

Доклады Российской академии наук. Науки о жизни, 2021, T. 497, № 1, стр. 125-130
АНОМАЛИИ ЗУБНОЙ СИСТЕМЫ У NESOPHONTIDAE (LIPOTYPHLA, MAMMALIA) ИЗ ПЛЕЙСТОЦЕНА КУБЫ
Академик РАН А. В. Лопатин *
Палеонтологический институт им. А.А. Борисяка Российской академии наук
Москва, Россия
* E-mail: alopat@paleo.ru
Поступила в редакцию 01.12.2020
После доработки 10.12.2020
Принята к публикации 10.12.2020
Аннотация
Изучение остатков Nesophontes micrus Allen, 1917 и N. major Arredondo, 1970 из пещеры Эль-Аброн (Республика Куба, провинция Пинар-дель-Рио, поздний плейстоцен) позволило впервые отметить аномалии зубной системы Nesophontidae. Нарушения прорезывания зубов (импакция I3 у N. micrus, ретенция или импакция P2 у N. major), как и мальформации (аберрантные I3 у N. micrus, P2 и P4 у N. major), являются аномалиями развития. Отсутствие у N. major P2 и M2 при реоссификации их альвеол обусловлено травмами и заболеваниями зубов.
Из плейстоцена и голоцена Кубы известны ископаемые и субфоссильные остатки двух видов насекомоядных вымершего эндемичного семейства Nesophontidae – более мелкого Nesophontes micrus Allen, 1917 и более крупного N. major Arredondo, 1970 [1, 2]. Таксономическая самостоятельность этих видов согласуется с результатами протеомного анализа [3].
Оба кубинских вида незофонтов известны по многочисленным экземплярам из ряда местонахождений [4]. Массовые материалы делают возможным изучение внутривидовой изменчивости морфологических признаков, включая различные отклонения. Из верхнеплейстоценовых отложений пещеры Эль-Аброн (провинция Пинар-дель-Рио, координаты 22°40' с.ш., 83°28' з.д.) в 2019 г. Совместной российско-кубинской палеонтологической экспедицией (СРКПЭ) были добыты около 20 черепных фрагментов и 100 нижнечелюстных костей N. micrus и более 40 черепных фрагментов и 200 нижнечелюстных костей N. major. Изученные материалы происходят из слоя VII с радиоуглеродной датировкой 17406 ± 161 лет (калиброванный возраст 20 050–21 474 лет) [5], а также из нижележащих слоев IX и VIII и вышележащих слоев VI и V. В результате изучения изменчивости морфологии среди них обнаружены несколько экземпляров нижнечелюстных костей с аномалиями зубной системы: N. major, экз. ПИН, № 5807/251 из слоя IX (глубина 2.93 м) и экз. ПИН, №№ 5807/66, 68, 76 из слоя VII (без указания глубины); N. micrus, экз. ПИН, № 5807/173 из слоя VII (без указания глубины).
При исследовании выявленных аномалий зубной системы учитывались новейшие данные о смене зубов у Nesophontidae, включая сведения о смене всех нижних резцов, клыка и премоляров P2 и P4 при зубной формуле I1–3C1P1–2,4M1–3 [6].
Изученный материал хранится в Палеонтологическом институте им. А.А. Борисяка РАН (ПИН) в Москве. Фотографии сделаны с помощью сканирующего электронного микроскопа Tescan Vega II XMU, рентгенографическое и томографическое исследование проведено на рентгеновском микротомографе SkyScan 1172 в ПИН.
На нижнечелюстной кости Nesophontes micrus (экз. ПИН, № 5807/173) перед обломанным клыком C1 имеется полуоткрытая альвеола с непрорезавшимся последним резцом I3 (рис. 1). Резец заметно скошен, его альвеола ориентирована дорсолабиально. Отверстие альвеолы овальной формы, смещено на лабиальную сторону и расположено заметно вентральнее альвеолярного края горизонтальной ветви. На рентгенограмме видно, что в этой альвеоле находится аномально мелкий нефункциональный зуб с редуцированной коронкой, узкой пульпарной полостью и нормальным корневым каналом, сообщающимся с нижнечелюстным каналом (рис. 1д). На слегка выступающей из альвеолы части коронки зуба повреждений не отмечено. Очевидно, при жизни животного I3 был полностью скрыт десной. Непосредственно перед клыком поверхность кости покрыта мелкими неровностями и углублениями, видимо, связанными с регенерацией костной ткани. Эта часть кости нависает над вершиной резца с лингвальной стороны. Скорее всего, данное состояние связано с отклонением развития (мальформацией) и нарушением прорезывания постоянного резца I3 при частичной наружной облитерации альвеолы молочного резца DI3.
Рис. 1.
Nesophontes micrus Allen, 1917, экз. ПИН, № 5807/173, правая нижнечелюстная кость с I3–C1 и M1–M3: а – с лабиальной стороны, б – с окклюзиальной стороны, в – передняя часть с лабиальной стороны, г – передняя часть с окклюзиальной стороны, д – рентгенограмма передней части с лингвальной стороны; Куба, пещера Эль-Аброн (слой VII); верхний плейстоцен.
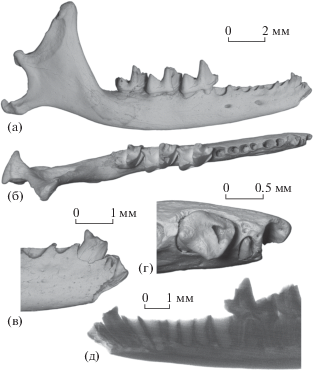
Патологическое нарушение (задержка и остановка) процесса прорезывания зубов называется ретенцией. Нарушение рассматривается как импакция, если оно обусловлено наличием препятствия для непрорезавшегося зуба в виде другого зуба или нерезорбированной костной ткани [7]. В описанном случае импакция I3 могла быть связана с ранней потерей его молочного предшественника (DI3) в результате травмы или заболевания и последующим сближением соседних зубов (DI2 и DC1) друг с другом [см. 7, с. 348] при частичном зарастании отверстия альвеолы DI3. Это смещение сильно сократило межзубной промежуток на месте выпавшего DI3 и воспрепятствовало нормальному формированию и прорезыванию I3 [см. 6].
Судя по слабо стертым молярам с узкими фасетками стирания на гребнях, данная нижнечелюстная кость принадлежала взрослой особи сравнительно молодого возраста. Размеры экз. ПИН, № 5807/173 (в мм): длина M1–M3 – 5.9; моляры (длина × ширина): M1 – 2.1 × 1.4; M2 – 2.1 × 1.4; M3 – 1.9 × 1.2; высота горизонтальной ветви нижней челюсти под M2/M3 – 2.2; высота венечного отростка – 5.5; ширина суставного мыщелка – 2.2.
Нижнечелюстная кость Nesophontes major, экз. ПИН, № 5807/76, несет аномально мелкий предпоследний премоляр P2 (рис. 2). Его корни короткие, а коронка очень маленькая, слабо удлиненная и не полностью выступает над альвеолярным краем горизонтальной ветви. На задней и лингвальной сторонах P2 хорошо выражен цингулид. Костные края альвеолы немного нависают над цингулидом P2, это хорошо заметно с окклюзиальной и лингвальной сторон горизонтальной ветви (рис. 2б, 2д). Альвеолы корней последнего премоляра P4 крупные. Вероятно, описанное состояние P2 представляет собой аномалию развития (мальформацию), связанную с задержкой прорезывания (либо ретенцией неясной природы, либо импакцией из-за приближенных к P2 соседних крупных премоляров, P1 и P4) [см. 7, рис. 11.55 ]. Нижнечелюстная кость принадлежала полностью взрослой или сенильной (стареющей) особи, так как сохранившиеся моляры (M1 и M2) довольно сильно стерты, фасетки стирания гребней на них широкие, слитые между собой. Размеры нижней челюсти и зубов N. major приведены в табл. 1.
Рис. 2.
Nesophontes major Arredondo, 1970, экз. ПИН, № 5807/76, левая нижнечелюстная кость с P2 и M1–M2: а – с лабиальной стороны, б – с окклюзиальной стороны, в – передняя часть с окклюзиальной стороны, г – передняя часть с лабиальной стороны, д – передняя часть с лингвальной стороны, е – рентгенограмма с лабиальной стороны; Куба, пещера Эль-Аброн (слой VII); верхний плейстоцен.

Таблица 1.
Размеры (в мм) нижней челюсти и зубов Nesophontes major Arredondo, 1970 из верхнего плейстоцена пещеры Эль-Аброн, Куба (обозначения: AL – альвеолярная длина; L – наибольшая длина; W – наибольшая ширина; D – высота горизонтальной ветви под M2/M3; HCP – высота венечного отростка; WAC – ширина суставного мыщелка)
| Экземпляр ПИН, № | Длина зубных рядов и альвеол | Размеры зубов (L × W) | D | HCP | WAC |
|---|---|---|---|---|---|
| 5807/66 | L M1–M3: 6.4 | P4: 1.3 × 0.85; M1: 2.3 × 1.65; M2: 2.3 × 1.7; M3: 2.1 × 1.35 | 2.7 | 6.2 | 2.8 |
| 5807/68 | – | P4: 1.6 × 0.9 | 2.8 | 6.4 | 2.7 |
| 5807/76 | AL M1–M3: 5.7 | P2: ~ 0.8 × 0.6; M1: 2.2 × 1.7; M2: 2.3 × 1.8 | 3.0 | 7.2 | 2.8 |
| 5807/251 | L P4–M3 – 7.3; L M1–M3 – 5.9 |
P1: 2.1 × 0.95; P4: 1.6 × 1.0; M1: 2.2 × 1.7; M2: 2.1 × 1.7; M3: 1.9 × 1.5 | 2.8 | – | – |
По длине P2 (около 0.8 мм, эмаль в передней части разрушена) экз. ПИН, № 5807/76 сильно уступает другим экземплярам N. major (длина 1.23–1.8 мм, в среднем 1.5 мм, n = 26) [2]. Наличие аномально мелких премоляров отмечено у современных кротовых [8, рис. 9 ].
Нижнечелюстная кость N. major, экз. ПИН, № 5807/66, содержит целые P4–M3 и альвеолы I1–P2 (рис. 3). Моляры слабо стертые, фасетки стирания гребней узкие. Зуб в локусе последнего премоляра имеет аберрантное строение, которое заметно отличается от типичного для P4N. major (см. рис. 4). Он характеризуется тесно прижатыми друг к другу корнями, высоким и коротким протоконидом с вертикальной задней стенкой, маленьким параконидом и очень коротким и низким талонидом. От M1 этот зуб отделен заметным промежутком, что также нехарактерно для N. major (см. рис. 4). Рентгенографическое и томографическое изучение образца позволило выявить длинные корни P4 и отсутствие сменяющих зубов в альвеолах локусов I1–P4 (рис. 3в). Таким образом, описываемый зуб – это постоянный последний премоляр [см. 6], демонстрирующий отклонение в строении коронки (мальформацию). Альвеолярный край кости непосредственно перед M1 резорбирован, что может указывать на наличие патологического процесса, с которым допустимо связать мальформацию P4.
Рис. 3.
Nesophontes major Arredondo, 1970, экз. ПИН, № 5807/66, правая нижнечелюстная кость с P4–M3: а – с лабиальной стороны, б – с окклюзиальной стороны, в – рентгенограмма с лабиальной стороны; Куба, пещера Эль-Аброн (слой VII); верхний плейстоцен.
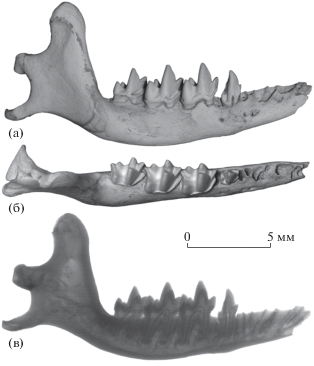
Рис. 4.
Nesophontes major Arredondo, 1970: а–в – экз. ПИН, № 5807/251, левая нижнечелюстная кость с P1, P4–M3 и заросшими альвеолами P2: а – с лабиальной стороны, б – с окклюзиальной стороны, в – рентгенограмма с лабиальной стороны; г–ж – экз. ПИН, № 5807/68, правая нижнечелюстная кость с P4, альвеолами C1–P2, корнями M1, M3 и заросшими альвеолами M2: г – с лабиальной стороны, д – с окклюзиальной стороны, е – область моляров с окклюзиальной стороны с лингвальным наклоном, ж – рентгенограмма с лингвальной стороны; Куба, пещера Эль-Аброн (а–в – слой IX; г–ж – слой VII); верхний плейстоцен.
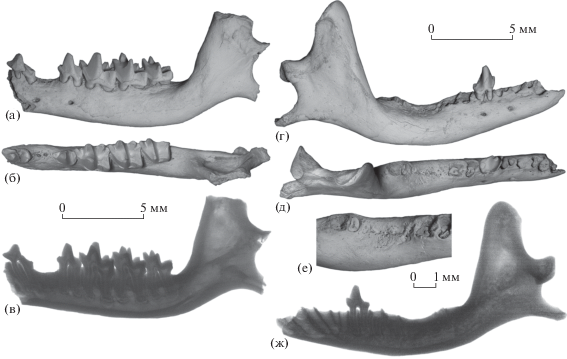
На нижнечелюстных костях двух других особей N. major (рис. 4) имеются свидетельства прижизненной утраты щечных зубов и наружного зарастания их альвеол. В одном случае утрачен P2 (экз. ПИН, № 5807/251), в другом – M2 (экз. ПИН, № 5807/68).
Экз. ПИН, № 5807/251 с умеренно стертыми молярами обнаруживает аномальное отсутствие предпоследнего премоляра P2 (рис. 4а, 4б). В промежутке между P1 и P4 хорошо заметны две крохотные поверхностные ямки на месте альвеол корней утраченного P2. Альвеолярный край в области межзубного промежутка немного прогнут. Рентгенографическое исследование позволило установить, что альвеолы изолированы от нижнечелюстного канала и заполнены пористой костной тканью (рис. 4в). Это свидетельствует о полном зарастании альвеол после выпадения P2 [см. 9, 10]. Соответственно, причиной отсутствия зуба в данном случае является травма или заболевание, а не генетически обусловленный дефект развития, приводящий к гиподонтии (олигодонтии). Заболевания и залеченные травмы нижней челюсти, сопровождающиеся утратой премоляров и моляров, описаны у современных землероек [11].
Экз. ПИН, № 5807/68 демонстрирует аномальное отсутствие предпоследнего моляра и патологическое деградированное состояние зубной ветви в области M2 (рис. 4г–4е) – сильный прогиб ее альвеолярной стороны [см. 7, рис. 24.50 ; 12, рис. 1, J]. В локусе M2 между альвеолами M1 и M3 отсутствуют отверстия альвеол для корней зуба, а на их месте в пределах прогнутой части челюсти находится продольно вытянутое углубление с многочисленными мелкими ямками и порами. Рентгенографическое исследование (рис. 4ж) показало наличие внутри кости заросших альвеол, заполненных пористой костной тканью и капсулированных как с наружной стороны, так и со стороны нижнечелюстного канала. Можно заключить, что видимое отсутствие альвеол M2 на экз. ПИН, № 5807/68 обусловлено их реоссификацией и наружной облитерацией после прижизненной утраты разрушенного или больного зуба [см. 13, 14]. Деградация костной ткани с сильным уменьшением высоты зубной ветви указывает на осложненный периодонтит как причину утраты зуба [7].
Таким образом, исследование массовых остатков Nesophontes micrus и N. major из пещеры Эль-Аброн позволило впервые выявить аномалии зубной системы Nesophontidae, включая нарушения прорезывания и отклонения в развитии резцов и премоляров, а также связанное с травмами и заболеваниями патологическое отсутствие премоляров и моляров при реоссификации их альвеол.
БЛАГОДАРНОСТИ
Автор благодарит доктора Ф.М. Гонсалеса Бермудеса и К.А. Мендеса Гарсия (Министерство науки, технологии и окружающей среды Республики Куба, CITMA) за организационную помощь и дружескую поддержку, Э. Перес Лоренсо (Национальный музей естественной истории Кубы), Н.В. Зеленкова, А.К. Агаджаняна (ПИН) и всех сотрудников СРКПЭ – за участие в экспедиции 2019 г., А.В. Пахневича и Р.А. Ракитова (ПИН) – за техническую помощь в проведении томографии и подготовке фотографий на СЭМ.
Список литературы
Condis Fernández M.М., Jiménez Vásquez O., Arredondo C. Revisión taxonómica del género Nesophontes (Insecti-vora: Nesophontidae) en Cuba: análisis de los caracteres diagnóstico // Monografies de la Societat d’Història Natural de les Balears. 2005. № 12. P. 95–100.
Rzebik-Kowalska B., Wołoszyn B.W. New data on Nesophontes subfossil populations from Cuba and Isla de la Juventud (Cuba) // Neues Jahrbuch für Geologie und Paläontologie Abhandlungen. 2012. V. 263. № 2. P. 155–166.
Buckley M., Harvey V.L., Orihuela J., et al. Collagen sequence analysis reveals evolutionary history of extinct West Indies Nesophontes (island-shrews) // Molecular Biology and Evolution. 2020. V. 37. № 10. P. 2931–2943.
Silva Taboada G., Suárez Duque W., Díaz Franco S. Compendio de los mamíferos terrestres autóctonos de Cuba vivientes y extinguidos. La Habana: Museo Nacional de Historia Natural, 2007.
Suárez W., Díaz-Franco S. A new fossil bat (Chiroptera: Phyllostomidae) from a Quaternary cave deposit in Cuba // Caribbean Journal of Science. 2003. V. 39. № 3. P. 371–377.
Лопатин А.В. Смена зубов у Nesophontidae (Lipotyphla, Mammalia) из плейстоцена Кубы // Доклады Российской Академии наук. Науки о жизни. 2021. Т. 497. С. 118–124.
Miles A.E.W., Grigson C. Colyer’s variations and diseases of the teeth of animals. Cambridge: Cambridge University Press, 1990.
Asahara M., Kryukov A., Motokawa M. Dental anomalies in the Japanese mole Mogera wogura from northeast China and the Primorsky region of Russia // Acta Theriologica. 2012. V. 57. № 1. P. 41–48.
Wilson R.W. Two cases of dental anomaly in early Tertiary mammals // Transactions of the Kansas Academy of Science. 1955. V. 58. № 4. P. 514–518.
Лопатин А.В. Древнейший Hapalodectes (Mesonychia, Mammalia) из палеоцена Монголии // Палеонтологический журнал. 2001. № 4. С. 90–96.
Jentzsch M., Kraft R., Lemkul A., et al. Anomalies and pathological changes of skulls and dentition of wild small mammal species from Germany // Journal of Vertebrate Biology. 2020. V. 69. № 4. P. 1–19.
Powers P.M., Mead A.J. Dental and mandibular anomalies in white-tailed deer (Odocoileus virginianus) from Central Georgia // Georgia Journal of Science. 2019. V. 77. № 2. Art. 15. P. 1–11.
Vilà C., Urios V., Castroviejo J. Tooth losses and anomalies in the wolf (Canis lupus) // Canadian Journal of Zoology. 1993. V. 71. № 5. P. 968–971.
Palmqvist P., Arribas A., Martínez-Navarro B. Ecomorphological study of large canids from the Lower Pleistocene of southeastern Spain // Lethaia. 1999. V. 32. № 1. P. 75–88.
Дополнительные материалы отсутствуют.
Инструменты
Доклады Российской академии наук. Науки о жизни