

Доклады Российской академии наук. Науки о жизни, 2021, T. 499, № 1, стр. 344-347
МУТАЦИИ САЙТОВ ФОСФОРИЛИРОВАНИЯ В БЕЛКЕ MSL1 НЕ ВЛИЯЮТ НА ДОЗОВУЮ КОМПЕНСАЦИЮ У Drosophila melanogaster
В. А. Бабоша 1, академик РАН П. Г. Георгиев 1, О. Г. Максименко 1, *
1 Федеральное государственное бюджетное учреждение науки Институт биологии гена Российской академии наук (ИБГ РАН)
Москва, Россия
* E-mail: maksog@mail.ru
Поступила в редакцию 17.03.2021
После доработки 03.04.2021
Принята к публикации 05.04.2021
Аннотация
Белки MSL1 и MSL2 формируют основу комплекса дозовой компенсации дрозофилы, который специфично связывается с Х-хромосомой самцов. Ранее было показано, что фосфорилирование определенных аминокислотных остатков регулирует активность MSL1. В настоящем исследовании были получены трансгенные линии дрозофил, экспрессирующие мутантные варианты белка MSL1, в которых были заменены аминокислоты, подвергающиеся фосфорилированию. В результате показано, что инактивация сайтов фосфорилирования не влияет на эффективность специфичного связывания комплекса дозовой компенсации с Х-хромосомой самцов и его функциональную активность.
Механизм дозовой компенсации у дрозофилы осуществляется посредством увеличения уровня экспрессии генов Х-хромосомы самца (Х/Y) в два раза по сравнению с самками (X/X). За увеличение экспрессии генов Х-хромосомы самцов отвечает мультисубъединичный комплекс дозовой компенсации (КДК), состоящий из пяти белков, MSL1, MSL2, MSL3, MOF и MLE, и двух некодирующих РНК – roX1 (3.7 т.п.н.) и roX2 (0.6 т.п.н.), выполняющих сходные функции [1, 2]. Белки MSL1 и MSL2 создают структурную основу КДК. На N-конце MSL1 находится домен (рис. 1а), формирующий гомодимер, который взаимодействует с двумя молекулами белка MSL2 [3]. С-концевой РЕНЕ-домен белка MSL1 [4] отвечает за взаимодействие с белками MSL3 и MOF (рис. 1а). Белок MOF является основной ацетилтрансферазой, обеспечивающей ацетилирование по лизину в 16 положении гистона H4 [5], которое коррелирует с усилением транскрипции генов.
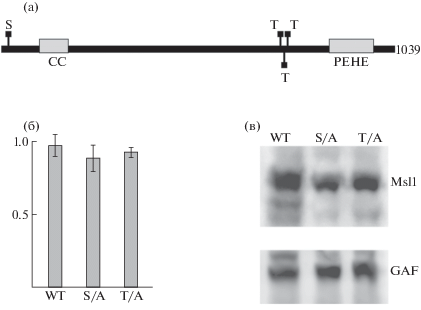
Рис. 1.
(а) Схема белка MSL1. Отмечены остатки серина и треонина, которые были заменены на аланины. (б) Гистограмма, демонстрирующая уровень выживаемости самцов полученных трансгенных линий относительно самок. (в) Результаты иммуноблот-анализа белковых экстрактов, выделенных из полученных трансгенных конструкций. WT – мухи дикого типа. S/A и T/A – мухи, экспрессирующие мутантные варианты белка MSL1 под убиквитиновым промотором. Окраска антителами к белкам Msl1 и GAF.
Белки MSL1, MSL3, MOF и MLE присутствуют также у самок и участвуют в регуляции экспрессии генов по независимому от дозовой компенсации механизму [6]. Белок MSL1 участвует совместно с MOF в формировании альтернативных комплексов, которые связываются с промоторами генов вне зависимости от дозовой компенсации [7]. Было показано, что MSL1 непосредственно функционально взаимодействует с киназой CDK7, которая регулирует активность промоторов генов [8]. При этом фосфорилирование MSL1 регулирует его активность и локализацию на хроматине.
Целью данной работы было исследование роли фосфорилирования MSL1 для правильной локализации комплекса дозовой компенсации на Х-хромосоме самцов. Согласно предыдущему исследованию [8], фосфорилирование белка MSL1 в основном происходит по серину 18 и треонинам в позициях 741, 747 и 751 (рис. 1а). Для исследования функциональной роли фосфорилирования MSL1 нами были получены две мутантные производные, в одной из которых был заменен серин (18) на аланин (MSL1S/A, S18A), а в другой – тре-онины в положениях 741, 747 и 751 на аланины (MSL1T/A, T743A, T747A и T751A).
Для экспрессии мутантных белков в трансгенных линиях дрозофилы кДНК мутантных вариантов MSL1 были встроены в вектор с убиквитиновым (Ubi) промотором, который был ранее успешно использован для экспрессии белка MSL2 [9]. Для получения сравнимых результатов конструкции были интегрированы в геном дрозофилы с помощью φC31-опосредованной рекомбинации по attP-сайту, расположенному в цитогенетическом локусе 86F 3-й хромосомы [10]. Ранее была охарактеризована нуль-мутация в гене msl1, msl1γ185, которая приводит к гибели самцов, но не влияет на выживаемость самок [11]. С помощью генетических скрещиваний с использованием балансеров СyO и TM6, Hu были получены синтетические трансгенные линии, в которых трансгены MSL1S/A(86Fb) или MSL1T/A(86Fb) находились в комбинации с гомозиготной нуль-мутацией msl1γ185. Данные линии были созданы для того, чтобы понять, способен ли мутантный белок, в котором нарушены сайты фосфорилирования, компенсировать нуль-мутацию. Неожиданно оказалось, что экспрессия мутантных белков MSL1 полностью компенсирует отсутствие эндогенного белка MSL1: в обоих синтетических линиях самцы и самки имели сравнимую выживаемость, что предполагает восстановление функции дозовой компенсации (рис. 1б). При этом, согласно результатам иммуноблот-анализа, экспрессия мутантных MSL1 в самцах из синтетических линий находится на сравнимом уровне с самцами из линии дикого типа (рис. 1в). Таким образом, полученные результаты демонстрируют, что инактивация сайтов фосфорилирования в белке MSL1 не влияет на его функциональную активность в дозовой компенсации.
Наиболее хорошо специфичное связывание комплекса дозовой компенсации с Х-хромосомой самцов можно наблюдать на политенных хромосомах из слюнных желез личинок [12]. Нами было проведено исследование связывания белков MSL1 и MSL2 с политенными хромосомами линии дикого типа и синтетических линий MSL1S/A(86Fb) и MSL1T/A(86Fb) (рис. 2). В исследовании были использованы антитела к белкам MSL1 и MSL2, полученные в кроликах [9]. Как и ожидалось, только Х-хромосома самцов линии дикого типа покрыта участками связывания белков MSL1 и MSL2 (рис. 2а). В самках дикого типа окрашивание Х-хромосомы антителами к белкам MSL1 и MSL2 не наблюдалось (рис. 2б). Таким образом, полученные результаты демонстрируют правильность профиля узнавания антител к MSL-белкам. В синтетических линиях MSL1S/A(86Fb) и MSL1T/A(86Fb) паттерн распределения участков связывания белков MSL1 и MSL2 на политенных хромосомах не отличается от контрольной линии самцов дикого типа (рис. 2в, г). Таким образом, мутации, инактивирующие сайты фосфорилирования в белке MSL1, не влияют на эффективность специфичного связывания комплекса дозовой компенсации с Х-хромосомой самцов.
Рис. 2.
Исследование связывания КДК с политенными хромосомами в различных линиях дрозофил: (а) самцы (MSL1 и MSL2) и (б) самки (MSL1) дикого типа, линии с экспрессией мутантных вариантов (в) MSL1S/A и (г) MSL1T/A. Окраска антителами к MSL1 и MSL2.

Полученные нами результаты не согласуются с предыдущим исследованием, в котором было показано, что мутация сайтов фосфорилирования MSL1 приводит к нарушению связывания комплекса дозовой компенсации с Х-хромосомой и летальности самцов [8]. Основным отличием в работе [8] являлось использование индуцируемой системы UAS/GAL4 для экспрессии мутантных вариантов MSL1 на фоне нуль-мутации по msl1. При этом для индукции экспрессии UAS-промотора была использована линия, содержащая GAL4-драйвер под контролем сильного тубулинового промотора (tub-Gal4). Вероятно, избыточная суперэкспрессия мутантных вариантов белка MSL1 приводила к снижению эффективности связывания комплекса дозовой компенсации с Х-хромосомой самцов. Механизм этого явления требует дальнейшего исследования.
Список литературы
Kuroda M.I., Hilfiker A., Lucchesi J.C. // Genetics. 2016. V. 204. P. 435–450.
Samata M., Akhtar A. // Annu. Rev. Biochem. 2018. V. 87. P. 323–350.
Hallacli E., Lipp M., Georgiev P., et al. // Mol. Cell. 2012. V. 48. P. 587–600.
Kadlec J., Hallacli E., Lipp M., et al. // Nat. Struct. Mol. Biol. 2011. V. 18. P. 142–149.
Prestel M., Feller C., Straub T., et al. // Mol. Cell. 2010. V. 38. P. 815–826.
Conrad T., Akhtar A. // Nat. Rev. Genet. 2012. V. 13. P. 123–134.
Straub T., Zabel A., Gilfillan G.D., et al. // Genome Res. 2013. V. 23. P. 473–485.
Chlamydas S., Holz H., Samata M., et al. // Nat. Struct. Mol. Biol. 2016. V. 23. P. 580–589.
Tikhonova E., Fedotova A., Bonchuk A., et al. // Development. 2019. V. 146 P. dev 179663
Bischof J., Maeda R.K., Hediger M., et al. // Proc. Natl. Acad. Sci. U.S.A., 2007. V. 104. P. 3312–3317.
Palmer M.J., Mergner V.A., Richman R., et al. // Genetics. 1993. V. 134. P. 545–557.
Demakova O.V., Kotlikova I.V., Gordadze P.R., et al. // Chromosoma. 2003. V. 12. P. 103–115.
Дополнительные материалы отсутствуют.
Инструменты
Доклады Российской академии наук. Науки о жизни