

Доклады Российской академии наук. Науки о жизни, 2020, T. 493, № 1, стр. 389-394
РАЗРУШЕНИЕ СТРУКТУРЫ ЗВУКОВОГО СИГНАЛА В КРАЕВОЙ ПОПУЛЯЦИИ (НА ПРИМЕРЕ СЕРОГО СУРКА Marmota baibacina)
А. А. Никольский 1, *, Е. А. Ванисова 1, **
1 Российский университет дружбы народов (РУДН)
Москва, Россия
* E-mail: bobak@list.ru
** E-mail: vanhelen@mail.ru
Поступила в редакцию 06.03.2020
После доработки 06.03.2020
Принята к публикации 10.03.2020
Аннотация
Описана изменчивость звукового предупреждающего об опасности сигнала серого сурка (Marmota baibacina) в периферической популяции в долине реки Арпа (Киргизия). Ранее сообщалось, что эта популяция обитает совместно с красным сурком (M. caudata). Красного сурка авторы не обнаружили. В краевой популяции серого сурка обнаружены две формы аберраций: амплитудная модуляция и отсутствие паузы между компонентами. Разрушение типичной для вида структуры сигнала связано, вероятно, с нарушением потока генов и не содержит явных признаков гибридизации. Материал собран в полевых условиях.
В работе представлены результаты анализа изменчивости структуры звукового сигнала в краевой, периферической популяции серого сурка (Marmota baibacina), обитающей, как сообщалось ранее [1, 2], в зоне контакта с соседом по ареалу – красным сурком (M. caudata). В периферических малочисленных популяциях в результате дрейфа генов изменчивость непредсказуема, так же, как и в зонах гибридизации. То и другое приводит к нарушению потока генов и генетической дезинтеграции популяций [3].
Консолидации популяций с родственным генотипом способствуют внутривидовые коммуникативные процессы, ключевую роль в которых играют генетически детерминированные средства коммуникации. В качестве модели изменчивости средств коммуникации мы используем доступный для массового сбора в полевых условиях предупреждающий об опасности сигнал, известный у многих видов млекопитающих. Сигнал генетически детерминирован, следствием чего является видовая специфика его структуры [4–6].
Преимущество звуковых сигналов, как модели изменчивости средств коммуникации, перед сигналами иной сенсорной модальности (например, химических) состоит в том, что звуковые сигналы поддаются визуализации и количественному описанию их параметров.
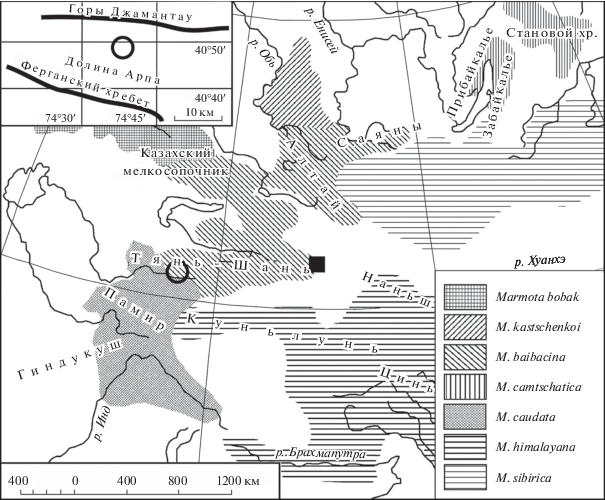
Материал собран в 2015 г. в Киргизии, в Восточном Тянь-Шане, в долине реки Арпа (рис. 1, далее – локальная популяция “Арпа”). Географические координаты: 40°50' с.ш.; 74°43' в.д.
Рис. 1.
Распространение серого сурка и его соседей по ареалу ([13] с изменениями). Кружком обозначена локальная популяции серого сурка “Арпа”, черным квадратом – “Урумчи”.
Сигнал записан на цифровой рекордер посредством направленного микрофона. Для анализа акустической структуры сигнала использовали программу SpectraLab. Статистическую обработку проводили с помощью программы Statgraphics. P ≤ 0.05. Общая выборка составляет 27 особей.
К обсуждению результатов мы привлекли собранные ранее записи сигнала красного сурка из Памиро-Алая, монгольского сурка (M. sibirica) − из различных районов Монголии, серого сурка − из Горного Алтая [4], а также собранные в 2014 г. записи сигнала серого сурка, населяющего восточные отроги Тянь-Шаня в Китае. Здесь, как и в долине Арпа, серый сурок представлен краевой популяцией (рис. 1). Географические координаты: 43°21' с.ш.; 87°01' в.д. Рабочее название популяции − “Урумчи”.
Совместное поселение красных и серых сурков в долине Арпа, о которых сообщали сотрудники противочумной системы [2, 3], мы не обнаружили.
Сигнал серого сурка был описан ранее [4]. Он состоит из двух компонентов, разделенных паузой, амплитудная модуляция отсутствует (рис. 2а, 2б).
Рис. 2.
Характеристика сигнала локальной популяции “Арпа”. Спектрограмма (а) и осциллограмма (б) типичной для серого сурка формы сигнала. Спектрограмма (в) и осциллограмма (г) сигнала с фрагментарной амплитудной модуляцией. (д) – фрагмент осциллограммы с высоким разрешением. (е) – спектрограмма сигнала с отсутствием паузы между низко- и высокочастотным компонентами, их место соединения обозначено кружком. (ж) – фрагмент осциллограммы в месте соединения низко- и высокочастотного компонентов. НЧ – низкочастотный компонент, П – пауза между низко- и высокочастотным компонентами, ВЧ – высокочастотный компонент, АМ – амплитудная модуляция, F0 – основная частота высокочастотного компонента, F1 – первая гармоника высокочастотного компонента, нарастающая (Нр) и затухающая (Зт) фазы характеристики частотной модуляции, ± Ωi– верхние и нижние боковые частоты, T0 – период основной частоты, Tам – период амплитудной модуляции.
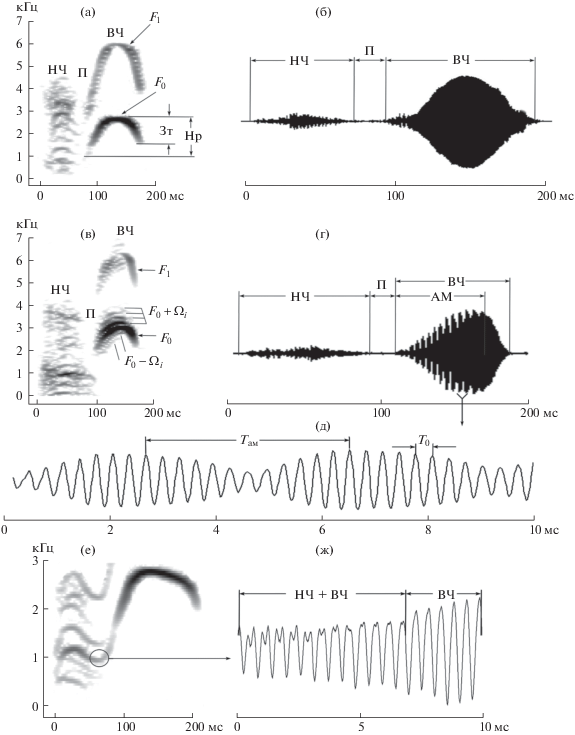
В популяции “Арпа” встречаются характерная для вида структура сигнала − фонотип (13 случаев из 27, 48.1%) и 2 формы отклонений от фонотипа: амплитудная модуляция и отсутствие паузы между низко- и высокочастотным компонентами.
Амплитудная модуляция, как и фонотип, присутствует в 48.1% случаев. Обычно это фрагментарная модуляция, когда модулирована часть сигнала, как показано на рис. 2г. Полная амплитудная модуляция обнаружена в одном случае из 13. Спектр модулированного сигнала выше и ниже несущей частоты и ее гармоник нагружен боковыми частотами (рис. 2в).
В 29.5% случаев в сигнале популяции “Арпа” отсутствует пауза между низко- и высокочастотным компонентами (рис. 2е, 2ж). Возможны сочетания амплитудной модуляции и отсутствия паузы между компонентами, поэтому в сумме мы не получаем 100%. Низко- и высокочастотный компоненты накладываются в момент перехода от одного компонента к другому. Они сформированы взаимно независимыми колебаниями, что подтверждается наличием двухвершинных пиков на осциллограммах (рис. 2ж).
Опыт предыдущих исследований показывает, что разрушение структуры сигнала может быть следствием дрейфа генов [7] и результатом гибридизации [8]. Учитывая, что амплитудная модуляция – самый характерный признак сигнала красного сурка [4], нельзя исключить следы гибридизации в сигнале популяции “Арпа”, несмотря на то, что красных сурков мы не обнаружили.
Но некоторые факты указывают на низкую вероятность того, что амплитудную модуляцию популяция “Арпа” унаследовала от красных сурков. Во-первых, это значительное отличие периода амплитудной модуляции сигнала красного сурка от периода амплитудной модуляции сигнала популяции “Арпа”. По этому признаку популяция “Арпа” ближе к монгольскому, чем к красному сурку (рис. 3, табл. 1). Во-вторых, в другой локальной популяции серого сурка, “Урумчи”, удаленной от ареала красного сурка (рис. 1), амплитудная модуляция обычна, а период модулирующего колебания мало отличается от периода модуляции сигнала красного сурка (рис. 3). Различия, главным образом, в глубине модулирующего колебания: у красного сурка она равна почти 1 ([4], рис. 3), в то время как у серого из популяции “Урумчи” значительно меньше. Не исключено, что сигнал локальной популяции “Урумчи”, не связанной с красным сурком, находится на промежуточной стадии отбора, на пути к сигналу с “полноценной” глубокой амплитудной модуляцией.
Рис. 3.
Амплитудная модуляция сигнала красного сурка (а), серого сурка локальной популяции “Урумчи” (б), серого сурка локальной популяции “Арпа” (в), монгольского сурка (г). (д) – фрагмент сигнала красного сурка, (е) – фрагмент сигнала серого сурка локальной популяции “Урумчи”. НЧ – низкочастотный компонент сигнала (см. рис. 2), Tam – период амплитудной модуляции сигнала, Aam – амплитуда модулирующего колебания.

Таблица 1.
Период амплитудной модуляции (Tам, мс) звукового предупреждающего об опасности сигнала сурков
| Вид, популяция | Tам, мс | ||
|---|---|---|---|
| n | $\bar {x}$ ± s.d. | lim | |
| M. caudata | 27 | 12.7 ± 1.57 | 10.4−15.9 |
| M. sibirica | 28 | 2.3 ± 0.34 | 1.8−2.9 |
| M. baibacina, “Арпа” | 13 | 3.7 ± 1.54 | 1.6−7.3 |
| M. baibacina, “Урумчи” | 8 | 8.7 ± 1.67 | 7.0−12.3 |
В сигнале серого сурка, записанном в Горном Алтае, ни в одном из 18 случаев, мы не обнаружили ни амплитудную модуляцию, ни отсутствие паузы между компонентами.
Оба отклонения от фонотипа серого сурка в сигнале популяции “Арпа”, наиболее вероятно, являются аберрациями, накопленными в генотипе краевой малочисленной популяции в результате дрейфа генов.
Как редчайшую аберрацию, амплитудную модуляцию мы обнаружили в изолированной популяции степного сурка (M. bobak) в Харьковской области [7].
Можно предположить, что появление сходных признаков звуковых сигналов независимо у разных видов является подтверждением Закона гомологических рядов в наследственной изменчивости Н.И. Вавилова [9].
Полученные нами результаты показывают глубокую деформацию структуры сигнала в краевой популяции, но мы не знаем, как сильно разрушение фонотипа влияет на выполнение сигналом его жизненно важной для популяции функции – предупреждение об опасности.
И так же мы не знаем, вызывает ли разрушение структуры сигнала генетическую дезинтегрированность популяций (см. [3]). Чтобы ответить на эти вопросы, необходимо провести специальные исследования. Учитывая относительную простоту регистрации реакции сурков на эмиссию данного сигнала, и доступность соответствующего оборудования, такие эксперименты возможны.
Краевые популяции являются аналогом локальных популяций в метапопуляциях, интерес к которым возрастает в связи с фрагментацией ареалов под влиянием антропогенных факторов [10–12]. Исследование изменчивости в краевых популяциях будет способствовать пониманию процессов, происходящих в создаваемых человеком метапопуляциях.
Список литературы
Майр Э. Зоологический вид и эволюция. М.: Мир, 1968. 597 с. Пер. с англ.: Mayr Ernst. Animal species and evolution. London: Oxford university press, 1963. 797 p.
Кулькова Н.А. // Труды Среднеазиатского противочумного института. Вып. 7. А-Ата – Фрунзе: Минздрав СССР, 1961. С. 377–378.
Семёнова Н.И. // Ресурсы фауны сурков в СССР (Географическое распространение, использование и восстановление запасов, эпидемиологическое значение). М.: Наука, 1967. С. 37–38.
Никольский А.А. Звуковые сигналы млекопитающих в эволюционном процессе. M.: Наука, 1984. 199 с.
Matrosova V.A., Rusin M.Y., Rashevska H.V., et al. // Mammalian Biology – Zeitschrift fur Saugetierkunde. 2016. T. 81. № 3. P. 255–265.
Matrosova V.A., Ivanova A.D., Volodin I.A., et al. // Integrative Zoology. 2019. T. 14. № 4. P. 341–353.
Никольский А.А. // Известия РАН. Серия биологическая. 2008. № 6. С. 718–729.
Никольский A.A., Янина И.Ю., Рутовская M.B. и др. // Зоологический журнал. 1983. Т. 62. Вып. 8. С. 1258–1266.
Вавилов Н.И. Закон гомологических рядов в наследственной изменчивости // Избранные произведения в двух томах. Т. 1. Л.: Наука, 1967. С. 7–61.
Закс С.С., Кузьмин А.А., Симаков М.Д. и др. // Известия высших учебных заведений. Поволжский регион. Естественные науки. 2018. № 3 (23). С. 80–91.
Закс С.С., Кузьмин А.А., Симаков М.Д. и др. // Известия высших учебных заведений. Поволжский регион. Естественные науки. 2018. № 4 (24). С. 42–50.
Салменкова Е.А. // Успехи современной биологии. 2018. Т. 138. № 1. С. 3–11.
Никольский А.А., Румянцев В.Ю. // ДАН. 2012. Т. 445. № 4. С. 175–178.
Дополнительные материалы отсутствуют.
Инструменты
Доклады Российской академии наук. Науки о жизни