

Доклады Российской академии наук. Науки о жизни, 2020, T. 495, № 1, стр. 567-571
САБЛЕЗУБЫЕ КОШКИ (Carnivora, Felidae, Machairodontinae) ИЗ НИЖНЕГО ПЛЕЙСТОЦЕНА ПЕЩЕРЫ ТАВРИДА, КРЫМ
А. В. Лавров 1, *, Д. О. Гимранов 2, 3, Д. Б. Старцев 4, Б. А. Зайцев 5
1 Палеонтологический институт им. А.А. Борисяка Российской академии наук
Москва, Россия
2 Институт экологии растений и животных
Уральского отделения Российской академии наук
Екатеринбург, Россия
3 Уральский федеральный университет им. Б.Н. Ельцина
Екатеринбург, Россия
4 Крымский федеральный университет
им. В.И. Вернадского
Симферополь, Россия
5 Всероссийский научно-исследовательский геологический институт им. А.П. Карпинского
Санкт-Петербург, Россия
* E-mail: lavrov_av@inbox.ru
Поступила в редакцию 17.02.2020
После доработки 05.05.2020
Принята к публикации 07.05.2020
Аннотация
Из раннеплейстоценового местонахождения пещера Таврида (Крым, поздний виллафранк, возраст 1.8–1.5 млн лет) описаны верхнечелюстные кости взрослых экземпляров саблезубых кошек Homotherium crenatidens (Fabrini, 1890) и Megantereon sp. Отмечается высокая морфометрическая изменчивость H. crenatidens в строении верхнего хищнического зуба и длине диастемы С1–Р3.
Пещера Таврида была открыта в 2018 г. Она расположена на северном макросклоне Крымских гор, в междуречье рек Бештерек и Фундуклы (притоки р. Зуи, бассейн р. Салгир). В южном коридоре пещеры было обнаружено большое число костей ископаемых наземных позвоночных. Этот комплекс соответствует поздневиллафранкским фаунам Европы (зоны MNQ18–MNQ19) и псекупскому фаунистическому комплексу России. Время существования ассоциации определено в интервале 1.8–1.5 млн л.н. [1],
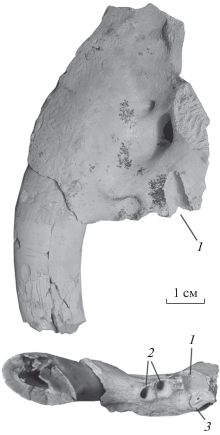
В ходе раскопок в 2018 г. были найдены остатки представителей Machairodontinae. Homotherium crenatidens (Fabrini, 1890) представлен правой верхнечелюстной костью с фрагментом клыка, альвеолой Р3, целым Р4 и альвеолой М1 (экз. КФУ, № 10-145; рис. 1). Обнаружены также изолированные резцы Homotherium: I3 (экз. ПИН, № 5644/62) и I2 (экз. ПИН, № 5644/63). Megantereon представлен изолированным левым i3 (экз. ПИН, № 5644/64) и фрагментом левой верхнечелюстной кости с частью клыка, альвеолой Р3 и передней альвеолой Р4 (экз. ПИН, № 5644/60; рис. 2).
Рис. 1.
Homotherium crenatidens (Fabrini, 1890), экз. КФУ, № 10-145, верхнечелюстная кость с С1, Р4 и альвеолами Р3 и М1; вид с латеральной стороны (вверху слева), вентральной (внизу слева), вид с краниомедиальной стороны на вскрытые корни Р4 (справа); нижний плейстоцен, пещера Таврида, Крым, Россия. Обозначения: 1 – корень протокона; 1а – выступ рудиментарного протокона; 2 – корень парастиля; 3 – фрагменты твердого нёба; 4 – альвеола М1; 5 – большое нёбное отверстие (foramen palatinum majus); 6 – альвеола Р3.

Рис. 2.
Megantereon sp., экз. ПИН, № 5644/60, фрагмент верхнечелюстной кости с С1 и альвеолами Р3 и частично сохранившейся альвеолой Р4, вид с латеральной (вверху) и вентральной (внизу) стороны; нижний плейстоцен, пещера Таврида, Крым, Россия. Обозначения: 1 – альвеола корня парастиля; 2 – альвеолы Р3; 3 – корень протокона.
Описанные в статье челюстные материалы хранятся в Палеонтологическом институте им. А.А. Борисяка РАН, Москва (ПИН), и Крымском федеральном университете им. В.И. Вернадского, Симферополь (КФУ). Промеры зубов (максимальная длина и ширина) проводились по общепринятой методике [2]. Измерения приведены в мм.
Верхнечелюстная кость H. crenatidens с сильно стертым Р4, принадлежит старой особи. Отнесение к данному виду основано на морфологии зубного ряда, строении Р4 и его корней (см. ниже). Фрагмент С1 имеет участки сохранившейся зазубренной поверхности режущих гребней, характерной для этого рода. Форма клыка в поперечном сечении отличается от таковой Megantereon (рис. 3) и является типичной для Homotherium, имеются передний и задний режущие гребни. Нервный канал С1 сильно склеротизирован. На задней стороне клыка зазубренный гребень развит до края альвеолы клыка.
Рис. 3.
Поперечное сечение верхнего правого клыка вблизи основания (вид на сечение клыка снизу) у предствителей Machairodontinae: а – Homotherium crenatidens (Fabrini, 1890), экз. КФУ, № 10-145, правый клык; б – Megantereon sp., экз. ПИН, № 5644/60, правый клык; нижний плейстоцен, пещера Таврида, Крым. Обозначения: 1 – заднее лезвие; 2 – переднее лезвие; 3 – нервный канал. Стрелка вверх означает лабиальное направление, стрелка вправо – переднее направление.

Р3 однокорневой (рис. 1), размеры альвеолы 6.3 × 4.8 мм (у большинства экземпляров H. crenatidens из Европы и Таджикистана Р3 также однокорневой), длина (L) коронки Р3 составляет 5.5–11.5 мм (n = 12; табл. 1) [2, 3].
Таблица 1.
Размеры верхнечелюстного зубного ряда у H. crenatidens (Fabrini, 1890) и Megantereon sp. (L – длина, W – ширина)
| Образцы/ промеры |
H. crenatidens экз. КФУ № 10-145 | H. crenatidens Таджикистан (Сотникова, 1988) |
H. crenatidens Европа (Viret, 1954; Sardella, Iurino, 2012; Palmquis et al., 2007) |
Meganthereon sp., экз. ПИН № 5644/60 | M. cultridens Европа (Palmquis et al., 2007) |
|---|---|---|---|---|---|
| LC1 | 28.5 | 32.4–33.01 | 25.0–38.2 | 23.4 | 17.1–29.0 |
| WC1 | 13.3 | 12.2–13.4 | 10.0–17.2 | 12.6 | 8.5–16.5 |
| LP3 | 6.31 | 9.01–11.5 | 5.5–11.0 | 10.91 | 12.2–18.5 |
| WP3 | 4.81 | 5.0–6.5 | 3.5–7.0 | 5.11 | 6.0–9.8 |
| LP4 | 37.6 | 35.11–40.0 | 37.0–45.3 | – | 26.8–36.0 |
| WP4 | 11.1 | 12.81–13.1 | 10.0–13.8 | 10.51 | 11.2–18.3 |
| LM1 | 4.11 | – | – | – | 4.0–7.8 |
| WM1 | 11.11 | – | – | – | 7.8–11.6 |
| длина диастемы Ld-C1-P3 | 21.8 | 13.3–16.0 | – | 11.0 | – |
| LC1-P4 | 98.2 | 96.0–98.01 | – | – | – |
| LC1-M1 | 105.0 | 100.42 | – | – | – |
Расстояние от переднего края альвеолы Р3 до переднего края Р4 у экз. КФУ, № 10-145 равно 10.9. Задний край коронки P3 у Homotherium, как правило, вплотную примыкает к Р4. Поэтому для экз. КФУ, № 10-145 реконструируемая длина коронки Р3 (L рек.) составляет 11.5 мм. Можно заключить, что размеры Р3 у гомотерия из Крыма близки к максимальным размерам Р3 у H. crenatidens.
Лабиальный цингулюм Р4 развит хорошо, что характерно для H. crenatidens. Парастиль крупный. Экторапастиль слабо различим. Его граница с парастилем из-за cильного износа зуба почти не заметна (рис. 1). Длина метастиля (от хищнической вырезки) составляет 47% длины Р4. Корень под параконом небольшой и примерно равен корню под протоконом (как и на образцах из других регионов). Корень под протоконом уплощен лабиолингвально. На экз. КФУ, № 10-145 этот корень почти на всем своем протяжении слит с корнем парастиля и только самый его конец является свободным. М1 располагался на костном гребне, образующим постеролабиальный край твердого нёба, и был высоко приподнят над его поверхностью. Судя по альвеоле, зуб имел один корень.
Считается, что в Европе и Черноморско-Кавказском регионе в виллафранское время существовал вид H. crenatidens (Fabrini, 1890) [4–6]. Ключевые отличия поствиллафранского H. latidens (Owen, 1846) от H. crenatidens заключаются в строении резцов и нижней челюсти [6], морфологии премоляров, усложнении зазубренности верхних клыков у H. latidens [7–9]. У H. latidens (включая морфотип “H. moravicum”) на Р4, как правило, корень протокона редуцирован или отсутствует, корень паракона отсутствует. Стратиграфическая граница между двумя видами Homotherium не является четкой. В составе таманского териокомплекса (конец позднего виллафранка–начало галерия) в Приазовье существовал H. latidens [6]. В размерах зубов Homotherium отмечается значительная трансгрессия, как между видами рода, так и с представителями Megantereon [3].
Длина диастемы С1–Р3 составляет 21.8 мм, существенно больше, чем у находок из Таджикистана. По размеру зубов и длине зубного ряда С1–М1 (табл. 1) размеры животного сходны с размерами H. crenatidens из Таджикистана [2]. Размеры Р4 и С1 у экз. КФУ, № 10-145 средние для вида [2, 3].
Обращает на себя внимание большая изменчивость параметров зубного ряда. У H. crenatidens размеры Р3 могут различаться по длине и ширине в два раза (табл. 1) [10, 3, 11, 9 ]. Р3 может быть одно- или двухкорневым, корни могут быть частично слитыми или раздельными.
У H. crenatidens Р4 имеет четыре корня: большой корень парастиля, небольшой корень паракона (расположен непосредственно лабиальнее корня протокона и меньше него), небольшой, часто уплощенный лабиолингвально корень протокона и крупный корень метакона. Такой полный набор корней Р4 свойственен также Machairodus.
Следует отметить, что у экз. КФУ, № 10-145 на Р4 корень протокона слит с корнем парастиля на всем протяжении. Выступ протокона Р4 у H. crenatidens рудиментарный или отсутствует. Считается, что у H. crenatidens “корень протокона на Р4, как правило, есть” [2]. О его наличии часто можно судить только после вскрытия кости или рентгенографического исследования. В связи с редукцией выступа протокона его корень может быть добавочным корнем на корне парастиля. У экз. КФУ, № 10-145 корень протокона крупный, уплощен лабиолингвально. Он смещен лабиально под основание паракона и плотно, со срастанием, прилегает к крупному корню парастиля (рис. 1).
У H. latidens Р4 имеет два крупных корня: корень парастиля и корень метакона. Корень паракона, как правило, полностью редуцирован [3]. Корень протокона часто сохраняется, но рудиментарный (тонкий и короткий). Например, на экз. № МР 158 (Монте-Апулия, Италия) есть небольшой корень протокона. На части экземпляров H. latidens на Р4 корня протокона нет (экз. G21, Пирро-Норд, Италия; экз. 5410, Странска-Скала, Чехия) [3].
Таким образом, строение корней Р4 важно для видовой диагностики находок Homotherium, особенно фрагментарных. Изменчивость представителей Homotherium по размерам альвеолы клыка С1, диастемы С1–Р3 и Р3 значительно перекрывает амплитуду значений этих промеров у Megantereon (табл. 1).
Фрагмент левой верхнечелюстной кости Megantereon sp. (экз. ПИН, № 5644/60) принадлежит взрослой нестарой особи, что установлено по состоянию кости и ее швов, а также большому диаметру нервного канала С1 (рис. 3). Таксономическое определение сделано на основании малой длины альвеолы С1, форме поперечного сечения клыка, отсутствию зазубренности на его режущем лезвии, по наличию короткой диастемы С1–Р3 и крупной альвеолы двухкорневого Р3 (рис. 2).
С1 узкий, его передний край округлый. На зубе развит только задний гладкий режущий гребень (без пильчатого края). Форма поперечного сечения клыка типичная для Megantereon, она каплевидная (рис. 3). Длина альвеолы клыка (23.4 мм) меньше (табл. 1), чем минимальные значения у H. crenatidens (25.0 мм). Значение длины альвеолы клыка у представителей Megantereon варьирует в пределах 16.9–33.0 мм (n = 29) [12]. Размер альвеолы Р3: длина составляет 10.9 мм, ширина 5.1 мм. Судя по альвеоле (табл. 1), размеры Р3 были близки к минимальным для M. cultridens. Реконструированная длина зуба P3 составляет 12.5 мм. Задний корень зуба немного крупнее переднего. Длина диастемы у экз. ПИН, № 5644/60 составляет 11.0, что близко к значению у M. cultridens из Сенеза (SE 311). У Megantereon sp. из Крыма корень протокона P4 изолирован от основания корня парастиля, не прирастает к корню паракона.
Присутствие H. crenatidens и Megantereon sp. в отложениях пещеры Таврида согласуется с ранее сделанным заключением о возрасте фаунистического комплекса [1]. На основании размеров зубов и альвеол можно предположить, что описанный экземпляр принадлежит виду Megantereon cultridens (Cuvier, 1824); однако недостаточная сохранность материала не позволяет сделать однозначное заключение о его видовой принадлежности.
Список литературы
Лопатин А.В., Вислобокова И.А., Лавров А.В. и др. Пещера Таврида–новое местонахождение раннеплейстоценовых позвоночных в Крыму // ДАН. 2019. Т. 485. № 3. С. 40–43. https://doi.org/10.31857/S0869-56524853381-385
Сотникова М.В. Отряд Carnivora // Биостратиграфия позднего плиоцена-раннего плейстоцена Таджикистана. М: Наука, 1988. С. 38–49.
Sardella R., Iurino D.A. The latest Early Pleistocene sabertoothed cat Homotherium (Felidae, Mammalia) from Monte Peglia (Ubria, central Italy) // Boll. Soc. Paleontol. Ital. 2012. V. 51. № 1. P. 15–22.
Fiscarelly G. The Villafranchian machairodonts of Tuscany // Paleontographica Italica. 1979. V. 71 (n.s. 41). P. 17–26.
Bishop M.J. The Mammalian fauna in the Early Middle Plistocene cavern infull site of Westbury-sub-Mendip, Somerset // Spec. paper in Paleonology. 1982. № 28. P. 1–106.
Sotnikova M.V., Titov V.V. Carnivora of the Tamanian faunal unit (the Azov Sea area)// Quat. Intern. 2009. V. 201. P. 43–52. https://doi.org/10.1016/j.quaint.2008.05.019
Galobart À.L. Phylogeny, anatomy and palaeobiology of the machairodont cat Homotherium latidens (Owen) (Felidae, Carnivora) // Acta Paleontologia. 2007. V. 12. P. 57–68.
Christiansen P. Phylogeny of the sabertoothed felids (Carnivora: Felidae: Machairodontinae) // Cladistics. 2013. V. 29. P. 543–559.
Diedrich C.G., McFarlane D.A. Homotherium from Middle Pleistocene archaeological and carnivore den sites of Germany. Taxonomy, taphonomy and a revision of the Schöningen, West-Runton and other saber-tooth cat sites // Quat. Intern. 2017. V. 436. P. 76–83.https://doi.org/10.1016/j.quaint.2016.10.015
Viret M.J. Le loess a bancs dursis de Saint-Vallier // Nov. Arch. Museum Natur. Lion. 1954. Fasc. 4. P. 1–104.
Antón M., Salesa M. J., Galobart A., Tseng Z. J. The Plio-Pleistocene scimitar-toothed felid genus Homotherium Fabrini, 1890 (Machairodontinae, Homotherini): diversity, palaeogeography and taxonomic implications // Quat. Sci. Rev. 2014. V.96. P. 259–268.
Palmquist P., Torregrossa V., Perez-Claros J. et al. A re-evaluation of the diversity of Megantereon (Mammaila, Carnivora, Machairodontinae) and the problem of species identification in extinct carnivores // J. Vertebrate Paleontology. 2007. V.27. № 1. P. 160–175.
Дополнительные материалы отсутствуют.
Инструменты
Доклады Российской академии наук. Науки о жизни