

Доклады Российской академии наук. Науки о жизни, 2021, T. 498, № 1, стр. 245-248
НОВОЕ МАССОВОЕ ЗАХОРОНЕНИЕ ПЕЩЕРНЫХ МЕДВЕДЕЙ (CARNIVORA, URSIDAE, URSUS KANIVETZ VERESHCHAGIN, 1973) НА СРЕДНЕМ УРАЛЕ
П. А. Косинцев 1, *, Д. О. Гимранов 1, И. А. Лавров 2, А. В. Кисагулов 1
1 Институт экологии растений и животных УрО РАН
Екатеринбург, Россия
2 Ассоциация спелеологов Урала
Кунгур, Россия
* E-mail: kpa@ipae.uran.ru
Поступила в редакцию 02.02.2021
После доработки 09.02.2021
Принята к публикации 10.02.2021
Аннотация
Изучены костные остатки пещерного медведя из нового местонахождения в пещере Прокошева на Среднем Урале (58°13´ с.ш., 58°12´ в.д.). Есть кости всех отделов скелета, все кости целые и на них нет следов деятельности человека или животных. Все они принадлежат большому пещерному медведю (Ursus kanivetz Vereshchagin, 1973). По образцу нижней челюсти взрослой особи получена методом AMS радиоуглеродная дата 53375 ± 765 BP, IGANAMS–8632. Кости принадлежат минимум 18 особям, среди которых 4 особи в возрасте около 1 года, по 1 особи в возрасте около 2 лет и около 3 лет и 12 особей старше 4 лет. Самцам принадлежало 3 черепа и самкам – 7 черепов. Анализ показал, что местонахождение относится к тафономическому типу “естественное кладбище” животных. Это первое на Урале “естественное кладбище” большого пещерного медведя, найденное in situ, не тронутое человеком.
Массовые захоронения без участия человека (тафономический тип “естественные кладбища”) крупных хищных млекопитающих в позднем плейстоцене редки. Известны такие захоронения пещерных львов (Panthera leo spelaea Goldfuss, 1810) [1–3], волков [3], малого пещерного медведя (U. savini Andrews, 1922) [4, 5] и бурого медведя (U. arctos L., 1758) [6–9]. Самыми распространенными местонахождениями подобного типа являются захоронения в пещерах большого пещерного медведя (Ursus spelaeus sensu lato), которые образовались в результате гибели животных во время зимней спячки [10]. Изучение местонахождений этого типа позволяет получить уникальную информацию об экологии и биологии видов.
На Урале известно несколько пещер с большим количеством костей пещерных медведей [11]. Целостность тафоценозов во всех этих пещерах была нарушена деятельностью человека до начала их научного изучения. Пещера Прокошева, о которой пойдет речь в данной публикации, является новым “кладбищем” пещерных медведей. Уникальность этого местонахождения в том, что тафоценоз в ней находится in situ, так как до ее открытия часть пещеры с захоронением пещерных медведей не посещалась человеком. Поэтому мы имеем возможность изучить не нарушенную структуру тафоценоза “естественное кладбище” пещерного медведя. Пещера названа в честь археолога Н.А. Прокошева, исследовавшего пещеры на р. Чусовая в 1930-е годы и погибшего на Великой Отечественной войне.
Пещера Прокошева расположена на Сред- нем Урале, в бассейне р. Чусовая (58°13´ с.ш., 58°12´ в.д.). Она имеет карстовое происхождение и смешанный тип строения [12]: коридор длиной 100 м, который заканчивается обрывом глубиной 8 м и далее переходит в большой грот с ответвлениями. Пол грота покрыт известняковыми глыбами и валунами с заполнением суглинком коричневого цвета, который имеет вязкую консистенцию. “Кладбище” находится в большом гроте. Кости медведей лежат на поверхности пола и частично или полностью погружены в суглинок. По поверхности пола кости распределены относительно равномерно. Анатомически связанные кости отсутствуют. На полу грота следов пребывания людей нет.
Кости собраны с поверхности пола. Из глины они не извлекались. Собрано 64 кости пещерного медведя (коллекция музея ИЭРиЖ УрО РАН, № 2726): 13 черепов, 3 нижних челюсти, 9 изолированных зубов, 2 позвонка, 5 ребер, 1 лопатка, 4 тазовых кости, 3 плечевых, 2 лучевых, 6 локтевых, 3 бедренных, 5 больших берцовых, 3 тарзальных кости, 3 метаподии и 2 фаланги. Все кости целые, но у части черепов и костей посткраниального скелета имеются небольшие разрушения в результате действия конденсированной влаги. На костях нет следов погрызов животными и от орудий человека.
Возраст фауны
Все костные остатки медведей имеют одинаковую окраску и степень фоссилизации. По образцу нижней челюсти взрослой особи получена методом AMS радиоуглеродная дата 53375 ± 765 BP IGANAMS–8632. Это соответствует началу морской изотопной стадии 3 или началу брянского (Moershoofd) интерстадиала [13]. Степень фоссилизации не противоречит этой дате.
Таксономическая принадлежность
Черепа имеют крутой изгиб в области лба, что характерно для пещерных медведей и не характерно для бурых медведей [14]. Зубы (Р4, М1 и М2) без дополнительных бугорков, свойственных малому пещерному медведю. В то же время на Р4 присутствует бугорок на внутренней стороне метаконида, образуя поперечный гребень, что также отличает черепа из пещеры Прокошева от черепов малого пещерного медведя [14]. Морфометрические данные показывают, что изучаемые черепа по размерам больше черепов малого пещерного медведя и медведя Денингера и соответствуют большому пещерному медведю (рис. 1). Таким образом, по морфологическим и морфометрическим признакам черепа из пещеры Прокошева принадлежат большому пещерному медведю (U. kanivetz Vereshchagin, 1973).
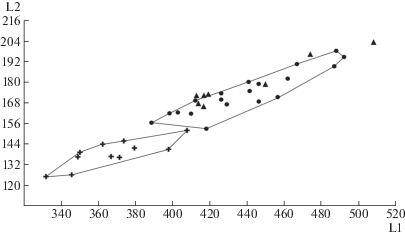
Возрастной состав
Определение возрастной структуры погибших особей проводили по комплексу признаков: стадии прорезывания зубов, прирастанию эпифизов и размерам трубчатых костей [11, 15, 16]. Взрослым особям (старше 4 лет) принадлежит 36 костей: 11 черепов, 2 нижние челюсти, 9 изолированных зубов, позвонок, 2 ребра, лопатка, 2 тазовых кости, 2 плечевых кости, 3 локтевых кости, лучевая, большая берцовая и пяточная кости минимально от 12 особей. Особям в возрасте около 3 лет принадлежит 1 метаподия. Особям в возрасте около 2 лет принадлежат: нижняя челюсть и бедренная кость, минимально от 1 особи. Особям в возрасте около 1 года принадлежат: 2 черепа, 2 тазовые кости, плечевая, лучевая, 2 локтевых, 2 бедренных и 4 больших берцовых кости минимально от 4 особей. Кости принадлежат минимально 18 особям, из которых 12 взрослые. В составе выборки представлены все возрастные группы за исключением группы “новорожденные”. Промежуток между гибелью животных около 1 года, т.е. они гибли в пещере 1 раз в год, во время зимней спячки.
Половой состав
Самцы и самки пещерного медведя хорошо различаются по наибольшим размерам верхних и нижних клыков – клыки у самцов значительно крупнее [17]. По размерам альвеол верхних клыков (C1) был построен график соотношения наибольшей длины (L) и наибольшей ширины (W) альвеол (рис. 2). Выборка черепов разделилась на группы. Группу с бо́льшими размерами альвеол составляют самцы, а группу с меньшими размерами – самки. Самцам принадлежало 3 черепа и самкам – 7 черепов. Полученные результаты показывают, что в пещере погибали и самцы, и самки. Самок погибло значительно больше, чем самцов. Преобладание самок над самцами среди погибших особей также было уставлено для пещер Игнатьевская, Тайн и Геологов 3 на Урале [11].
Рис. 2.
Отношение длины альвеолы верхнего клыка (L) к ширине альвеолы верхнего клыка (W) пещерных медведей из пещеры Прокошева (мм).
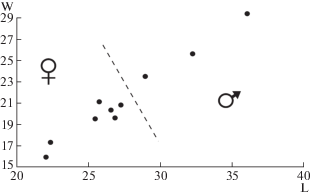
Изученная выборка имеет небольшой объем и сильно смещенный состав. В ней преобладают кости краниального скелета. Это отражает специфику состава костных остатков на поверхности пола. После гибели животных их кости погружались в рыхлые отложения, имеющие вязкую консистенцию. На поверхности оставались наиболее объемные кости (черепа) и кости, в силу случайных причин не погребенные в отложения. Очевидно, что практически все кости погибших особей находятся в рыхлых отложениях пещеры.
В выборке представлены остатки самцов и самок и всех возрастных групп, кроме новорожденных. Гибель животных в пещере происходила 1 раз в год. Есть кости всех отделов скелета, все кости целые и на них нет следов деятельности человека или животных. Все это показывает, что местонахождение относится к тафономическому типу “естественное кладбище” животных, погибших во время зимней спячки. В изученной выборке представлены остатки минимум 18 особей, что позволяет предполагать гибель в пещере нескольких десятков особей.
Список литературы
Merriam J.C., Stock C. The Felidae of Rancho La Brea // Carnegie Institution of Washington Publication. 1932. V. 422. P. 1–232.
Гимранов Д.О., Котов В.Г., Румянцев М.М., и др. Крупнейшее в Северной Евразии захоронение пещерных львов // Доклады академии наук. 2018. Т. 482. С. 234–237.
Верещагин Н.К. Хищные (Carnivora) из бинагадинского асфальта // Труды Естественно-исторического музея им. Г. Зардаби. 1951. Вып. IV. С. 28–140.
Верещагин Н.К. Кизеловская пещера – ловушка зверей на Среднем Урале // Труды Зоологического. ин-та АН СССР. 1982. Т. 111. С. 37–44.
Gimranov D.O., Kosintsev P.A. Quaternary large mammals from the Imanay Cave // Quaternary International. 2020. V. 546. P. 125–134.
Косинцев П.А., Бачура О.П. Массовое захоронение позднеплейстоценовых бурых медведей (Ursus arctos L., 1758) на Северном Урале // Доклады академии наук. 2015. Т. 462. С. 249–251.
Черский И.Д. Об ископаемой фауне Нижнеудинской пещеры // Труды Санкт-Петербургского общества естествоиспытателей. Серия 4: зоологическая. 1888. Вып. 19. С. 66–70.
Crégut E., Fosse P., Argant A., et al. Holocene brown bears (Ursus arctos L.) in natural traps: exceptional sites of Mont Ventoux (Vaucluse, France) // Cadernos Laboratorium Xeolóxico de Laxe Coruña. 2001. V. 26. P. 325–340.
Васильев С.К., Гребнев И.Е. Морфология костей скелета голоценового бурого медведя (Ursus arctos L., 1758) Кузнецкого Алатау // Енисейская провинция. Альманах. 2009. Вып. 4. С. 68–76.
Kurten B. Leaf and death of the Pleistocene cave bear, a study in paleoecology // Acta Zoologica Fennica. 1958. V. 90. P. 1–59.
Косинцев П.А., Воробьев А.А. Биология большого пещерного медведя (Ursus spelaeus Ros. et Hein.) на Урале. Ю.А. Розанов (ред.). В кн.: Мамонт и его окружение: 200 лет изучения. М.: Геос, 2001. С. 266–278.
Оводов Н.Д. Палеофаунистическое изучение пещер. Общие методы изучения истории современных экосистем М.: Наука, 1979. С. 102–128.
Rasmussen S.O., Bigler M., Blockley S.P., et al. A stratigraphic framework for abrupt climatic changes during the Last Glacial period based on three synchronized Greenland ice-core records: refining and extending the INTIMATE event stratigraphy // Quaternary Science Reviews. 2014. V. 106. P. 14–28.
Барышников Г.Ф. Медвежьи (Carnivora, Ursidae). Фауна России и сопредельных стран. Млекопитающие. Т. I. Вып. 5. СПб.: Наука, 2007. 541 с.
Клевезаль Г.А. Принципы и методы определения возраста млекопитающих. М.: Наука, 2007. 283 с.
Fosse P., Crégut-Bonnoure E. Ontogeny/growth of (sub)modern brown bear (Ursus arctos) skeleton: A guideline to appraise seasonality for cave bear (Ursus spelaeus) sites? // Quaternary International. 2014. V. 339–340. P. 275–288.
Kurten B. Sex dimorphism and size trends in the cave bear, Ursus spelaeus Rosenmiiller and Heinroth // Acta Zoologica Fennica. 1955. V. 90. P. 1–48.
Дополнительные материалы отсутствуют.
Инструменты
Доклады Российской академии наук. Науки о жизни