

Доклады Российской академии наук. Науки о жизни, 2021, T. 497, № 1, стр. 118-124
СМЕНА ЗУБОВ У NESOPHONTIDAE (LIPOTYPHLA, MAMMALIA) ИЗ ПЛЕЙСТОЦЕНА КУБЫ
Академик РАН А. В. Лопатин *
Палеонтологический институт им. А.А. Борисяка Российской академии наук
Москва, Россия
* E-mail: alopat@paleo.ru
Поступила в редакцию 11.11.2020
После доработки 21.11.2020
Принята к публикации 22.11.2020
Аннотация
На материалах по Nesophontes major Arredondo, 1970 и N. micrus Allen, 1917 из пещеры Эль-Аброн (Республика Куба, провинция Пинар-дель-Рио, поздний плейстоцен) впервые для Nesophontidae обнаружено наличие молочных премоляров и установлена смена C1, P2, P4, I1, I2, I3, P2 и P4. В онтогенезе смена зубов (кроме P1 и P1) происходила после прорезывания моляров, но до достижения животным взрослых размеров. По характеру процесса зубной смены незофонтиды отличаются от большинства представителей Lipotyphla и сближаются с Solenodontidae.
Незофонты (Nesophontidae) известны в ископаемой летописи островов Карибского региона с позднего плейстоцена до позднего голоцена. Последние представители этого эндемичного семейства насекомоядных вымерли около 500 лет назад [1, 2].
В настоящее время морфологически хорошо обоснована валидность семи видов Nesophontes: N. edithae Anthony, 1916 (Пуэрто-Рико, Вьекес, Сент-Джон, Сент-Томас), N. zamicrus Miller, 1929, N. paramicrus Miller, 1929 (Гаити), N. hypomicrus Miller, 1929 (Гаити, Гонав), N. major Arredondo, 1970 (Куба), N. micrus Allen, 1917 (Куба, Хувентуд) и N. hemicungulus Morgan et al., 2019 (Кайман-Брак, Большой Кайман) [3–5]. Эти выводы согласуются с новейшими результатами протеомного анализа [6] и молекулярно-генетических исследований [7].
Незофонты известны по многочисленным ископаемым и субфоссильным остаткам из пещерных местонахождений. В некоторых местах найдены остатки сотен особей, включая многочисленные черепные и нижнечелюстные фрагменты с зубами [4, 8]. При этом некоторые характеристики зубной системы незофонтов остаются плохо изученными. В частности, до настоящего времени имелось очень мало сведений о смене и порядке прорезывания зубов у незофонтов [4, 9, 10], полностью отсутствовали данные о молочных премолярах.
Г. Энтони [9, с. 74] отметил наличие субпремоляриформного молочного нижнего клыка (DC1) у молодой половозрелой особи N. edithae. У этого экземпляра в одной альвеоле с DC1 находится постоянный клык (C1), вершина которого видна у основания молочного клыка. DC1 существенно ниже C1, сильнее наклонен вперед и имеет лучше развитый задний выступ. Энтони полагал [9, с. 90], что на этой возрастной стадии функционировали молочные резцы.
С. Макдауэлл [10, с. 160] c помощью рентгенографии обнаружил у изученной Энтони молодой особи N. edithae эмалевый колпачок зачаточного второго резца (I2) в глубине альвеолы DI2, но не отметил каких-либо следов DI1, DI3 и молочных премоляров. Макдауэлл полагал, что столь скудная представленность молочных зубов в весьма обильных по числу особей материалах может быть связана с тем, что молочные зубы незофонтов выпадали очень рано в онтогенезе – еще до того, как зверьки начинали активно самостоятельно питаться на поверхности земли, становясь добычей хищных птиц и попадая в пещерные тафоценозы [10, с. 149]. Для молочных премоляров он рассматривал также альтернативную возможность – предполагал, что именно молочная смена функционировала в течение жизни животных, тогда как постоянная смена не проявлялась вовсе [10, с. 150, 160]. В итоге Макдауэлл предложил следующую последовательность прорезывания нижних зубов у Nesophontes: -DI2-DC1----M1M2M3I1I2I3C1P1P2-P4 (при зубной формуле I1–3/1–3C1/1P1–2,4/1–2,4M1–3/1–3), и заключил, что наличие молочной смены зубов (по меньшей мере, DI2 и DC1) резко отличает незофонтид от землеройковых (Soricidae) [10, с. 160].
Б. Ржебик-Ковальская и Б. Волошин [4, с. 159, рис. 3H, табл. 4 ] отметили DC1 у двух субфоссильных молодых экземпляров N. major (Куба, Хувентуд; минимальное число особей в выборке – 102). Они показали, что у этого вида DC1 также отличается от C1 меньшими размерами (включая высоту) и субпремоляриформным строением коронки.
Для исследования смены зубов у незофонтов необходимо изучение массовых материалов при целенаправленном внимании к остаткам наиболее молодых особей.
В настоящей статье приведены результаты исследования наиболее полных образцов Nesophontes major и N. micrus из верхнеплейстоценовых отложений пещеры Эль-Аброн в горах Сьерра-де-ла-Гуира (Sierra de La Güira), муниципалитет Лос-Паласиос (Los Palacios), провинция Пинар-дель-Рио Республики Куба (координаты 22°40' с.ш., 83°28' з.д.) [11]. В ноябре 2019 г. здесь проводила сборы ископаемых остатков Совместная российско-кубинская палеонтологическая экспедиция (СРКПЭ). Основной изученный материал происходит из слоя (уровня) VII (радиоуглеродная датировка 17406±161 лет, калиброванный возраст 20050–21474 лет; см. [11]), дополнительно изучены образцы из нижележащих слоев IX и VIII и вышележащих слоев VI и V.
Материал хранится в Палеонтологическом институте им. А.А. Борисяка РАН (ПИН) в Москве. Фотографии сделаны с помощью сканирующего электронного микроскопа Tescan Vega II XMU, рентгенографическое и томографическое исследование проведено на рентгеновском микротомографе SkyScan 1172 в ПИН. Измерения в мм.
Всего изучено более 40 черепных фрагментов и свыше 200 нижнечелюстных костей N. major, а также около 20 черепных фрагментов и более 100 нижнечелюстных костей N. micrus. Представлены экземпляры с C1, P1, P2, P4, M1, M2, M3, I1, I2, I3, C1, P1, DP2, P2, DP4, P4, M1, M2, M3 (включая C1, P2, I1, I2, I3, C1, P2, P4 в состоянии прорезывания). В их числе экз. ПИН, №№ 5807/253, 254 из слоя VIII (глубина 2.35 м), экз. ПИН, № 5807/255 из слоя VII (глубина 1.5–1.54 м), экз. ПИН, №№ 5807/261–266 из слоя VII (глубина 1.2–1.5 м), экз. ПИН, № 5807/260 из слоя VII (глубина 1.25 м), экз. ПИН, №№ 5807/67, 71, 75, 77, 78, 267 из слоя VII (без указания глубины), экз. ПИН, №№ 5807/269–273 из слоя VI (без указания глубины), экз. ПИН, №№ 5807/275–277 из слоя V (глубина 0.45–0.5 м).
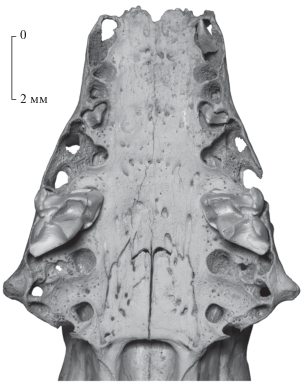
На фрагментарном черепе Nesophontes major (экз. ПИН, № 5807/269) впервые обнаружены свидетельства смены верхних зубов в локусах C1, P2 и P4 (рис. 1). Левый C1 и оба P2 не прорезались из альвеол, их вершины лишь немного выступают за альвеолярный край верхнечелюстных костей. Основание и корни клыка не сформированы, межкорневой перегородки нет, коронка развернута в альвеоле (передняя сторона обращена лингвально) и наклонена вперед. Передняя и антеролингвальная бороздки C1 (предположительно, служившие для стока яда, см. [12]) хорошо выражены. Альвеолы P1 с межкорневой перегородкой без следов резорбции, что указывает на отсутствие смены зубов в этом локусе. P2 одновершинный, вершина слегка загнута назад, корни короткие. Межкорневая перегородка альвеолы DP2/P2 резорбирована. Оба P4 не сохранились, но их альвеолы широко раскрыты и не имеют межкорневых перегородок, что указывает на недавнюю (или неоконченную) смену зубов этого локуса. M1 апикально стертые, с резкими, едва стертыми гребнями. Альвеолярная длина C1–M3 – 11.5 (левые), 11.3 (правые). Размеры M1 (длина × × ширина): 2.4 × 2.9 (левый, правый).
Рис. 1.
Nesophontes major Arredondo, 1970, экз. ПИН, № 5807/269, фрагмент черепа молодой особи, передняя часть с вентральной стороны; Куба, пещера Эль-Аброн (слой VI); верхний плейстоцен.
Среди материалов по N. major представлены нижнечелюстные кости с I3, C1, P2 и P4 на разных стадиях прорезывания, а также фрагмент нижней челюсти молодой особи (экз. ПИН, № 5807/254) с несформированными краями альвеол M1–M3 (табл. 1).
Таблица 1.
Размеры (в мм) нижней челюсти и зубов Nesophontes major Arredondo, 1970 из верхнего плейстоцена пещеры Эль-Аброн, Куба (обозначения: AL – альвеолярная длина; L – наибольшая длина; W – наибольшая ширина; D – высота горизонтальной ветви под M2/M3; HCP – высота венечного отростка; WAC – ширина суставного мыщелка)
| Экземпляр ПИН, № | Длина зубных рядов и альвеол | Размеры зубов (L × W) | D | HCP | WAC |
|---|---|---|---|---|---|
| 5807/67 | L M1–M3: 6.6 | M1: 2.3 × 1.65; M2: 2.3 × 1.7; M3: 2.1 × 1.35 | 2.8 | – | 2.7 |
| 5807/71 | AL M1–M3: 5.0 | M3: 2.0 × 1.4 | 2.6 | – | 2.6 |
| 5807/75 | L M1–M3: 6.6 | M1: 2.4 × 1.6; M2: 2.3 × 1.7; M3: 2.2 × 1.45 | 2.7 | 6.4 | 2.6 |
| 5807/77 | L M1–M3: 6.3 | M1: 2.2 × 1.55; M2: 2.2 × 1.6; M3: 2.0 × 1.35 | 2.7 | – | 2.5 |
| 5807/78 | AL M1–M3: 6.0 | M3: 2.2 × 1.45 | 3.1 | – | 2.45 |
| 5807/253 | L M1–M3: 5.9 | M1: 2.1 × 1.4; M2: 2.1 × 1.5; M3: 1.9 × 1.3 | 2.6 | 6.7 | 2.7 |
| 5807/254 | AL M1–M3: 5.9 | – | 2.6 | 6.0 | 2.3 |
| 5807/261 | AL M1: 2.0 | – | 2.8 | – | – |
| 5807/262 | L M1–M3: 6.7 | P1: 2.0 × 0.8; M1: 2.4 × 1.55; M2: 2.4 × 1.7; M3: 2.1 × 1.45 | 2.8 | 6.5 | 2.7 |
| 5807/263 | L M1–M3: 6.3 | P1: 2.0 × 0.8; M1: 2.3 × 1.5; M2: 2.3 × 1.6; M3: 1.9 × 1.4 | 2.8 | 7.0 | 3.0 |
| 5807/264 | AL M1–M3: 6.3 | – | 3.0 | 6.4 | 2.4 |
| 5807/270 | L M1–M3: 6.4 | M1: 2.2 × 1.5; M2: 2.2 × 1.6; M3: 2.05 × 1.4 | 2.5 | 6.6 | 2.4 |
| 5807/271 | L M1–M3: 6.0 | M1: 2.2 × 1.5; M2: 2.3 × 1.55; M3: 1.9 × 1.3 | 2.6 | – | – |
| 5807/275 | L M1–M3: 6.15 | P1: 1.9 × 0.8; M1: 2.2 × 1.5; M2: 2.2 × 1.6; M3: 1.95 × 1.4 | 2.6 | 6.3 | – |
| 5807/276 | L M1–M3: 6.0 | M3: 1.9 × 1.4 | 2.6 | 6.2 | 2.5 |
Сохранившийся I3 (экз. ПИН, № 5807/271; рис. 2а–2в) смещен из своей альвеолы в альвеолу DC1/C1 (из-за резорбции и частичного разрушения вентральной части межальвеолярной стенки DI3/DC1) и налегает сверху на клык C1.
Рис. 2.
Nesophontes major Arredondo, 1970: а–в – экз. ПИН, № 5807/271, левая нижнечелюстная кость с фрагментом DP2, M1–M3 и непрорезавшимися I3, C1, P2 и P4: а – с лабиальной стороны, б – с окклюзиальной стороны, в – передняя часть с окклюзиальной стороны с лингвальным наклоном; г–е – экз. ПИН, № 5807/262, левая нижнечелюстная кость с P1, M1–M3 и непрорезавшимися P2 и P4 в альвеолах: г – с окклюзиальной стороны, д – передняя часть с окклюзиальной стороны, е – рентгенограмма с лабиальной стороны; ж – экз. ПИН, № 5807/71, передняя часть левой нижнечелюстной кости с непрорезавшимся Р4 с окклюзиальной стороны; Куба, пещера Эль-Аброн (а–в – слой VI; г–ж – слой VII); верхний плейстоцен.
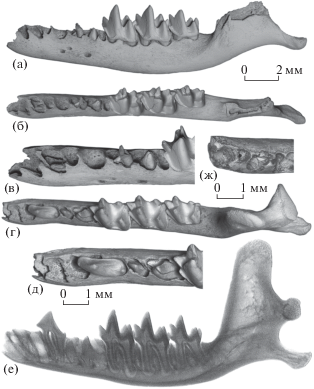
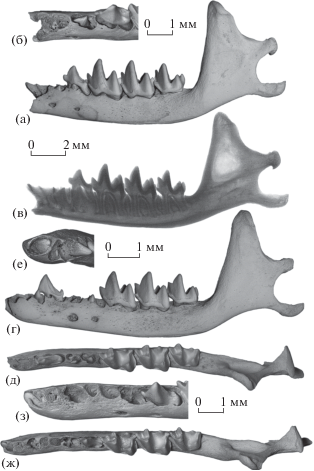
Рис. 3.
Nesophontes micrus Allen, 1917: а–в – экз. ПИН, № 5807/260, левая нижнечелюстная кость с DP4–M3 и непрорезавшимися P2 и P4: а – с лабиальной стороны, б – передняя часть с окклюзиальной стороны, в – рентгенограмма с лабиальной стороны; г–е – экз. ПИН, № 5807/272, левая нижнечелюстная кость c P1 и M1–M3, с непрорезавшимися I1, C1, P2 и P4: г – с лабиальной стороны, д – с окклюзиальной стороны, е – передняя часть с передней стороны; ж, з – экз. ПИН, № 5807/267, левая нижнечелюстная кость с M1–M3 и полупрорезавшимися I1 и P4: ж – с окклюзиальной стороны, з – передняя часть с окклюзиальной стороны с лингвальным наклоном; Куба, пещера Эль-Аброн (а–в, ж, з – слой VII; г–е – слой VI); верхний плейстоцен.
На наиболее ранней внутриальвеолярной стадии C1 лежит в основании альвеолы DC1 и не виден с боковых сторон зубной кости (экз. ПИН, №№ 5807/261, 271; рис. 2а–2в). Судя по рентгенографическим наблюдениям, минерализована лишь верхняя часть коронки этого зуба. На следующей стадии клык имеет сформированное основание коронки с задним выступом и короткий корень, вершина коронки слегка возвышается над альвеолярным краем зубной кости (экз. ПИН, №№ 5807/67, 270).
У большинства экземпляров с непрорезавшимися зубами (например, экз. ПИН, №№ 5807/262, 263; рис. 2г–2е) P1 уже полностью функциональный (хотя и не стертый) и имеет альвеолу с полной (нерезорбированной) межкорневой перегородкой; лишь на экз. ПИН, № 5807/78 межкорневая перегородка неполная, что указывает либо на недавнее прорезывание P1, либо на особенность его строения – сильно сближенные корни. Видимо, передние премоляры (P1 и P1) незофонтид не имели смены (как у большинства плацентарных) и прорезывались в одно время с молярами.
На ранней стадии P2 и P4 были полностью скрыты в альвеолах, а функционировали DP2 и DP4. От DP2 сохранился только короткий задний корень, под которым находится талонид скрытого в альвеоле P2 (экз. ПИН, № 5807/271; рис. 2в). Альвеолы переднего и заднего корней молочных премоляров широко расставлены, а вентральнее DP2 и DP4 располагаются минерализованные коронки сменяющих зубов. На экз. ПИН, № 5807/71 в сломе стенки альвеолы утраченного DP4 виден участок эмали находящегося внутри челюсти зуба, который еще не прорезался с окклюзиальной стороны (рис. 2ж). Рентгенографическое и томографическое изучение показало наличие в этой области крупной полости c коронкой P4 в стадии формирования.
На стадии резорбции межкорневых перегородок и раскрытия альвеол P2 и P4 имеют полностью сформированные коронки с параконидом и талонидом и короткие корни (экз. ПИН, №№ 5807/78, 261, 262, 264, 271; рис. 2е). Широкое раскрытие и частичная резорбция краев и перегородок альвеол P2 и P4, свидетельствующие о незавершенном процессе прорезывания зубов этих локусов, фиксируется у ряда экземпляров с апикально стертыми молярами, у которых P2–P4 не сохранились (экз. ПИН, №№ 5807/67, 75, 77, 253, 270, 275).
На следующем этапе прорезывания P4 приблизительно на треть высоты главного бугорка возвышается над альвеолярным краем зубной кости, тогда как P2 полностью скрыт в альвеоле (экз. ПИН, № 5807/276). У P4, прорезавшегося над альвеолярным краем зубной кости примерно на половину высоты главного бугорка, корни приблизительно вдвое уступают по длине корням моляров (экз. ПИН, № 5807/263).
Среди остатков Nesophontes micrus представлены нижнечелюстные кости с резцами (I1, I2, I3), клыком (C1) и премолярами (P2, P4) на разных стадиях прорезывания (табл. 2), а также один образец (экз. ПИН, № 5807/260) с DP4 – это первая находка сохранившегося молочного предкоренного зуба у Nesophontidae, если не считать описанный выше фрагмент DP2N. major. DP4 имеет широко расставленные корни (рис. 3а–3в) и характеризуется сравнительно низким и узким протоконидом и широким двухбугорковым талонидом (рис. 3б). Альвеолы P2 и P4 на начальной стадии раскрытия, над альвеолярным краем зубной кости возвышаются только вершины главных бугорков этих зубов, протоконид P4 упирается в вентральную часть основания DP4. Рентгенография показала наличие сформированных коротких корней у P2 и P4 (рис. 3в).
Таблица 2.
Размеры (в мм) нижней челюсти и зубов Nesophontes micrus Allen, 1917 из верхнего плейстоцена пещеры Эль-Аброн, Куба (обозначения см. табл. 1)
| Экземпляр ПИН, № | Длина зубных рядов и альвеол | Размеры зубов (L × W) | D | HCP | WAC |
|---|---|---|---|---|---|
| 5807/255 | AL M1: 1.7; AL M2: 1.8 | – | 2.2 | – | – |
| 5807/260 | L M1–M3: 6.0 | DP4: 1.6 × 0.75; M1: 2.15 × 1.5; M2: 2.05 × 1.5; M3: 1.9 × 1.2 | 2.2 | 5.7 | 2.15 |
| 5807/265 | AL M1–M3: 5.7 | – | 2.0 | 5.5 | 2.0 |
| 5807/266 | AL M1–M3: 5.7 | – | 2.2 | – | 2.0 |
| 5807/267 | L M1–M3: 5.9 | M1: 2.1 × 1.4; M2: 2.0 × 1.45; M3: 1.9 × 1.2 | 2.1 | 5.9 | – |
| 5807/272 | L I1–M3: ~11.7; L M1–M3: 5.9 | P1: 1.6 × 0.6; M1: 2.2 × 1.3; M2: 2.1 × 1.4; M3: 1.9 × 1.15 | 2.0 | 5.7 | 2.0 |
| 5807/273 | L M1–M3: 5.9 | M1: 2.2 × 1.4; M2: 2.1 × 1.45; M3: 1.85 × 1.2 | 2.0 | 5.65 | 2.1 |
| 5807/277 | AL M1–M3: 5.0; L M2–M3: 3.9 | M2: 2.2 × 1.4; M3: 1.9 × 1.2 | 2.0 | 5.1 | 1.7 |
На экз. ПИН, № 5807/272 (рис. 3г–3е) в глубине альвеолы переднего резца (DI1) видна сравнительно широкая коронка I1 с несформированным корнем (рис. 3е). Зуб полностью находится внутри альвеолы и компактно в нее вмещается, впереди него имеется небольшое пространство для корня DI1. На этой стадии вершина C1 немного возвышается над альвеолярным краем зубной кости (экз. ПИН, №№ 5807/265, 272, 273), его коронка не вполне сформирована, корень отсутствует. Р1 крупный, узкий, с коротким талонидом, без следов стирания (рис. 3г, 3д). Его корни полностью сформированные, длинные, передний существенно тоньше заднего. Вершина Р2 не выступает из альвеолы, над его коронкой сохранилась перемычка верхней части межкорневой перегородки альвеолы DP2 (рис. 3д). Вершина Р4 едва возвышается над альвеолярным краем зубной кости, альвеолы корней DP4 широко расставлены.
На экз. ПИН, № 5807/267 (рис. 3ж, 3з) I1 слегка выступает из альвеолы, имеет широкую коронку шпателевидной формы и, судя по рентгенографии, сравнительно короткий корень. Межкорневая стенка альвеолы P2 резорбирована. Р4 в стадии прорезывания почти на половину высоты коронки выступает над альвеолярным краем зубной кости.
У экз. ПИН, № 5807/255 двухлопастная коронка I2 лежит в основании альвеол, причем ее передняя лопасть располагается под альвеолами DI1 и DI2, а задняя лопасть видна в сломе стенки альвеолы DI3. Двухлопастная коронка I3 в глубине альвеолы DI3 сохранилась на экз. ПИН, № 5807/266.
На экз. ПИН, № 5807/277 P2 в стадии прорезывания развернут в альвеоле передней стороной вверх; вершина Р4 едва выступает над альвеолярным краем зубной кости.
Суммируя приведенные данные, можно отобразить набор одновременно функционирующих и находящихся в процессе смены зубов у Nesophontes рядом последовательных состояний. Для краткости при их характеристике ниже использованы оригинальные формулы зубной смены (DRF, dental replacement formula), в которых буквой “e” (erupting) перед символом зуба (I, C, P) обозначены зубы в состоянии прорезывания, частично показавшиеся над альвеолой (после выпадения молочного предшественника), а буквой “a” (alveolar) обозначены зубы, полностью или почти полностью скрытые в альвеолах, при фактическом или реконструированном наличии молочного предшественника; отсутствующие из-за состояния сохранности зубы в формулах пропущены (знак “-”); полностью прорезавшиеся функциональные зубы не имеют специального знака перед символами; моляры опущены, поскольку не имеют смены.
Для верхних зубов N. major известны два состояния: ---eC1P1eP2eP4 (экз. ПИН, № 5807/269) и I1I2I3C1P1P2P4 (взрослые экземпляры). Для нижних зубных рядов фактически установлены следующие сочетания сменяющихся зубов: --aI3aC1-DP2aP2aP4 (экз. ПИН, № 5807/271), ---aC1-aP2aP4 (экз. ПИН, № 5807/261), ----P1aP2aP4 (экз. ПИН, № 5807/262), -----aP2aP4 (экз. ПИН, № 5807/78), ------DP4aP4 (экз. ПИН, № 5807/71), ----P1-eP4 (экз. ПИН, № 5807/263), -----aP2eP4 (экз. ПИН, № 5807/276). Объединение этих данных позволяет реконструировать следую-щую последовательность состояний DRF: --aI3aC1P1DP2aP2DP4aP4 → ---eC1P1aP2eP4 → I1I2I3C1P1P2P4 (у взрослых экземпляров).
Для нижних зубных рядов N. micrus фактически установлены следующие сочетания зубов: aI1--aC1P1aP2aP4 (экз. ПИН, № 5807/272), -aI2-aC1--- (экз. ПИН, № 5807/255), --aI3--aP2aP4 (экз. ПИН, № 5807/266), -----aP2DP4aP4 (экз. ПИН, № 5807/260), -----aP2aP4 (экз. ПИН, № 5807/277), eI1-----eP4 (экз. ПИН, № 5807/267). Синтез этих данных позволяет реконструировать следующую последовательность состояний DRF: aI1aI2aI3aC1P1DP2aP2DP4aP4 → eI1-----eP4 → I1I2I3C1P1P2P4.
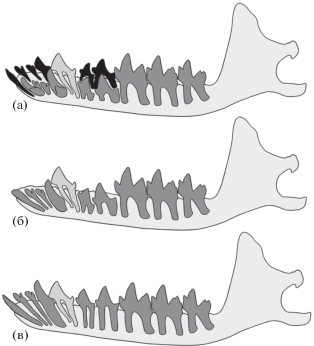
В итоге для изученных видов Nesophontes можно реконструировать следующую последовательность основных стадий прорезывания и смены нижних зубов и состояний DRF (с учетом моляров): DI1aI1DI2aI2DI3aI3DC1aC1P1DP2aP2DP4aP4M1M2M3 (рис. 4а) → eI1eI2eI3eC1P1eP2eP4M1M2M3 (рис. 4б) → I1I2I3C1P1P2P4M1M2M3 (рис. 4в). Предполагаемая последовательность прорезывания клыка и премоляров – P1, C1, P4, P2.
Рис. 4.
Схема основных стадий смены нижних зубов у Nesophontes (полный левый зубной ряд с лабиальной стороны): а – стадия молочных зубов и моляров (DI1aI1DI2aI2DI3aI3DC1aC1P1DP2aP2DP4aP4M1M2M3); б – стадия прорезывания передних зубов (eI1eI2eI3eC1P1eP2eP4M1M2M3); в – взрослая стадия (I1I2I3C1P1P2P4M1M2M3). Черным цветом обозначены молочные зубы, темно-серым – постоянные зубы, светло-серым – передний премоляр с одной установленной сменой.
Судя по изученным материалам, после завершения смены зубов рост животных продолжался. Последующие изменения в строении передней части зубного аппарата незофонтов, включающие формирование заметных межзубных промежутков в ряду премоляров, были связаны с удлинением челюстей.
Итак, у незофонтид в молочной зубной смене представлены резцы, клыки и премоляры. В онтогенезе смена всех этих зубов (кроме P1) происходила после прорезывания моляров, но до достижения животным взрослых размеров. Таким образом, по характеру процесса зубной смены незофонтиды существенно отличаются от большинства представителей Lipotyphla и сближаются с Solenodontidae [см. 10, 13, 14].
Список литературы
MacPhee R.D.E., Flemming C., Lunde D.P. “Last occurrence” of the Antillean insectivoran Nesophontes: new radiometric dates and their interpretation // American Museum Novitates. 1999. № 3261. P. 1–20.
Orihuela J., Vinola L.W., Jiménez Vásquez O., et al. Assessing the role of humans in Greater Antillean land vertebrate extinctions: new insights from Cuba // Quaternary Science Reviews. 2020. V. 249: 106597.
Condis Fernández M.M., Jiménez Vásquez O., Arredondo C. Revisión taxonómica del género Nesophontes (Insectivora: Nesophontidae) en Cuba: análisis de los caracteres diagnóstico // Monografies de la Societat d’Història Natural de les Balears. 2005. № 12. P. 95–100.
Rzebik-Kowalska B., Wołoszyn B.W. New data on Nesophontes subfossil populations from Cuba and Isla de la Juventud (Cuba) // Neues Jahrbuch für Geologie und Paläontologie Abhandlungen. 2012. V. 263. № 2. P. 155–166.
Morgan G.S., MacPhee R.D.E., Woods R., Turvey S.T. Late Quaternary fossil mammals from the Cayman Islands, West Indies // Bulletin of the American Museum of Natural History. 2019. № 428. P. 1–79.
Buckley M., Harvey V.L., Orihuela J., et al. Collagen sequence analysis reveals evolutionary history of extinct West Indies Nesophontes (island-shrews) // Molecular Biology and Evolution. 2020. V. 37. № 10. P. 2931–2943.
Woods R., Turvey S.T., Brace S., et al. Rapid size change associated with intra-island evolutionary radiation in extinct Caribbean “island-shrews” // BMC Evolutionary Biology. 2020. V. 20. № 106. P. 1–12.
Silva Taboada G., Suárez Duque W., Díaz Franco S. Compendio de los mamíferos terrestres autóctonos de Cuba vivientes y extinguidos. La Habana: Museo Nacional de Historia Natural, 2007.
Anthony H.E. Mammals of Porto Rico, living and extinct. Chiroptera and Insectivora. In: Scientific Survey of Porto Rico and the Virgin Islands. New York: New York Acad., 1927. V. 9, Pt. 1. P. 140–149.
McDowell S.B. The Greater Antillean insectivores // Bulletin of the American Museum of Natural History. 1958. № 115. P. 113–214.
Suárez W., Díaz-Franco S. A new fossil bat (Chiroptera: Phyllostomidae) from a Quaternary cave deposit in Cuba // Caribbean Journal of Science. 2003. V. 39. № 3. P. 371–377.
Turvey S.T. Evolution of non-homologous venom delivery systems in West Indian insectivores? // Journal of Vertebrate Paleontology. 2010. V. 30. № 4. P. 1294–1299.
Orihuela J. Endocranial morphology of the extinct Antillean shrew Nesophontes (Lipotyphla: Nesophontidae) from natural and digital endocasts of Cuban taxa // Palaeontologia Electronica. 2014. V. 17. № 2. P. 1–21.
Brace S., Thomas J.A., Dalén L., et al. Evolutionary history of the Nesophontidae, the last unplaced Recent mammal family // Molecular Biology and Evolution. 2016. V. 33. № 12. P. 3095–3103.
Дополнительные материалы отсутствуют.
Инструменты
Доклады Российской академии наук. Науки о жизни