

Доклады Российской академии наук. Науки о жизни, 2020, T. 495, № 1, стр. 577-580
БОЛЬШОЙ СВИНОЙ БАРСУК Arctonyx collaris (Mustelidae, Carnivora) ИЗ ПЛЕЙСТОЦЕНА ВЬЕТНАМА (пещера Лангчанг)
Академик РАН А. В. Лопатин *
Палеонтологический институт им. А.А. Борисяка Российской академии наук
Москва, Россия
* E-mail: alopat@paleo.ru
Поступила в редакцию 23.06.2020
После доработки 10.07.2020
Принята к публикации 10.07.2020
Аннотация
Описаны зубные остатки крупного барсука из плейстоценовых отложений карстовой пещеры Лангчанг в северном Вьетнаме (сборы 2020 г.). Коренные зубы М1 и М1 характеризуются признаками, свойственными Arctonyx collaris Cuvier, 1825. При этом по длине M1 форма из пещеры Лангчанг превосходит современных больших свиных барсуков. На основании величины и строения зубов она может быть отнесена к подвиду A. c. rostratus Matthew et Granger, 1923, известному из плейстоцена Китая и Юго-Восточной Азии.
Большой свиной барсук Arctonyx collaris Cuvier, 1825 – один из самых крупных современных видов барсуков (кондилобазальная длина черепа до 172–174 мм, длина тела до 87–88 см, вес до 14–21.5 кг) [1–3]. В настоящее время этот вид обитает на территории восточной Индии и в Индокитае вплоть до полуостровных частей Мьянмы и Таиланда [1, 3]. В среднем – позднем плейстоцене A. collaris был обычным представителем фауны Stegodon–Ailuropoda Китая и Юго-Восточной Азии; при этом ископаемые формы, как правило, рассматриваются в составе вымершего подвида A. collaris rostratus Matthew et Granger, 1923 [1, 4]. В результате изучения изменчивости зубной морфологии систематическая принадлежность части ископаемых экземпляров из Китая недавно была пересмотрена в пользу рода Meles [5], поэтому в случае находок изолированных зубов Arctonyx и Meles необходимо их детальное морфологическое описание, включающее характеристику вариантов строения.
Ниже описаны зубные остатки барсуков из плейстоценовых отложений карстовой пещеры Лангчанг (Lang Trang) в северном Вьетнаме (провинция Тханьхоа, уезд Батхыок), собранные в марте 2020 г. экспедиционным отрядом Палеонтологического института им. А.А. Борисяка РАН (ПИН). Ранее из пещеры Лангчанг указывались 33 экземпляра A. collaris [6: табл. 1 ], без их изображений и характеристики.
Терминология строения, морфотипы и методика измерения зубов по [5]. Размеры в мм. Обозначение измерений: L – наибольшая длина, EL – лабиальная длина (паракон – метакон или паракон – метастиль при наличии последнего), IL – лингвальная длина, ETRL – лабиальная длина тригонида (параконид – гипофлексид), ITRL – лингвальная длина тригонида (параконид – метаконид), W – наибольшая ширина, TRW – ширина тригонида, TALW – ширина талонида; μ – среднее значение.
Образцы хранятся в коллекции ПИН, г. Москва. Они представлены двумя изолированными M1 (экз. ПИН, №№ 5792/1, 5792/55) и одним М1 в небольшом фрагменте зубной кости (экз. ПИН, № 5792/2).
М1 (экз. ПИН, № 5792/1) характеризуется признаками, более свойственными Arctonyx нежели Meles [5]. Коронка относительно узкая (рис. 1а). Лабиальная вырезка между параконом и метаконом слабая (морфотип A2), вырезка позади метаконуля умеренно выраженная (морфотип C2). Между метаконом и метаконулем нет лабиальной вырезки (морфотип B1), но имеется легкая складка, окаймленная слабо выраженным цингулоподобным метастилем (морфотип D2). Лингвальной вырезки нет. Антеролингвальный угол лингвального цингулюма выступает вперед приблизительно до уровня вершины паракона (морфотип E4). Постеролабиальный гребень (между дистальным бугорком протокона и метаконулем) отсутствует (морфотип G1), постеролингвальный гребень (между дистальным бугорком протокона и гипоконом) слабо развитый, неполный (морфотип H2). На талоне имеется один сравнительно крупный дополнительный бугорок около метаконуля (морфотип F3) и еще один маленький дополнительный бугорок вблизи гипокона.
Рис. 1.
Arctonyx collaris rostratus Matthew et Granger, 1923: а – экз. ПИН, № 5792/1, правый M1 с окклюзиальной стороны, б – экз. ПИН, № 5792/55, левый M1 с окклюзиальной стороны; Вьетнам, провинция Тханьхоа, уезд Батхыок, пещера Лангчанг; верхний плейстоцен.
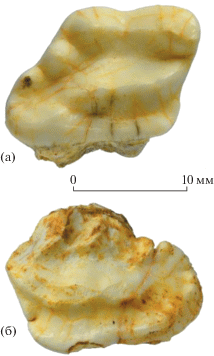
Размеры М1 (экз. ПИН, № 5792/1): L – 17.0, EL – 13.3, IL – 15.5, W – 13.0. Диапазон изменчивости размеров М1 для современных представителей вида [1: табл. 3 ]: L – 14.1–19.1 (μL – 16.7), W – 9.4–12.2 (μW – 11.2). Размеры М1 у плейстоценового A. c. rostratus из Китая [4: табл. 2 ]: L – 16.0–20.0 (μL – 18.0), W – 10.6–12.9 (μW – 11.8).
Экз. ПИН, № 5792/55 (разрушенный в области паракона и метакона) характеризуется узким талоном с пятью мелкими дополнительными бугорками (морфотип F2). Вырезка позади метаконуля хорошо выражена (морфотип C3). Лингвальная вырезка очень слабая (рис. 1б). Антеролингвальный угол лингвального цингулюма выступает вперед заметно слабее паракона. Постеролабиальный гребень отсутствует (морфотип G1), постеролингвальный гребень отчетливый (морфотип H3).
М1 (экз. ПИН, № 5792/2) длинный и узкий (рис. 2). Параконид сильно выступает вперед. Хищнической вырезки между параконидом и протоконидом нет, на передней стороне протоконида имеется короткое лезвие. Передний край гипоконида находится приблизительно на одной линии с вершиной метаконида (морфотип A2), а вершина гипоконида чуть смещена вперед по сравнению с вершиной мезиального энтоконида. Заднюю часть талонида окаймляют гипоконулид c протяженным постеролингвальным выступом, дистальный энтоконид и один довольно крупный дополнительный талонидный бугорок (морфотип B1).
Рис. 2.
Arctonyx collaris rostratus Matthew et Granger, 1923, экз. ПИН, № 5792/2, фрагмент правой нижнечелюстной кости с M1 с лабиальной (вверху), окклюзиальной (в середине) и лингвальной (внизу) сторон; Вьетнам, провинция Тханьхоа, уезд Батхыок, пещера Лангчанг; верхний плейстоцен.
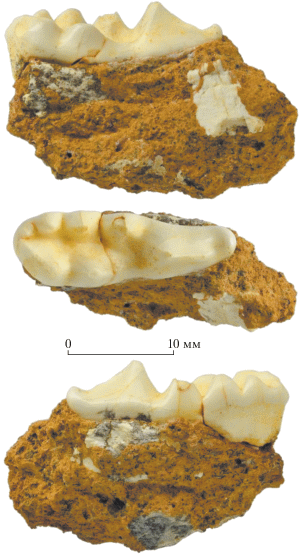
Размеры М1 (экз. ПИН, № 5792/2): L – 21.2, ETRL – 10.0, ITRL – 12.5, TRW – 6.5, TALW – 7.4. Пределы изменчивости размеров М1 для современных представителей вида [1: табл. 3 ]: L – 15.2–20.1 (μL – 18.1), TALW – 6.1–7.7 (μTALW – 6.8). Эти же значения у плейстоценового A. c. rostratus из разных местонахождений Китая [4: табл. 2 ]: L – 16.8–21.0 (μL – 17.9), TALW – 6.5–8.0 (μTALW – 7.0).
Таким образом, по длине M1 (экз. ПИН, № 5792/2) и ширине М1 (экз. ПИН, № 5792/1) форма из пещеры Лангчанг крупнее современных больших свиных барсуков и лишь слегка превосходит известные экземпляры A. c. rostratus. Соответственно, на основании величины и строения зубов она может быть отнесена к подвиду A. c. rostratus.
Ископаемый A. collaris rostratus был первоначально описан из Китая в статусе самостоятельного вида A. rostratus Matthew et Granger, 1923, который считался существенно более крупным, чем A. collaris [7: рис. 12–14 ]. При этом A. rostratus сравнивался с экземплярами из современной популяции, обитающей в центральном Китае (Шэньси), которая в настоящее время относится к отдельному виду северных свиных барсуков, A. albogularis (Blyth, 1853) [1]. Однако кондилобазальная длина черепа A. c. rostratus составляет 148 мм [7], что близко к верхнему пределу A. albogularis (110–149 мм) и даже немного ниже, чем у современного A. collaris (150–174 мм) [1, 2]. При этом A. c. rostratus характеризуется сравнительно крупными и массивными щечными зубами. Ранее ряд находок в плейстоцене Индокитая и Таиланда был идентифицирован как A. collaris [6, 8], A. collaris cf. rostratus [9, 10] и A. c. rostratus [11, 12], поэтому можно полагать, что ареал хронологического подвида A. c. rostratus помимо центральных, восточных и южных частей Китая [4, 13] охватывал и Юго-Восточную Азию.
Для Arctonyx collaris характерна необычно большая для Carnivora внутрипопуляционная изменчивость щечных зубов по размерам и строению [1, 5, 14]. Это явление может быть связано с сильно выраженной у данного вида (даже по сравнению с другими барсуками) вермифагией – питанием земляными червями [1], которые иногда составляют до 70–100% его пищевого рациона [2]. Вермифагия у млекопитающих часто коррелирует с очень высокой морфологической вариабельностью моляров, предположительно отражающей снижение строгости генетического контроля развития этих зубов в связи с ослаблением требований отбора к точности зубной окклюзии [1, 15]. Ископаемые материалы позволяют заключить, что для плейстоценовых популяций A. collaris был характерен в целом не менее высокий уровень морфологической и морфометрической изменчивости коренных зубов.
Список литературы
Helgen K.M., Lim N.T., Helgen L.E. The hog-badger is not an edentate: systematics and evolution of the genus Arctonyx (Mammalia: Mustelidae) // Zoological Journal of the Linnean Society. 2008. V. 154. P. 353–385.
Кузнецов Г.В. Млекопитающие Вьетнама. М.: Т-во научных изданий КМК; 2006.
Proulx G., Abramov A.V., Adams I., et al. World distribution and status of badgers – a review. In: Proulx G., Do Linh San E., editors. Badgers: systematics, biology, conservation and research techniques. Sherwood Park, Alberta, Canada: Alpha Wildlife Publications; 2016. P. 31–116.
Tong H., Wu X., Dong Z., et al. Preliminary report on the mammalian fossils from the ancient human site of Hualong Cave in Dongzhi, Anhui // Acta Anthropologica Sinica. 2018. V. 27. № 2. P. 284–305.
Jiangzuo Q.-G., Liu J.-Y., Wagner J., Chen J. Taxono-mical revision of “Arctonyx” fossil remains from the Liucheng Gigantopithecus Cave (South China) by means of morphotype and morphometrics, and a review of Late Pliocene and Early Pleistocene Meles fossil records in China // Palaeoworld. 2018. V. 27. P. 282–300.
Long V.T., De Vos J., Ciochon R.L. The fossil mammal fauna of the Lang Trang caves, Vietnam, compared with Southeast Asian fossil and recent mammal faunas: the geographical implications // Bulletin of the Indo-Pacific Prehistory Association. 1996. V. 14. P. 101–109.
Matthew W.D., Granger W. New fossil mammals from the Pliocene of Sze-chuan, China // Bulletin of the American Museum of Natural History. 1923. V. 48. P. 563–598.
Suraprasit K., Jaeger J.-J., Chaimanee Y., et al. The Middle Pleistocene vertebrate fauna from Khok Sung (Nakhon Ratchasima, Thailand): biochronological and paleobiogeographical implications // ZooKeys. 2016. V. 613. P. 1–157.
Arambourg C., Fromaget J. Le gisement quaternaire de Tam Nang (Chaîne Annamitique septentrionale). Sa stratigraphique et ses faunes // Comptes Rendus de l’Académie des Sciences. 1938. V. 203. P. 793–795.
Bacon A.-M., Demeter F., Duringer P., et al. The Late Pleistocene Duoi U’Oi cave in northern Vietnam: palaeontology, sedimentology, taphonomy and palaeoenvironments // Quaternary Science Reviews. 2008. V. 27. P. 1627–1654.
Bacon A.-M., Duringer P., Antoine P.-O., et al. The Middle Pleistocene mammalian fauna from Tam Hang karstic deposit, northern Laos: new data and evolutionary hypothesis // Quaternary International. 2011. V. 245. № 2. P. 315–332.
Tougard C. Biogeography and migration routes of large mammal faunas in South–East Asia during the Late Middle Pleistocene: focus on the fossil and extant faunas from Thailand // Palaeogeography, Palaeoclimatology, Palaeoecology. 2001. V. 168. № 3–4. P. 337–358.
Tong H.W. Occurrences of warm-adapted mammals in north China over the Quaternary Period and their paleoenvironmental significance // Science in China. Series D: Earth Sciences. 2007. V. 50. № 9. P. 1327–1340.
Pocock R.I. The fauna of British India, including Ceylon and Burma. Mammalia. V. II: Carnivora [suborders Aeluroida (part) and Arctoida]. London: Taylor and Francis; 1941.
Charles C., Solé F., Gomes Rodrigues H., Viriot L. Under pressure? Dental adaptations to termitophagy and vermivory among mammals // Evolution. 2013. V. 67. № 6. P. 1792–1804.
Дополнительные материалы отсутствуют.
Инструменты
Доклады Российской академии наук. Науки о жизни