

Доклады Российской академии наук. Науки о жизни, 2020, T. 493, № 1, стр. 413-416
АНАЛИЗ ГЕОГРАФИЧЕСКОЙ ИЗМЕНЧИВОСТИ МОРФОГЕНЕТИЧЕСКИХ ТРАЕКТОРИЙ НА ПРИМЕРЕ ОБЫКНОВЕННОЙ СЛЕПУШОНКИ (Ellobius talpinus Pall.)
А. Г. Васильев 1, *, академик РАН В. Н. Большаков 1, И. А. Васильева 1, Н. В. Синева 1
1 Институт экологии растений и животных
Уральского отделения Российской академии наук
Екатеринбург, Россия
* E-mail: vag@ipae.uran.ru
Поступила в редакцию 02.10.2019
После доработки 26.12.2019
Принята к публикации 20.02.2020
Аннотация
Впервые методами геометрической морфометрии выявлен феномен географической изменчивости морфогенетических траекторий при параллельном сравнении в общем морфопространстве формы нижней челюсти разновозрастных групп трех популяций обыкновенной слепушонки (Ellobius talpinus Pall.) в Южном Зауралье.
Географическая изменчивость обычно характеризует устойчивые межпопуляционные различия по тому или иному признаку, а также проявление внутривидовой дифференциации и дивергенции [1, 2]. Географическую изменчивость разных стадий постнатального морфогенеза до настоящего времени не изучали. Геометрическая морфометрия [3–5], позволяющая отдельно изучить изменчивость размеров и формы, допускает морфогенетическую интерпретацию изменений формы биологических объектов как многомерного единичного признака [3, 5] и может способствовать решению данной проблемы.
Ранее П. Олберч с соавторами [6] обосновали представления об онтогенетических траекториях, развитые позднее М.В. Миной [7, 8], которые характеризуют последовательность фенотипических изменений особей на разных этапах постнатального онтогенеза.
Геометрическая морфометрия позволяет описывать изменение формы объектов в онтогенезе, исключая их размеры, поэтому в данном случае речь идет о морфогенетических изменениях [3–5]. Поэтому мы предложили последовательное изменение формы объекта в морфопространстве на разных этапах его постнатального развития определить как морфогенетическую траекторию [5]. Гипотетические морфогенетические траектории географически удаленных популяций можно представить в морфопространстве как следующие вероятные типы: а – синтопный, когда траектории почти идентичны, б – параллельный – смещены относительно друг друга без расхождения; в – дивергентный, когда с возрастом траектории расходятся; г – эквифинальный, когда исходно удаленные траектории в конечном итоге сближаются; д – аллотопный, если траектории направлены почти параллельно, но сдвинуты относительно друг друга; е – аллотопно-дивергентный – то же, но с расхождением траекторий (рис. 1).
Рис. 1.
Гипотетические типы морфогенетических траекторий (этапы морфогенеза: juv – ювенильный. ad – взрослый, sen – сенильный) географически удаленных популяций вида (1, 2) в общем морфопространстве: (а) синтопный, (б) параллельный, (в) дивергентный, (г) эквифинальный, (д) аллотопный, (е) аллотопно-дивергентный.

Цель исследования – проверка гипотезы о том, что географическая изменчивость может проявиться не только на уровне отдельных фенотипических признаков, но и морфогенетических траекторий. Основной задачей в этой связи было изучение методами геометрической морфометрии географической изменчивости формы нижней челюсти у разновозрастных групп особей из географически удаленных популяций модельного вида грызунов – обыкновенной слепушонки (Ellobius talpinus Pall., 1770) в Южном Зауралье.
Использовали коллекции Музея ИЭРиЖ УрО РАН, включающие выборки из трех удаленных популяций слепушонки: 1 – кунашакская (п. Кунашак, Челябинской обл. – 95 экз.: 55°35′ с.ш., 61°40′ в.д.), 2 – баймакская (п. Баймак, Республика Башкортостан – 106 экз.: 52°33′ с.ш., 58°13′ в.д.), 3 – наурзумская (Наурзумский заповедник, Сев. Казахстан – 106 экз.: 51°31′ с.ш., 64°29' в.д.). Географическое положение популяций показано на карте-схеме (рис. 2а). Южная наурзумская популяция расположена примерно на той же долготе, что и северная кунашакская, а географически промежуточная – баймакская – несколько смещена от них в западном направлении.
Рис. 2.
Географическое положение кунашакской (K), баймакской (B) и наурзумской (N), популяций обыкновенной слепушонки в Южном Зауралье (а); схема размещения меток-ландмарок (1–15) на лингвальной стороне правой нижней челюсти (б); ординация центроидов возрастных групп (1–5) и построенных по ним траекторий изменения формы нижней челюсти в общем морфопространстве вдоль первых трех главных компонент – PC1–PC3 (в).
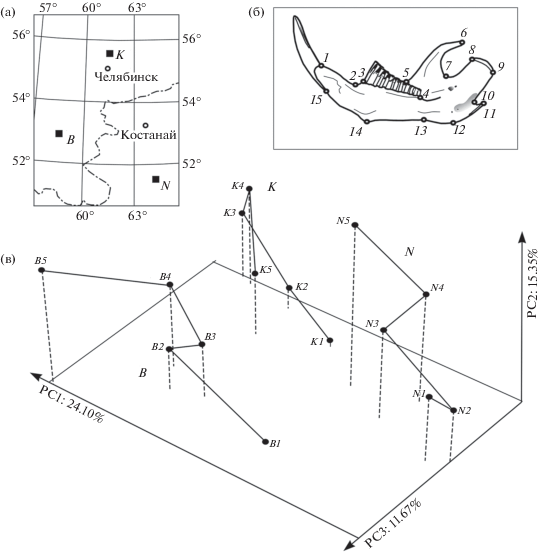
Возраст определен по степени развитости корней первого нижнего щечного зуба m1 [9]. Выборки представлены особями 5 возрастных групп: 1 – сеголетки 0+, 2 – 1+, 3 – 2+, 4 – 3+, 5 – сборная группа сенильных зверьков 4+ и старше (до 6 лет). В качестве объекта исследования выбрана нижнюю челюсть, поскольку слепушонка активно использует нижние резцы при рытье и добывании корма, а ее морфологическая изменчивость отражает функциональную адаптацию к локальным условиям [10]. С помощью программ пакета TPS [11, 12] на цифровые изображения правой нижней челюсти (лингвальная сторона) поместили конфигурации 15 меток-ландмарок (landmarks), позволяющие характеризовать изменчивость ее формы (рис. 2б). В работе применили генерализованный Прокрустов анализ на основе наименьших квадратов (GPA) [3], а прокрустовы координаты (Procrustes coordinates), характеризующие изменчивость формы мандибул, использовали для межгруппового сравнения. Статистический анализ выполнен с помощью программ TPS [11, 12], PAST 2.17с [13] и MorphoJ 1.6d [4].
Методом главных компонент провели ординацию прокрустовых координат, а затем, соединив линией средние значения каждой последующей возрастной группы в трехмерном морфопространстве, образованном первыми тремя главными компонентами (PC1–PC3), построили “морфогенетические траектории” для трех сравниваемых популяций (рис. 2в).
Доля объясненной дисперсии вдоль первых трех главных компонент составила 51.1%. Значимые корреляции Спирмена между значениями осей с полом не выявлены, что позволяет сравнить объединенные по полу выборки.
Локализация морфогенетических траекторий трех популяций в общем морфопространстве во многом согласуется с их взаимным географическим положением в Южном Зауралье. Ординаты вдоль первой и третьей осей значимо связаны с географическим положением популяций (PC1 с широтой rsp = 0.72, p = 0.0025 и с долготой rsp = 0.70, p = 0.0037; PC3 только с долготой rsp = 0.68, p = 0.0054), а вдоль второй (PC2) – только с возрастом (rsp = 0.80, p = 0.0003). Четвертая (PC4) и пятая (PC5) компоненты также значимо связаны с географическим положением и возрастом (p < 0.01). Однако их вклад в общую дисперсию мал (соответственно 7.7 и 6.9%), поэтому мы их здесь отдельно не рассматриваем. Непараметрический многомерный двухфакторный дисперсионный анализ (two-way PERMANOVA) с применением 10000 перестановок (Permutation) по комплексу первых пяти главных компонент выявил значимые эффекты географического положения выборок с учетом широты (F = 16.63; p < 0.0001) и возраста (F = 5.46; p < 0.0001), но взаимодействие этих факторов оказалось незначимым (p = 0.9306).
Морфогенетические траектории популяций занимают разные области морфопространства (см. рис. 2в), поэтому можно полагать, что морфогенез нижней челюсти специфичен для каждой популяции. Траектория кунашакской популяции смещена относительно наурзумской, а промежуточное положение на рисунке со смещением относительно их влево занимает траектория баймакской популяции. Если сравнить расположение в морфопространстве ординат одноименных возрастных групп трех популяций, то на всех последовательных этапах морфогенеза форма нижней челюсти будет различаться пропорционально их географической удаленности. Таким образом, локализация ординат всех возрастных групп, формирующих морфогенетические траектории в общем морфопространстве трех сравниваемых популяций, отражает географическую изменчивость онтогенетических изменений конфигурации нижней челюсти. При этом общее направление морфогенетических траекторий совпадает: во всех популяциях траектории изменений формы мандибул от наиболее молодых к сенильным группам ориентированы почти параллельно. Данное направление отчасти согласуется со смещением северных выборок относительно южных, так как группы молодых зверьков во всех популяциях в морфопространстве расположены условно “южнее”, а сенильных – “севернее”. Поскольку расположение морфогенетических траекторий в морфопространстве коррелирует с их географическим положением, то выявленный характер изменения траекторий наиболее соответствует описанным выше аллотопному или аллотопно-дивергентному типам (см. рис. 1). В итоге можно заключить, что данный роющий вид, постепенно проникая из южных частей ареала в северные (кунашакская популяция обитает на северной границе ареала), выработал специфичные для каждой локальной популяции особенности постнатального морфогенеза мандибул, которые затронули практически все возрастные стадии. Таким образом, популяционная структура и дифференциация данного вида могут являться примерами осуществления начальных стадий процесса географического формообразования, а морфогенетические различия между популяциями, вероятно, отражают последовательные этапы микроэволюционных изменений слепушонки при освоении видом степных и лесостепных ландшафтов Южного Зауралья.
Список литературы
Яблоков А.В. Изменчивость млекопитающих. М.: Наука, 1966. 364 с.
Майр Э. Зоологический вид и эволюция. М.: Мир, 1968. 597 с.
Zelditch M.L., Swiderski D.L., Sheets H.D., Fink W.L. Geometric Morphometrics for Biologists: A Primer. New York: Elsevier Academic Press, 2004. 437 p.
Klingenberg C.P. // Hystrix, the Italian Journal of Mammal. 2013. V. 24. № 1. P. 43–58.
Васильев А.Г., Васильева И.А., Шкурихин А.О. Геометрическая морфометрия: от теории к практике. М.: Тов-во научных изданий КМК. 2018. 471 с.
Alberch P., Gould S.J., Oster G.F. et al. // Paleobiology. 1979. V. 5. P. 296–317.
Мина М.В. // Онтогенез. 2001. Т. 32. № 6. С. 471–476.
Мина М.В., Шкиль Ф.Н., Абдисса Белай // Вопросы ихтиологии. 2010. Т. 50. № 4. С. 471–479.
Евдокимов Н.Г. Популяционная экология обыкновенной слепушонки (Ellobius talpinus Pall., 1770) Южного Урала и Зауралья. Екатеринбург: Екатеринбург, 2001. 144 с.
Васильев А.Г., Большаков В.Н., Евдокимов Н.Г., Синева Н.В. // ДАН. 2016. Т. 468. № 1. С. 118–121.
Rohlf F.J. tpsUtil. Vers. 1.60. Stony Brook: Depart. Ecol. and Evolut.; State Univ. N.Y., 2013.
Rohlf F.J. tpsDig2. Vers. 2.17. Stony Brook: Depart. Ecol. and Evolut.; State Univ. N.Y., 2013.
Hammer Ø., Harper D.A.T., Ryan P.D. // Palaeontol. Electronica. 2001. V. 4. № 1. 9 p.
Дополнительные материалы отсутствуют.
Инструменты
Доклады Российской академии наук. Науки о жизни