

Доклады Российской академии наук. Науки о жизни, 2021, T. 499, № 1, стр. 348-353
ЭКЗОГЕННЫЙ МЕЛАТОНИН СНИЖАЕТ ТОКСИЧЕСКОЕ ДЕЙСТВИЕ ПОЛИМЕТАЛЛИЧЕСКОГО СТРЕССА НА РАСТЕНИЯ ЯЧМЕНЯ
Е. Д. Данилова 1, *, И. Е. Злобин 2, член-корреспондент РАН В. В. Кузнецов 1, 2, М. В. Ефимова 1
1 Национальный исследовательский Томский государственный университет
Томск, Россия
2 Институт физиологии растений им. К.А. Тимирязева Российской академии наук
Москва, Россия
* E-mail: nusy.l.d@gmail.com
Поступила в редакцию 09.03.2021
После доработки 05.04.2021
Принята к публикации 05.04.2021
Аннотация
Исследовали влияние мелатонина на ростовые параметры, фотохимическую активность фотосистемы II, содержание основных фотосинтетических пигментов и перекисное окисление липидов в листьях растений ячменя в условиях полиметаллического стресса. Впервые установили, что защитное действие фитомелатонина проявляется в снижении токсического действия полиметаллов на аккумуляцию биомассы, рост корневой системы и размер ассимилирующей поверхности. В основе протекторного действия гормона лежит его способность снижать интенсивность окислительного стресса за счет поддержания уровня каротиноидов и повышения активности супероксиддисмутазы, но не за счет регуляции фотохимической активности хлоропластов. Эффективность применения мелатонина не зависит от длительности его воздействия. Полученные данные могут служить основой для оптимизации технологий применения мелатонина, в качестве вещества индуцирующего состояние прайминга растений.
В настоящее время огромные территории загрязнены солями тяжелых металлов [1], что приводит к снижению продуктивности растений и создает риски попадания токсичных элементов в пищевую цепь человека [2]. Повышенное содержание подвижных форм кадмия, никеля, цинка и других тяжелых металлов присуще кислым почвам, на которые приходится более трети всех пахотных угодий [1, 3]. Закисление почв является результатом индустриальной деятельности, а также чрезмерного использования удобрений, пестицидов и сточных вод для орошения возделываемых земель [1, 3].
Принятые в настоящее время технологии снижения кислотности почв и доступности ионов тяжелых металлов для растений могут оказывать негативное влияние на окружающую среду [3]. Применение соединений гормональной природы для повышения продуктивности растений в условиях техногенного стресса является эффективной и безопасной технологией [2, 4]. Особый интерес представляет фитомелатонин, который представляет собой индоламин (N-ацетил-5-метокси-триптамин). Фитомелатонин способен увеличивать устойчивость растений к действию различных стрессоров, прежде всего, за счет повышения антиоксидантного статуса организма [5–7] и индукции прайминга, т.е. способности повышать устойчивость растений к отсроченному действию повреждающего фактора в результате кратковременной обработки растений мелатонином [4, 8]. Кроме того, известно, что мелатонин снижает токсическое действие меди и кадмия на растениях рапса и редиса соответственно [9, 10], повышает устойчивость арбуза к высоким концентрациям ванадия [11] и активирует антиоксидантную систему Medicago truncatula на фоне действия свинца [12]. Однако в естественной среде растения, как правило, сталкиваются с умеренным повышением уровня доступности в почве одновременно целого ряда тяжелых металлов [13, 14], что делает приоритетной задачей изучение реакции растения именно на полиметаллический стресс. Между тем, защитный эффект мелатонина при действии на растения полиметаллического стресса остается в настоящее время открытым.
Цель данной работы заключалась в том, чтобы выяснить, снижает ли мелатонин токсичность совместного действия ряда тяжелых металлов на растения ячменя и каковы возможные защитные механизмы данного явления.
Исследования проводили на растениях ячменя (Hordеum vulgаre) в условиях, максимально приближенных к условиям произрастания растений в агроценозах и на загрязненных тяжелыми металлами природных территориях. Растения выращивали в грунте с добавлением перлита в течение пяти суток при температуре 20 ± 2°С в фитотроне с 16-часовым фотопериодом. Далее в течение 7 сут растения адаптировали к условиям жидкой питательной среды по Blamey (pH 4.5) [13] с низким содержанием фосфора – 5 мкМ [15]. Такая питательная среда лучше соответствовала реальному составу почвенного раствора на кислых почвах по сравнению с традиционно используемыми питательными средами, где из-за высоких значений рН раствора и высоких концентраций фосфат-ионов доступность ионов металлов для растений (а значит, и их токсический эффект) значительно снижена [13]. Далее часть растений переносили на ту же питательную среду по Blamey на 6 сут (контроль, вариант 1) или на среду с мелатонином (10 мкМ, вариант 2); другие растения (вариант 3) подвергали полиметаллическому стрессу: (Al3+ (20 мкМ); Mn2+ (50 мкМ); Cd2+ (2,8 мкМ); Cu2+ (2 мкМ); Ni2+ (16 мкМ); Zn2+ (40 мкМ); Pb2+ (0,30 мкМ); следующую группу растений (вариант 4) подвергали прикорневой обработке мелатонином (10 мкМ, 1 сут) с последующим полиметаллическим стрессом (6 сут) (прайминг мелатонином); остальные растения (вариант 5) выращивали в течение 6 сут при одновременном действии мелатонина (10 мкМ) и тяжелых металлов. Концентрации используемых тяжелых металлов (Mn2+, Cd2+, Cu2+, Ni2+, Zn2+ и Pb2+) и алюминия (Al3+) подбирали на основании типичных значений концентраций этих ионов в почвенном растворе промышленно загрязненных кислых почв [13, 14]. Через 6 сут после начала эксперимента измеряли ростовые и морфологические характеристики, включая аккумуляцию биомассы, максимальную длину корня, содержание фотосинтетических пигментов и параметры фотохимической активности фотосистемы II (ФС II); растительный материал для определения активности супероксиддисмутазы (СОД) и оценки уровня перекисного окисления липидов (ПОЛ) фиксировали жидким азотом и хранили при – 70°С.
Параметры фотохимической активности ФС II измеряли с помощью РАМ флуориметра (MINI-PAM-II, Heinz-Walz, Germany). Максимальный (Fv/Fm) и эффективный (Y(II)) квантовые выходы ФС II, квантовые выходы нерегулируемой (Y(NO)) и регулируемой (Y(NPQ)) диссипации энергии рассчитывали с использованием программного обеспечения MINI-PAM-II. Содержание основных фотосинтетических пигментов измеряли согласно Лихтенталлеру [16], интенсивность ПОЛ оценивали спектрофотометрическим методом, основанным на образовании окрашенного комплекса – продукта малонового диальдегида (МДА) в реакции с тиобарбитуровой кислотой (ТБК) при нагревании [17]. Определение активности СОД проводили по методу Beauchamp и Fridovich [18]. Эксперименты проводили в трех биологических повторностях; использовали не менее 90 растений на каждый вариант опыта. Полученные результаты представлены в таблицах и на рисунке в виде средней арифметической со стандартной ошибкой. Для сравнения независимых выборок, подчиняющихся закону нормального распределения, использовали параметрический критерий Стьюдента. Значения t-критерия находили для 95% уровня значимости (р < 0.05).
Токсическое действие ионов металлов на растениях ячменя проявлялось в ингибировании накопления биомассы, торможении роста корней и уменьшении площади листовой поверхности на 26, 38 и 23% соответственно (табл. 1, рис. 1). Мелатонин, на фоне полиметаллов, значительно снижал токсическое действие стресса на ростовые процессы; наиболее эффективным было кратковременное воздействие мелатонина на растения (табл. 1, рис. 1).
Таблица 1.
Влияние мелатонина на ростовые показатели растений ячменя в условиях полиметаллического стресса (ПМ)
| Суммарная площадь листовой поверхности, см2 | Длина корня, см | Сырая масса всего растения, г | |
|---|---|---|---|
| Контроль | 12.48 ± 0.21 | 11.68 ± 0.28 | 0.96 ± 0.03 |
| Мелатонин (6 сут) | 13.10 ± 0.24 | 13.24 ± 0.19* | 1.07 ± 0.09 |
| % от контрольного значения | 105 | 113 | 111 |
| ПМ (6 сут) | 9.59 ± 0.17* | 7.24 ± 0.31* | 0.71 ± 0.02* |
| % от контрольного значения | 77 | 62 | 74 |
| Мелатонин (1 сут) + ПМ (6 сут) | 11.87 ± 0.14*,# | 11.18 ± 0.32# | 0.87 ± 0.02# |
| % от контрольного значения | 95 | 96 | 91 |
| Мелатонин (6 сут) + ПМ (6 сут) | 12.16 ± 0.13# | 12.74 ± 0.36# | 0.84 ± 0.02# |
| % от контрольного значения | 97 | 97 | 87 |
Рис. 1.
Влияние кратковременной (1 сут) и длительной (6 сут) обработки мелатонином растений ячменя на фоне действия полиметаллов и алюминия.
1 – контрольный вариант, 2 – полиметаллический стресс (6 сут), 3 – 1 сут предобработки растений мелатонином с последующим действием полиметаллов (6 сут), 4 – одновременное влияние мелатонина и полиметаллического стресса (6 сут).

Способность поддерживать высокий уровень фотосинтетических пигментов является важным показателем физиологического состояния растения в условиях стресса. Из полученных данных следует, что избыточное содержание тяжелых металлов и алюминия в питательном растворе вызывало снижение уровня хлорофиллов и каротиноидов (на 18–21%) по сравнению с их содержанием в растениях контрольного варианта (табл. 2). Предобработка растений мелатонином или его внесение в питательный раствор одновременно с полиметаллами сопровождались снижением их токсического действия на содержание пигментов.
Таблица 2.
Влияние мелатонина (10 мкМ) на состояние фотосинтетического аппарата листьев ячменя на фоне полиметаллического стресса (ПМ)
| Содержание фотосинтетических пигментов, мг/г сырой массы | Параметры фотосинтетической активности ФСII | ||||||
|---|---|---|---|---|---|---|---|
| Хф а | Хф b | Кароти-ноиды | Fv/Fm | Y(II) | Y(NO) | Y(NPQ) | |
| Контроль | 0.524 ± ± 0.029 | 0.139 ± ± 0.015 | 0.151 ± ± 0.013 | 0.804 ± ± 0.014 | 0.702 ± ± 0.023 | 0.204 ± ± 0.021 | 0.104 ± ± 0.011 |
| Мелатонин (6 сут) | 0.562 ± ± 0.034 | 0.152 ± ± 0.019 | 0.162 ± ± 0.019 | 0.796 ± ± 0.009 | 0.705 ± ± 0.017 | 0.207 ± ± 0.015 | 0.101 ± ± 0.012 |
| % от контрольного значения | 107 | 109 | 107 | 99 | 100 | 101 | 97 |
| ПМ (6 сут) | 0.414 ± ± 0.027* | 0.113 ± ± 0.014 | 0.124 ± ± 0.008 | 0.773 ± ± 0.013 | 0.681 ± ± 0.012 | 0.231 ± ± 0.013 | 0.092 ± ± 0.012 |
| % от контрольного значения | 79 | 82 | 82 | 96 | 97 | 113 | 88 |
| Мелатонин (1 сут) + ПМ (6 сут) | 0.475 ± ± 0.058 | 0.131 ± ± 0.017 | 0.136 ± ± 0.012 | 0.793 ± ± 0.023 | 0.693 ± ± 0.013 | 0.214 ± ± 0.021 | 0.092 ± ± 0.013 |
| % от контрольного значения | 91 | 94 | 90 | 99 | 99 | 105 | 88 |
| Мелатонин (6 сут) + ПМ (6 сут) | 0.507 ± ± 0.024# | 0.143 ± ± 0.012 | 0.152 ± ± 0.009# | 0.784 ± ± 0.011 | 0.692 ± ± 0.021 | 0.213 ± ± 0.014 | 0.091 ± ± 0.014 |
| % от контрольного значения | 97 | 103 | 101 | 98 | 99 | 104 | 88 |
Использование мелатонина (6 сут, 10 мкМ) на фоне полиметаллического воздействия повышало содержание хлорофиллов и каротиноидов относительно стрессовых условий на 24–27%, тогда как эффективность кратковременного воздействия мелатонина при последующем полиметаллическом загрязнении была ниже и составляла 16–18% (табл. 2). Следует отметить, что именно каротиноиды, являющиеся неферментативными компонентами антиоксидантной системы, участвуют в тушении активных форм кислорода (АФК) и снижении интенсивности окислительного стресса [19]. На этом основании можно предположить, что поддержание уровня каротиноидов в растениях ячменя при полиметаллическом стрессе является одной из регулируемых мелатонином защитных реакций.
Для выявления действия тяжелых металлов, алюминия и экзогенного мелатонина на фотосинтетическую функцию листьев ячменя оценивали параметры фотохимической активности ФС II. Значение максимальной квантовой эффективности (Fv/Fm) отражает потенциальный квантовый выход ФС II и является чувствительным индикатором функционального состояния фотосинтетической системы. Полученные данные свидетельствуют о том, что Fv/Fm листьев контрольных растений составляла 0.804 (табл. 2), что соответствует величинам данного показателя, характерным для растений, не подвергнутым стрессорным воздействиям [7]. Ионы тяжелых металлов и алюминия несколько снижали максимальный квантовый выход флуоресценции ФС II (Fv/Fm) до 0.773, тогда как мелатонин частично восстанавливал значения Fv/Fm (табл. 2).
Из представленных данных видно, что эффективность фотохимических реакций ФС II, характеризующаяся величиной эффективного квантового выхода (Y(II)), и квантовый выход нефотосинтетической регулируемой диссипации энергии (Y(NPQ) в присутствии полиметаллов в среде незначительно понижались относительно контрольных растений, тогда как квантовый выход нерегулируемой диссипации энергии (Y(NO)) несколько возрастал (табл. 2). Различная по продолжительности прикорневая обработка растений мелатонином на фоне полиметаллического воздействия оказывала некоторый защитный эффект на фотохимическую активность ФС II, достоверность которого, тем не менее, не была доказана.
Избыточные концентрации тяжелых металлов активируют генерацию АФК, одним из проявлений которой является перекисное окисление липидов. Критерием оценки ПОЛ является содержание малонового диальдегида в реакции с тиобарбитуровой кислотой. Полученные данные показали, что полиметаллический стресс увеличивал уровень МДА на 52–74% по отношению к контролю (табл. 3). На фоне стресса мелатонин частично снижал интенсивность ПОЛ [20]. Наиболее эффективной была кратковременная предобработка растений мелатонином, при которой уровень ПОЛ в листе и стебле снижался на 14% по сравнению с техногенным стрессом (табл. 3).
Таблица 3.
Влияние кратковременной и длительной обработки мелатонином (10 мкМ) на величину перекисного окисления липидов на фоне полиметаллического стресса (ПМ)
| Содержание МДА, нМ/г сырой массы | |||
|---|---|---|---|
| Лист | Стебель | Корень | |
| Контроль | 39.14 ± 3.81 | 117.47 ± 7.13 | 63.87 ± 4.53 |
| Мелатонин (6 сут) | 35.42 ± 2.49 | 128.12 ± 5.94 | 70.32 ± 6.11 |
| % от контрольного значения | 91 | 109 | 110 |
| ПМ (6 сут) | 62.57 ± 5.38* | 203.86 ± 9.72* | 97.04 ± 5.74* |
| % от контрольного значения | 160 | 174 | 152 |
| Мелатонин (1 сут) + ПМ (6 сут) | 55.01 ± 4.60 | 175.06 ± 10.16* | 94.06 ± 8.73* |
| % от контрольного значения | 141 | 149 | 147 |
| Мелатонин (6 сут) + ПМ (6 сут) | 61.50 ± 4.50* | 176.78 ± 8.12* | 97.33 ± 3.17* |
| % от контрольного значения | 157 | 150 | 152 |
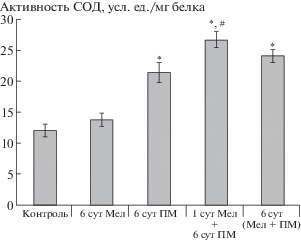
Для снижения негативного влияния окислительного стресса, вызываемого тяжелыми металлами, растения используют систему детоксикации АФК, важным компонентом которой является фермент супероксиддисмутаза. В ответ на присутствие в среде ионов тяжелых металлов и алюминия растения отвечали увеличением активности СОД на 78% относительно контрольных значений (рис. 2). Прайминг растений мелатонином был эффективнее длительного воздействия гормона и способствовал повышению активности СОД по сравнению с действием одного полиметаллического стресса на 22% (рис. 2).
Рис. 2.
Влияние мелатонина на активность СОД в листьях растений ячменя на фоне полиметаллического стресса.
Примечание. *p < 0.05 при сравнении с контрольным значением; #p < 0.05 при сравнении с ПМ.
Таким образом, нами впервые получены данные о протекторной роли мелатонина в растениях ячменя на фоне совместного действия ионов ряда тяжелых металлов и алюминия (Al3+, Mn2+, Cd2+, Cu2+, Ni2+, Zn2+ и Pb2+). Защитный эффект мелатонина проявлялся на уровне аккумуляции биомассы, роста корневой системы, поддержания площади ассимилирующий поверхности и содержания основных фотосинтетических пигментов, но не фотохимической активности ФС II, которая оказалась достаточно устойчивой к полиметаллическому стрессу. Степень наблюдаемого протекторного эффекта мелатонина не зависела от длительности гормональной обработки. В основе положительного влияния мелатонина, очевидно, лежит его способность активировать клеточную антиоксидантную систему, о чем свидетельствуют рост активности СОД при кратковременном прайминге растений мелатонином и поддержание уровня каротиноидов в условиях длительного стресса. Способность мелатонина вызывать состояние прайминга на фоне действия тяжелых металлов может быть использована для разработки эффективных технологий защиты растений в условиях техногенного стресса.
Список литературы
Srivastava V., Sarkar A., Singh S., et al. // Frontiers in Environmental Science. 2017. V. 5. № 64.
Bucker-Neto L., Paiva A.L., Machado R.D., et al. // Genet. Mol. Biol. 2017. V. 40 (1). P. 373–386.
Bian M., Zhou M., Sun D., et al. // The Crop Journal. 2013. V. 1. № 2. P. 91–104.
Efimova M.V., Khripach V.A., Boyko E.V., et al. // Doklady Biological Sciences. 2018. V. 478. № 1. P. 33– 36.
Arnao M.B., Marino B., Hernandez-Ruiz J. // Trends in Plant Science. 2019. V. 24. P. 38–48.
Moustafa-Farag M., Elkelish A., Dafea M., et al. // Molecules. 2020. V. 25. P. 5359.
Danilova E.D., Kolomeichuk L.V., Efimova M.V., et al // Doklady Biochemistry and Biophysics. 2020. V. 495. P. 271–275.
Kolomeichuk L.V., Danilova E.D., Khripach V.A., et al. // Russian Journal of Plant Physiology. 2021. V. 68. № 3. P. 297–307.
Kholodova V.P., Vasil’ev S.V., Efimova M.V., et al. // Russian Journal of Plant Physiology. 2018. V. 65. P. 882–889.
Xu L., Zhang F., Tang M., et al. // Journal of Pineal Research. 2020. V. 69. № 1. № 12659.
Nawaz A., Jiao Y., Chen Ch., et al. // Journal of Plant Physiology. 2018. V. 220. P. 115–127.
Zhang X., Zhang H., Zhang H., et al. // Fronties in Microbiology. 2020. V. 11. № 516.
Li Z., Wang P., Menzies N.W., et al. // Ecotoxicology and Environmental Safety. 2018. V. 147. P. 872–880.
Kopittke P.M., Blamey F.P.C., Asher C.J., et al. // Journal of Experimental Botany. 2010. V. 61. P. 945–954.
Blamey F.P.C., Hernandez-Soriano M.C., Cheng M., et al. // Plant Physiology. 2015. V. 169. № 3. P. 2006– 2020.
Lichtenthaler H.K. // Methods Enzymology. 1987. V. 148. P. 350–382.
Buege J.A., Aust S.D. // Methods Enzymology. 1978. V. 52. P. 302–310.
Beauchamp C., Fridovich I. // Analytical Biochemistry. 1971. V. 44. P. 276–287.
Stahl W., Sies H. // Molecular Aspects of Medicine. 2003. V. 24. № 6. P. 345–351.
Boyko E.V., Golovatskaya I.F., Bender O.G., et al. // Russian Journal of Plant Physiology. 2020. V. 67. P. 351– 359.
Дополнительные материалы отсутствуют.
Инструменты
Доклады Российской академии наук. Науки о жизни