

Доклады Российской академии наук. Науки о жизни, 2020, T. 492, № 1, стр. 297-299
АНТАГОНИСТЫ D1 И D2 РЕЦЕПТОРОВ ДОПАМИНА ВЫЯВЛЯЮТ РАЗНЫЕ МЕХАНИЗМЫ НЕЙРОПРОТЕКТОРНОГО ДЕЙСТВИЯ В ВЕСТИБУЛЯРНОМ АППАРАТЕ ЛЯГУШКИ
И. В. Рыжова 1, Т. В. Тобиас 1, *, академик РАН А. Д. Ноздрачев 1
1 Институт физиологии им. И.П. Павлова Российской академии наук
Санкт-Петербург, Россия
* E-mail: tvtobias@yandex.ru
Поступила в редакцию 06.02.2020
После доработки 20.02.2020
Принята к публикации 20.02.2020
Аннотация
Высокая функциональная пластичность глутаматэргического синапса вестибулярного эпителия поддерживается тонким балансом возбуждающих и тормозных взаимодействий. Одним из пептидов, колокализованных с ацетилхолином (АХ) в эфферентных волокнах, является допамин. В работе на изолированном вестибулярном аппарате лягушки в условиях внешней перфузии синаптической зоны электрофизиологически исследовали модулирующее воздействие антагонистов допамина на фоновую активность сенсорных волокон полукружных каналов. Установлено, что антагонист D1 рецепторов допамина SCH-23390 существенно понижал уровень фоновой активности в высоких концентрациях. Антагонист D2 этиклопрайд в зависимости от концентрации вызывал позитивно-негативные сдвиги фоновой активности сенсорных волокон, что подтверждает гипотезу о том, что допамин, тонически выделяющийся из эфферентных окончаний, оказывает нейропротекторное ингибирующее влияние на активность афферентного глутаматэргического синапса в вестибулярном эпителии.
Функциональная пластичность глутаматэргического синапса между волосковой клеткой и сенсорным волокном поддерживается тонким балансом возбуждающих и тормозных взаимодействий афферентной и эфферентной систем. Известно, что фоновая и вызванная активность афферентных волокон формируется за счет различных механизмов [1]. Из базальной мембраны волосковых клеток происходит тоническое выделение глутамата из базальной мембраны, высокая концентрация которого может вызывать эксайтотоксичность. В связи с этим предcтавляется исключительно важным исследовать нейропротекторную роль допамина, колокализованного с АХ в эфферентных волокнах вестибулярного эпителия.
Работа выполнена на 39 изолированных вестибулярных аппаратах лягушки в условиях внешней перфузии синаптической области и отведения активности нервных волокон, контактирующих с задним полукружным каналом. Регистрация импульсной активности сенсорных волокон, контактирующих с рецепторным эпителием полукружного канала, осуществлялась посредством засасывающего стеклянного микроэлектрода. Спайки подавались на усилитель A-M Systems Inc 3000, преобразовывались в стандартные прямоугольные импульсы (2 мс) и регистрировались on-line в течение всего эксперимента. В работе были использованы L-Глутамат (L-Glutamic acid), антагонист D1 рецепторов SCH-23390 (R(+)-7-Chloro-8-hydroxy-3-methyl-1-phenyl-2,3,4,5-tetrahydro-1H-3-benzazepine hydrochloride), антагонист D2 рецепторов этиклопрайд (S-(–)-Eticlopride hydrochloride) фирмы SIGMA. Разведение веществ до необходимых концентраций проводилось перед каждым опытом в нормальном растворе (рН 7.4). Иллюстративный материал, отражающий изменение частоты импульсной активности во времени при действии исследуемых веществ, готовился с помощью пакета программ MS-Excel. Оценка достоверности результатов осуществлялась при помощи парного t-теста. Статистические решения принимались на 5%-м уровне значимости.
Изучение эффекта антагониста D1 рецепторов SCH-23390 на уровень фоновой активности сенсорных волокон заднего полукружного канала проводили в диапазоне концентраций 0.01–100 мкМ. Низкие концентрации SCH-23390 (0.01–10 мкМ) не изменяли ее уровня. Увеличение содержания антагониста до 100 мкМ приводило к резкому падению частоты фоновой активности, которая не восстанавливалась в пределах 15 мин.
Изучение эффекта антагониста D2 рецепторов этиклопрайда на разряд фоновой активности афферентных волокон проводилось в концентрациях 0.01–100 мкМ (рис. 1).
Рис. 1.
Эффект воздействия антагонистов D1 и D2 рецепторов допамина на уровень фоновой активности афферентных волокон, контактирующих с задним полукружным каналом лягушки. (а) Оригинальная запись типичного эксперимента с различными концентрациями антагониста D2 рецепторов этиклопрайда. (б) Суммарные кривые, отражающие ингибирующий эффект D1 и D2 антагонистов допамина на активность афферентных волокон. По оси абсцисс концентрация антагонистов допамина. По оси ординат: изменение частоты фоновой активности в % от фона. N = 11.

Перфузия синаптической области раствором D2 антагониста сопровождалась зависимым от концентрации позитивно-негативным изменением уровня фоновой активности. При концентрации 0.1 мкМ наблюдалось незначительное, но статистически значимое увеличение частоты фоновой активности (t-test 0.0023). Повышение концентрации сопровождалось уменьшением амплитуды позитивной волны ответа и увеличением его негативной фазы. При концентрации 0.1 мкМ 10 препаратов из 11 характеризовались позитивно-негативным ответом. При концентрации 1 мкМ соотношение составляло 4 из 11. Этиклопрайд в концентрации 10 мкМ статистически значимо подавлял фоновую активность на 28.8 ± ± 4.7% (t-test 0.0023). При концентрации этиклопрайда 100 мкМ на всех препаратах наблюдалось понижение уровня фоновой активности до 29.6 ± ± 6.2% от первоначального (t-test 0.0023). Отчетливый эффект сохранялся не менее 30 мин.
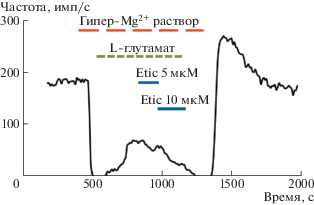
Для исследования возможной постсинаптической локализации рецепторов допамина проводили специальные опыты с блокадой выделения медиатора волосковой клеткой в гипермагниевом-гипокальциевом растворе. В этом случае добавление глутамата в гипермагниевый раствор на фоне блока синаптической передачи приводило к частичному восстановлению частоты импульсации, которая отражала активность рецепторов глутамата на постсинаптической мембране. Последующая аппликация растворов этиклопрайда демонстрировала модулирующее влияние антагониста D2 рецептора на постсинаптические глутаматные рецепторы (рис. 2).
Рис. 2.
Эффект воздействия антагониста D2 рецепторов этиклопрайда на активность постсинаптических глутаматных рецепторов после блокады пресинаптической мембраны гипермагниевым-гипокальциевым раствором. Оригинальная запись эксперимента.
Полученные результаты свидетельствуют о том, что в вестибулярном аппарате земноводных антагонисты D1 и D2 рецепторов допамина играют различную роль в модуляции уровня фоновой активности афферентных волокон. Антагонист D1 подавляет фоновую активность лишь при высоких концентрациях. Напротив, антагонист D2 вызывает зависимый от концентрации позитивно-негативный эффект. Можно предположить, что позитивная волна ответа при воздействии этиклопрайда может соответствовать взаимодействию антагониста D2 рецепторов с пресинаптическими D2 рецепторами, ингибированию тонического выделения допамина из эфферентных волокон и, как следствие, устранению его депрессивного влияния, что приводит к увеличению частоты импульсной активности за счет отсутствия тонического выделения допамина.
Развитие негативной волны при высоких концентрациях этиклопрайда можно объяснить его взаимодействием с постсинаптическими D2 рецепторами и последующим ингибированием постсинаптических глутаматных рецепторов. Показано, что пресинаптические ауторецепторы допамина могут активироваться более низкими концентрациями допамина, чем необходимо для активации постсинаптических рецепторов. Такая разница может объясняться различными сплайс вариантами D2 рецепторов [2–4].
Показанное влияние антагонистов D1 и D2 рецепторов допамина на постсинаптические глутаматные рецепторы подтверждает данные о постсинаптической локализации рецепторов допамина, полученные нами ранее с использованием D1 и D2 агонистов SKF-82958 и квинелорана (quinerolane) [5].
Учитывая тот факт, что в вестибулярном аппарате фоновая активность афферентных волокон отражает тоническое выделение глутамата волосковой клеткой [1] и активацию постсинаптических AMPA рецепторов [6], можно сделать вывод, что допамин, тонически выделяясь из эфферентных волокон, оказывает нейропротекторное влияние на активность глутаматэргического синапса и поддерживает его высокую пластичность.
ИСТОЧНИК ФИНАНСИРОВАНИЯ
Работа выполнена при поддержке Программы фундаментальных научных исследований государственных академий на 2013–2020 гг. (ГП-14, раздел 64).
Список литературы
Guth P.S., Perin P., Norris C.H., et al. The vestibular hair cells: post-transductional signal processing// Prog. Neurobiol. 1998. V. 54. P. 193–247.
De Mei C., Ramos M., Kinoshita C., et al. Getting specialized: presynaptic and postsynaptic dopamine D2 receptors // Curr. Opin. Pharmacol. 2009. V. 9. P. 53–58.
Beaulieu J.-M., Gainetdinov R.R. The physiology, signaling, and pharmacology of dopamine receptors // Pharmacol. Rev. 2011. V. 63. P. 182–217.
Usiello A., Baik J.H., Rouge´-Pont F., et al. Distinct functions of the two isoforms of dopamine D2 receptors // Nature. 2000. V. 408. P. 199–203.
Andrianov G.N., Ryzhova I.V., Tobias T.V. Dopaminergic modulation of afferent synaptic transmission in the semicircular canals of frogs // Neurosignals. 2009. V. 17. P. 222–228.
Bonsacquet J., Brugeaud A., Compan V., et al. AMPA type glutamate receptor mediates neurotransmission at turtle vestibular calyx synapse // J. Physiol. 2006. V. 576. P. 63–71.
Дополнительные материалы отсутствуют.
Инструменты
Доклады Российской академии наук. Науки о жизни