

Доклады Российской академии наук. Науки о жизни, 2021, T. 498, № 1, стр. 241-244
ИЗОФОРМЫ ТТК ВЗАИМОДЕЙСТВУЮТ С ДВУМЯ УЧАСТКАМИ БЕЛКА МЕР-1 DROSOPHILA MELANOGASTER
К. Ю. Халисова 1, И. С. Осадчий 1, академик РАН П. Г. Георгиев 1, О. Г. Максименко 1, *
1 Федеральное государственное бюджетное учреждение науки Институт биологии гена Российской академии наук (ИБГ РАН)
Москва, Россия
* E-mail: maksog@mail.ru
Поступила в редакцию 13.01.2021
После доработки 06.03.2021
Принята к публикации 06.03.2021
Аннотация
Белок ТТК дрозофилы участвует в процессах клеточной дифференцировки и представлен двумя изоформами, ТТК69 и ТТК88, которые имеют общий N-концевой ВТВ домен и различные С-концевые последовательности. Ранее было показано, что TTK69 репрессирует активность энхансеров и промоторов за счет рекрутирования на хроматин консервативного среди высших эукариот NURD-комплекса. В составе NURD-комплекса дрозофилы был найден белок Mep-1, который взаимодействует с С-концевым районом TTK69. В настоящем исследовании с помощью дрожжевой двугибридной системы нами были картированы взаимодействующие участки белков TTK и Mep-1. В уникальных С-концевых участках изоформ ТТК были определены районы, способные взаимодействовать одновременно с двумя участками белка Мер-1. Полученные результаты демонстрируют, что несмотря на низкую гомологию С-концевых районов, изоформы TTK сохраняют способность взаимодействовать с двумя консервативными участками белка Мер-1, что предполагает функциональную значимость такого взаимодействия.
Ген tramtrack (TTK) был исходно идентифицирован как негативный регулятор раннего эмбриогенеза [1, 2]. Ген кодирует две изоформы, TTK69 и TTK88, которые отличаются по С-концевому домену, включающему два цинковых пальца С2Н2-типа [3]. Изоформа TTK69 участвует в регуляции транскрипции в процессе эмбриогенеза, определяет закладку и развитие нервной системы, спецификацию мышечных клеток, развитие трахеи [4, 5]. Изоформа TTK88 определяет закладку фоторецепторов и сенсорных органов на более поздних этапах развития дрозофилы [5, 6]. Изоформы ТТК имеют N-концевой ВТВ-домен, который формирует гомо-мультимерные комплексы и также может взаимодействовать с ВТВ-доменами других регуляторных факторов дрозофилы [7].
Ранее было показано, что изоформа ТТК69 может рекрутировать на регуляторные элементы консервативный среди высших эукариот NURD-комплекс за счет взаимодействия с белком Mep-1 [8]. NURD-комплекс локализуется в области энхансеров и промоторов и выполняет до конца неисследованные функции в негативной регуляции транскрипции [9]. Предполагается, что TTK репрессирует транскрипцию, участвуя в рекрутировании на регуляторные элементы NURD-комплекс. Целью настоящего исследования стало исследование взаимодействия изоформ TTK69 и ТТК88 с одним из компонентов NURD-комплекса, Mep-1.
Ранее было показано [10], что специфичный для изоформы ТТК69 район (392–500 а.о.) взаимодействует с Мер-1. Для уточнения участка взаимодействия между белками была использована дрожжевая двугибридная система, успешно примененная для исследования взаимодействий между ТТК69 и Мер-1 в предыдущей работе [10].
В первую очередь было исследовано взаимодействие между полноразмерными белками, так как ранее было описано взаимодействие только для изоформы TTK69, а TTK88 оставалась практически неизученной. В результате нами было не только подтверждено взаимодействие между ТТК69 и Мер-1, но и показано взаимодействие с Мер-1 изоформы ТТК88 (рис. 1).
Рис. 1.
Определение в составе белка Mep-1 участка взаимодействия с изоформами TTK в дрожжевой двугибридной системе. Фрагменты Mep-1 были соединены с активационным доменом GAL4 (AD), белки TTK69 и TTK88 – c ДНК-связывающим доменом GAL4 (BD). На схеме показано расположение в молекуле белка Mep-1 цинковых пальцев С2Н2-типа прямоугольниками, линии отражают различные фрагменты белка, аминокислотные остатки соответствующих фрагментов указаны слева. Результаты представлены в колонках справа, где + и – означают наличие или отсутствие взаимодействия соответственно. В качестве отрицательного контроля использовали ДНК-связывающий (BD) домен белка GAL4.
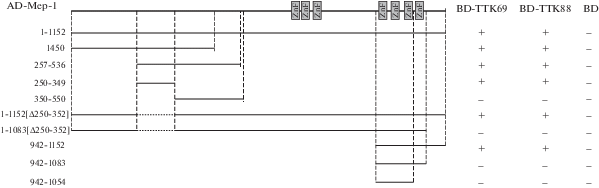
Для картирования района в составе белка Мер-1, с которым взаимодействуют изоформы ТТК, на основе вектора для дрожжевой двугибридной системы были получены различные делеционные производные гена mep-1, кодирующие отдельные части белка. Белок Мер-1 из известных доменов содержит два кластера цинковых пальцев С2Н2-типа – между 681 и 766 а.о. (3 С2Н2-домена) и между 949 и 1083 а.о. (4 С2Н2-домена). В результате проведенного анализа было показано, что обе изоформы ТТК взаимодействуют с ранее описанной частью белка Мер-1, расположенной между 257 и 536 а.о. (рис. 1). Более точное картирование в дрожжевой двугибридной системе позволило локализовать участок в 100 а.о. (250–349) Мер-1, ответственный за данное взаимодействие. Этот участок Мер-1 высоко консервативен среди видов Drosophila, но имеет сравнительно небольшую гомологию при сравнении с другими представителями двукрылых.
Неожиданно оказалось, что делеция 250–349 а.о. в полном белке Мер-1 не нарушает взаимодействие с изоформами ТТК (рис. 1). Этот результат дал основание предполагать, что в белке Мер-1 присутствует второй домен, участвующий в установлении данного взаимодействия. При одновременной делеции 250–349 а.о. и С-концевой части белка Мер-1 взаимодействие с изоформами ТТК исчезает. Этот результат свидетельствует о том, что второй взаимодействующий участок расположен в С-концевой части белка Мер-1, который содержит четыре цинковых пальца С2Н2-типа и неструктурированный район длиной 69 а.о. между 1083 и 1152 а.о. (рис. 1). Для более точного картирования участка, участвующего во взаимодействии, были созданы производные, экспрессирующие разные делеционные производные в С-концевом домене (942–1152 а.о.) белка Мер-1. Результаты, полученные в дрожжевой двугибридной системе, показали, что изоформы ТТК эффективно взаимодействуют с локализованным С-концевым районом (69 а.о.) белка Мер-1. Стоит отметить, что этот район белка Мер-1 является консервативным среди всех насекомых.
Следующей задачей было картирование взаимодействующих с Мер-1 участков в составе изоформ ТТК (рис. 2).
Рис. 2.
Определение в составе белков TTK69 (а) и TTK88 (б) участков взаимодействия с белком Mep-1 в дрожжевой двугибридной системе. На схеме прямоугольниками показано расположение в молекулах белков TTK69 и TTK88 ВТВ-домена и цинковых пальцев С2Н2-типа. Остальные обозначения аналогичны Рис. 1. (в) Сравнение участков белков TTK69 и TTK88, прилегающих к цинковым пальцам и необходимых для взаимодействия с Mep-1. Рамками и линиями показаны участки, предположительно имеющие общее происхождение.
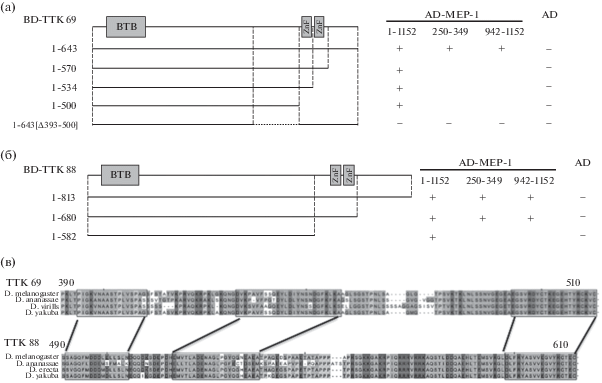
Согласно [10] во взаимодействии с Мер-1 участвует район 392–500 а.о. изоформы ТТК69. Нами был подтвержден этот результат (рис. 2а). Параллельно мы определили, что в составе изоформы ТТК88 с Мер-1 взаимодействует район между 500 и 642 а.о. (рис. 2б) Неожиданно оказалось, что один и тот же район у изоформ ТТК обеспечивает взаимодействие с обеими участками Мер-1. Интересно, что взаимодействующие с Мер-1 участки в составе TTK69 и TTK88 практически не содержат протяженных областей гомологии и не имеют выраженных структурных особенностей (рис. 2в). Белки с BTB-доменами представлены у большинства животных, при этом каждый ген может кодировать много изоформ, у которых BTB-домен – общий, а вариабельная часть возникает путем множественных дупликаций одного и того же экзона, которые становятся альтернативными частями разных изоформ. В процессе эволюции данные экзоны сильно расходятся, в результате вариабельные части изоформ начинают выполнять разные функции. У изоформ TTK такие участки тоже сильно разошлись и при сравнении можно выявить только единичные совпадающие аминокислоты в составе дуплицированного экзона. Поэтому тот факт, что несмотря на отсутствие выраженной гомологии (рис. 2в), участки двух изоформ TTK сохранили взаимодействие с Mep-1 говорит о важности данного взаимодействия в процессе функционирования обеих изоформ TTK.
Наличие двух взаимодействующих с изоформами TTK участков в составе Мер-1 увеличивает специфичность и эффективность рекрутирования NURD-комплекса на хроматиновые мишени. Различные функции изоформ ТТК определяются их уникальными С-концевыми районами, которые отвечают за специфичность связывания каждой из изоформ с определенными регуляторными элементами. При этом С-концевые домены при отсутствии выраженной гомологии сохранили способность взаимодействовать с одними и теми же участками Мер-1. Таким образом, полученные результаты демонстрируют, что взаимодействие ТТК с Мер-1 является важным в процессе рекрутирования NURD-комплекса и репрессии транскрипции целевых генов.
Список литературы
Harrison S.D., Travers A.A. // EMBO J. 1990. V. 9. P. 207–216.
Brown J.L., Wu C. // Development. 1993. V. 117. P. 45–58.
Read D., Manley J.L. // EMBO J. 1992. V. 11. P. 1035–1044.
Ciglar L., Girardot C., Wilczyński B., et al. // Development. 2014. V. 141. P. 2633–2643.
Badenhorst P., Finch J. T., Travers A.A. // Mech. Dev. 2002. V. 117. P. 87–101.
Lai Z.C., Harrison S.D., Karim F., et al. // Proc. Natl. Acad. Sci. U.S.A. 1996. V. 93. P. 5025–5030.
Bonchuk A., Denisov S., Georgiev P., et al. // J Mol Biol. 2011. V. 412. P. 423–436.
Kunert N., Wagner E., Murawska M., et al. // EMBO J. 2009. V. 28. P. 533–544.
Bracken A.P., Brien G.L., Verrijzer C.P. // Genes Dev. 2019. V. 33. P. 936–959.
Reddy B.A., Bajpe P.K., Bassett A., et al. // Mol Cell Biol. 2010. V. 30. P. 5234–5244.
Дополнительные материалы отсутствуют.
Инструменты
Доклады Российской академии наук. Науки о жизни