

Доклады Российской академии наук. Науки о жизни, 2020, T. 491, № 1, стр. 180-183
НОВЫЙ РОД BRACHYERICINAE (ERINACEIDAE, ERINACEOMORPHA) ИЗ НИЖНЕГО МИОЦЕНА МОНГОЛИИ
Академик РАН А. В. Лопатин 1, *
1 Палеонтологический институт им. А.А. Борисяка Российской академии наук
Москва, Россия
* E-mail: alopat@paleo.ru
Поступила в редакцию 10.02.2020
После доработки 10.02.2020
Принята к публикации 10.02.2020
Аннотация
Из нижнемиоценовой свиты лу местонахождения Тацин-Гол (Долина Озер, Монголия) описан Ereberix erebericulus gen. et sp. nov., самый мелкий представитель азиатских Brachyericinae (Erinaceidae, Erinaceomorpha), своеобразной вымершей группы короткомордых ежей с хищническими адаптациями. Описание новой формы дополняет представления о таксономическом и морфологическом разнообразии Brachyericinae Азии.
Короткомордые ежи (Brachyericinae) – вымершее подсемейство ежовых с хищническими адаптациями. Эта группа характеризуется укороченным лицевым отделом черепа, специализированным строением слуховой области и сокращенным числом щечных зубов (P2/1–2M2/2). В составе подсемейства установлено пять родов: Brachyerix Matthew, 1933 и Metechinus Matthew, 1929 из миоцена Северной Америки, Exallerix McKenna et Holton, 1967 (= Metexallerix Qiu et Gu, 1988) из олигоцена Монголии и Китая и базального миоцена Казахстана, Synexallerix Lopatin et Zazhigin, 2003 из раннего миоцена Китая, Казахстана и Германии, Postexallerix Lopatin et Zazhigin, 2003 из раннего и среднего миоцена Монголии [1–13]. Находки азиатских брахиэрицин очень редки [3, 6, 8–10, 12, 13].
Азиатские Brachyericinae отличаются от североамериканских короткомордых ежей комплексом признаков, как примитивных, так и специализированных [10]. Самые поздние и наиболее специализированные азиатские Brachyericinae рода Postexallerix характеризуются очень узким и длинным тригонидом М1 и сильно редуцированным М2 [10]. Степень удлинения тригонида и редукции талонида М1 Brachyericinae количественно описывается индексами (%), отражающими отношение длины тригонида к общей длине зуба (LTR/L), длины зуба к ширине тригонида (L/WTR), длины тригонида к его ширине (LTR/WTR), длины талонида к длине тригонида (LTL/LTR) [10], а также ширины талонида к ширине тригонида (WTL/WTR).
В настоящей статье описан фрагмент нижней челюсти нового представителя брахиэрицин из нижнего миоцена Монголии. Образец найден в местонахождении Тацин-Гол (сборы Д. Бадамгарав и Б. Баярмаа, Институт палеонтологии Академии наук Монголии, 2012 г.) в Долине Озер, в части разреза (на правом берегу р. Тацин-Гол), относимой к свите лу и биозоне D [14]. Материал хранится в коллекции Палеонтологического института им. А.А. Борисяка РАН (ПИН) в Москве.
Терминология и методика промеров зубов по [5, 7, 10].
Фотографии сделаны на сканирующем электронном микроскопе TESCAN VEGA-II XMU в ПИН.
Семейство Erinaceidae Fischer, 1817
Подсемейство Brachyericinae Butler, 1948
Род Ereberix Lopatin, gen. nov.
Название рода от Erebus, др.-греч. божество вечного мрака, и erix, лат. еж.
Типовой вид – Ereberix erebericulus sp. nov.
Диагноз. Brachyericinae мелких размеров. М1 с узким и длинным тригонидом (отношение длины тригонида к его ширине около 180%, отношение общей длины зуба к ширине тригонида около 270%) и сильно удлиненным низким паралофидом; метаконид умеренно редуцирован; талонид заметно шире тригонида (отношение ширины талонида к ширине тригонида более 110%), по длине вдвое уступает тригониду; талонидный бассейн глубокий, открытый лингвально; энтоконид высокий, вытянутый продольно; задняя ветвь гипоконида соединяется с энтоконидом; эмалевая скульптура не выражена. Нижний массетерный гребень горизонтальной ветви нижней челюсти отсутствует.
Видовой состав. E. erebericulus sp. nov., нижний миоцен Монголии.
Сравнение. Отличается от всех родов Brachyericinae, кроме Metechinus, отсутствием нижнего массетерного гребня на горизонтальной ветви нижней челюсти; от всех родов, исключая Postexallerix, отличается значительно более узким и длинным тригонидом М1 (отношение длины тригонида М1 к его ширине у Metechinus – 90–123%, у Brachyerix – 105–147%, у Synexallerix – 119–151%, у Exallerix – 126–143%, у Postexallerix – 194–200%; отношение общей длины М1 к ширине тригонида в той же последовательности родов – 166–200%, 179–264%, 196–242%, 192–204%, 283–284%) [10, 11]. Помимо этого, в строении M1 новый род отличается от Synexallerix более коротким талонидом, лингвально открытым талонидным бассейном и редукцией эмалевой скульптуры, от Postexallerix – низким паралофидом, менее редуцированным метаконидом, более длинным и широким талонидом, высоким энтоконидом, более глубоким и закрытым сзади талонидным бассейном.
Замечания. Для большинства брахиэрицин характерно наличие хорошо развитого гребня на лабиальной стороне горизонтальной ветви нижнечелюстной кости, который интерпретируется как нижний массетерный гребень [3, 6, 9, 10], служащий для прикрепления вынесенной далеко вперед гипертрофированной передней порции медиальной жевательной мышцы (m. masseter medialis) с функцией очень мощного аддуктора [3, 10]. У Exallerix этот гребень особенно мощный и высокий [3, 6, 8, 10], у Synexallerix развит несколько слабее [9], у Brachyerix и Postexallerix – еще более слабый, но все же отчетливый [3, 4, 9–11], у Metechinus полностью отсутствует [3–5], как и у Ereberix gen. nov.
Ereberix erebericulus Lopatin, sp. nov.
Название вида от Erebus, др.-греч. божество вечного мрака, и ericulus, лат. маленький еж.
Голотип – ПИН, № 5672/1, фрагмент левой нижнечелюстной кости с M1 и альвеолами M2; Монголия, Долина Озер, Тацин-Гол (правый берег); нижний миоцен, свита лу.
Описание (рис. 1). Горизонтальная ветвь нижнечелюстной кости низкая. Основание переднего края венечного отростка отклонено от горизонтальной ветви под углом около 110°. Позади альвеол М2 на медиальном выступе кости в основании венечного отростка находится резко обозначенная полукруглая площадка для прикрепления крупной доли височной мышцы (рис. 1в). Массетерная впадина на лабиальной стороне венечного отростка умеренно глубокая, четко очерченная спереди. Нижнего массетерного гребня на лабиальной стороне горизонтальной ветви нет; однако от массетерной впадины до четко выраженного углубления, лежащего под серединой М1, тянется малозаметная выровненная площадка, вероятно, служившая для крепления передней порции медиальной жевательной мышцы. Нижнечелюстное отверстие крупное, расположено чуть вентральнее уровня альвеолярного края горизонтальной ветви.
Рис. 1.
Ereberix erebericulus Lopatin, gen. et sp. nov., голотип ПИН, № 5672/1, фрагмент левой нижнечелюстной кости с M1 и альвеолами M2: а – с лабиальной стороны, б – с окклюзиальной стороны, в – то же, с лабиальным наклоном, г – с лингвальной стороны; Монголия, Тацин-Гол (правый берег); нижний миоцен, свита лу.
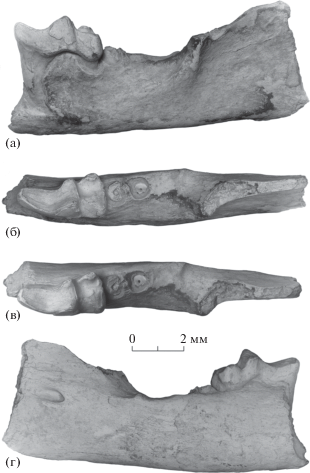
M1 длинный и узкий, с сильно удлиненным тригонидом и коротким, но широким талонидом. Паралофид (паракристид) мощный, слегка выгнутый лабиально, с утолщенным лабиальным эмалевым слоем (рис. 1б, 1в). Тригонидный бассейн полностью выровнен в результате стирания и объединен с паралофидом. В стертом состоянии параконид и метаконид почти равной высоты, протоконид немного выше. Задняя стенка тригонида субвертикальная. Косой кристид присоединен к середине основания задней стенки протоконида и восходит на протоконид. Талонидный бассейн глубокий, открытый лингвально (энтокристид не достигает стенки тригонида, гребня метастилида нет), но замкнутый сзади. Гипоконид умеренно выдается постеролабиально. Энтоконид высокий, овальной формы, в передней части вытянут продольно, образуя короткий, резко понижающийся энтокристид. Талонидной вырезки нет. Задняя ветвь гипоконида длинная, присоединена к задней стороне энтоконида. Постцингулида и эктоцингулида нет, какие-либо скульптурные элементы или морщинистость эмали на лабиальной стороне зуба отсутствуют (часть поверхности эмали эродирована), лишь в основании протоконида идентифицируется легкое продольное утолщение, более заметное в наиболее задней части, непосредственно перед гипофлексидом.
Судя по альвеолам, М2 почти в два раза уступал М1 по длине.
Размеры голотипа в мм. М1: общая длина (L) – 3.35; длина тригонида (LTR) – 2.25; лабиальная длина талонида (LTL) – 1.05; лингвальная длина талонида – 0.85; ширина тригонида (WTR) – 1.25; ширина талонида (WTL) – 1.4; индексы, %: LTR/L – 67; L/WTR – 268; LTR/WTR – 180; LTL/LTR – 47; WTL/WTR – 112.
Длина М2 по альвеолам – 1.7.
Высота зубной кости под М2 с лингвальной стороны – 2.8.
Замечания. Ereberix erebericulus gen. et sp. nov. по степени удлиненности тригонида М1 наиболее близок Postexallerix securis Lopatin et Zazhigin, 2003, также описанному из нижнего миоцена Монголии (местонахождение Улан-Тологой, свита лу) [10]. Помимо отличительных признаков, упомянутых выше при сравнении родов, E. erebericulus gen. et sp. nov. отличается от P. securis существенно более мелкими размерами (у P. securis длина M1 – 4.55–5.1, ширина тригонида – 1.6–1.8, ширина талонида – 1.65–1.95, длина М2 по альвеолам – 2.2, высота зубной кости под М2 – 3.3) и некоторыми дополнительными индексами М1 (у P. securis LTR/L – 69–70%; LTL/LTR – 42–46%; WTL/WTR – 103–108%) [10]. То же относится к P. mustelidens Lopatin et Zazhigin, 2003 из среднего миоцена Монголии (М1: LTL – 1.5; лингвальная длина талонида – 1.3; WTR – 1.7; WTL – 1.75; WTL/WTR – 103%).
Описанный Р. Циглером с соавторами [13, с. 74, рис. 6.3 ] по фрагменту нижнечелюстной кости с P4 и сильно разрушенным M1 мелкий представитель брахиэрицин (“Exallerix sp.”) из нижнемиоценовых слоев (биозона D) местонахождения Унхэлцэг [14, табл. 8 ] по размерам М1 (WTR – 1.45) близок E. erebericulus gen. et sp. nov. (WTR – 1.25). Однако, судя по описанию и изображению [13], у формы из Унхэлцэга на M1 имеется хорошо развитая лабиальная скульптура в виде эктоцингулида, состоящего из продольного ряда эмалевых бугорков, что четко отличает ее от Ereberix gen. nov. и Postexallerix.
Бугорчатый скульптурный рельеф эмали лабиальной поверхности нижних зубов характерен для азиатских брахиэрицин [10], но отсутствует у представителей обоих североамериканских родов [5]. Однако степень развития скульптуры у азиатских видов разная. У Exallerix pustulatus Ziegler, Dahlmann et Storch, 2007 (верхний олигоцен Монголии) на P4–M2 имеется четыре полных продольных ряда тесно расположенных эмалевых бугорков и еще несколько их неупорядоченных скоплений (фактически, бугорками покрыта вся лабиальная сторона зубов, исключая вершины параконида, протоконида и гипоконида) [13], у E. hsandagolensis McKenna et Holton, 1967 (нижний олигоцен Монголии) – два продольных ряда эмалевых бугорков в основании коронки [3], у E. efialtes Lopatin, 1996 (базальный миоцен Казахстана) – один ряд [8, 10]. У E. manahan Lopatin et Zazhigin, 2003 (нижний олигоцен Монголии) и Synexallerix otus Lopatin et Zazhigin, 2003 (нижний миоцен Восточного Казахстана) эмалевые бугорки объединены в эктоцингулид, но у первого он сравнительно слабый, а у второго – хорошо развитый [10]. У S. junggarensis (Bi, 1999) из нижнего миоцена Китая эмалевые бугорки в единственном продольном ряду мелкие, редко расставленные [9], у обоих видов Postexallerix бугорки в основании коронки единичные, очень маленькие, эмаль местами морщинистая [10], а у Exallerix gaolanshanensis (Qiu et Gu, 1988) из верхнего олигоцена Китая [6] и Ereberix erebericulus gen. et sp. nov. эмалевая скульптура полностью редуцирована. Подобная лабиальная эмалевая скульптура на нижних зубах имеется у Scymnerix tartareus Lopatin, 2003 из нижнего олигоцена Монголии (три продольных ряда довольно крупных эмалевых бугорков на P4, два ряда на M1 и один ряд на M2); однако Scymnerix принадлежит к подсемейству Erinaceinae [12, 15].
Материал. Голотип.
Специализированное строение зубов (очень крупный передний резец, увеличенный М1 с высоким, узким и длинным лезвиеподобным паралофидом, редукция остальных зубов) и нижней челюсти (укороченная горизонтальная ветвь, мощно развитый и вынесенный далеко вперед нижний массетерный гребень и др.) азиатских короткомордых ежей, не имеющее аналогов среди современных и ископаемых Erinaceidae, интерпретируется как приспособление к преимущественно хищническому типу питания, ориентированному на мелких позвоночных [10]. Вероятно, с хищническим образом жизни связаны и сравнительно крупные для Erinaceidae размеры большинства азиатских брахиэрицин. Однако Ereberix erebericulus gen. et sp. nov. демонстрирует небольшие размеры, по длине ряда нижних моляров и высоте горизонтальной ветви нижнечелюстной кости сравнимые с Brachyerix incertis (Matthew, 1924) [5]. Эреберикс был самым мелким из азиатских Brachyericinae; вероятно, он в основном охотился на крупных наземных беспозвоночных – насекомых, паукообразных, брюхоногих моллюсков.
Описание нового рода дополняет представления о таксономическом и морфологическом разнообразии Brachyericinae Азии.
БЛАГОДАРНОСТИ
Автор благодарен покойной Д. Бадамгарав за переданный материал и бесценный опыт сотрудничества в экспедициях 2011–2013 гг. в Монголии, П.Ю. Пархаеву и Р.А. Ракитову (ПИН) – за техническую помощь в подготовке фотографий.
Список литературы
Matthew W.G. // Univ. Calif. Publ. Depart. Geol. Sci. 1929. V. 18. № 4. P. 92–102.
Matthew W.G., Mook C.C. // Amer. Museum Novit. 1933. № 601. P. 1–7.
McKenna M.C., Holton C.P. // Amer. Museum Novit. 1967. № 2311. P. 1–11.
Rich T.H.V., Rich P.V. // Amer. Museum Novit. 1971. № 2477. P. 1–59.
Rich T.H.V. // Bull. Amer. Museum Natur. History. 1981. № 171. P. 1–116.
Qiu Z.X., Gu Z.G. // Vertebr. PalAsiat. 1988. V. 26. № 3. P. 198–213.
Gould G.C. // Amer. Museum Novit. 1995. № 3131. P. 1–45.
Лопатин А.В. // Стратигр. Геол. корреляция. 1996. Т. 4. № 2. С. 65–79.
Bi S.-D. // Vertebr. PalAsiat. 1999. V. 37. № 2. P. 140–155.
Лопатин А.В., Зажигин В.С. // Палеонтол. журн. 2003. № 1. С. 64–77.
Martin L.D., Lim J.-D. // Mammal. Biol. 2004. V. 69. № 3. P. 202–209.
Lopatin A.V. // Paleontological Journal. 2006. V. 40. Suppl. 3. P. S205–S405.
Ziegler R., Dahlmann T., Storch G. // Ann. Naturhist. Mus. Wien. Ser. A. 2007. Bd 108. S. 53–164.
Daxner-Höck G., Badamgarav D., Barsbold R. et al. // Palaeobiodiv. Palaeoenv. 2017. V. 97. № 1. P. 111–218.
Лопатин А.В. // Палеонтол. журн. 2003. № 6. С. 94–104.
Дополнительные материалы отсутствуют.
Инструменты
Доклады Российской академии наук. Науки о жизни