с пониженной турбулентностью. Для более точной идентификации источников элементов минерального питания, вызывающих «зеленые приливы» Spirogyra в оз. Байкал необходимо провести комплексные геологические (гидрогеологические) и географические исследования, а также биофизические эксперименты и модельные расчеты с целью выработки научно обоснованных рекомендаций по охране уникального водоема – озера Байкал.
ИСТОЧНИКИ ФИНАНСИРОВАНИЯ
Экспедиция была поддержана группой компаний En+ Group и Экологическим фондом Сибирского федерального университета. Работа поддержана Государственным заданием в рамках программы фундаментальных исследований РФ, тема № 51.1.1.
Список литературы
Smetacek V., Zingone A. // Nature 2013. V. 504. P. 84–88.
Timoshkin O.A., Moore M.V., Kulikova N.N., Tomberg I.V., Malnik V.V., et al. // J. Great Lakes Res. 2018. V. 44. P. 230–244.
Gladyshev M.I., Gubelit Y.I. // Contemp. Probl. Ecol. 2019. V. 12. P. 109–125.
Kravtsova L.S., Izhboldina L.A., Khanaev I.V., Pomazkina G.V., Rodionova E.V., et al. // J. Great Lakes Res. 2014. V. 40. P. 441–448.
Gladyshev M.I., Kolmakova O.V., Tolomeev A.P., Anishchenko O.V., Makhutova O.N., et al. // Limnol. Oceanogr. 2015. V. 60. P. 1314–1331.
Иванова Е.А., Анищенко О.В., Зуев И.В., Авра-мов А.П. // J. Sib. Fed. Univ. Biol. 2015. V. 3. P. 347–361.
Anishchenko O.V., Sushchik N.N., Makhutova O.N., Kalachova G.S., Gribovskaya I.V., et al. // Food Chem. Toxicol. 2017. V. 101. P. 8–14.
Gubelit Y.I., Makhutova O.N., Sushchik N.N., Kolmakova A.A., Kalachova G.S., Gladyshev M.I. // J. Appl. Phycol. 2015. V. 27. P. 375–386.
Дегерменджи А.Г., Печуркин Н.С., Шкидченко А.Н. Аутостабилизация факторов, контролирующих рост в биологических системах. Новосибирск: Наука, 1979. 141 с.
Дополнительные материалы отсутствуют.
Инструменты
Доклады Российской академии наук. Науки о жизни