

Доклады Российской академии наук. Науки о жизни, 2021, T. 499, № 1, стр. 312-316
АРОМОРФОЗЫ В ЭВОЛЮЦИИ ОДНОКЛЕТОЧНЫХ ЭУКАРИОТ (НА ПРИМЕРЕ ФОРАМИНИФЕР – FORAMINIFERA D’ORBIGNY, 1826)
В. И. Михалевич *
Зоологический институт Российской академии наук
Петербург, Россия
* E-mail: mikha07@mail.ru
Поступила в редакцию 10.07.2020
После доработки 22.04.2021
Принята к публикации 23.04.2021
Аннотация
Ароморфозы одноклеточных организмов почти не исследованы. Фораминиферы дают уникальную возможность такого изучения, поскольку они сложно устроены и наиболее полно представлены в геологической летописи. В их развитии впервые выявлено более 10 ароморфозов, возникших в разных классах фораминифер независимо и параллельно. Из них ключевые: возникновение агглютинированной, а затем секреционной известковой раковины, бифонтинальной (bilamellar) стенки, многокамерности, дифференциации камер, интегрирующей системы каналов и ядерного дуализма. Они представляют своеобразные пути эволюции на одноклеточном уровне. Многокамерность можно сравнить с многоклеточностью, дифференциацию камер – с дифференциацией тканей, систему каналов, поразительную по сложности и разносящую О2 – с кровеносной системой Metazoa.
Закономерности эволюционного развития живых организмов были изучены, главным образом, на многоклеточных. Одноклеточные эукариоты такими исследованиями затронуты меньше. Фораминиферы (Ф) представляют уникальную группу для изучения эволюции одноклеточных, так как имеют необычайно сложный для одноклеточного уровня скелет, широко распространены в современных морях и наиболее полно представлены в геологической летописи.
Первые однокамерные безраковинные Ф11 возникли в неопротерозое ~ 950–1350 млн лет назад [1]. Настоящая статья представляет попытку распространить понятие “ароморфоз” (А) как “прогрессивное эволюционное изменение строения, приводящее к общему повышению уровня организации организмов” [2] на одну из наиболее сложно устроенных групп одноклеточных. В их развитии выявлены следующие А:
1. Приобретение безраковинными Ф сначала органической (со), а в кембрии – твердой агглютинированной (са) раковины (Р) (рис. 1), обеспечивающей защиту и лучшую выживаемость. В процессе эволюции Ф именно скелет, отражающий общий план строения их организма, претерпел наибольшие изменения.

2. Переход от однокамерных Р к многокамерности (М) – наиболее важный, ключевой ароморфоз (А) в развитии Ф. Он представляет особый путь эволюции – вариант полимеризации внутриклеточных компонентов в пределах одной клетки, сравнимый с полимеризацией клеток в организме. М возникла во всех классах Ф, кроме самого примитивного из них – Astrorhizata (рис. 1). У Miliolata и Rotaliata возникли супермногокамерные (СМ) Р, что привело к еще большему повышению сложности строения и появлению жизненной стратегии, сходной с многоклеточными (продолжительность жизни 4–6 лет, повторные циклы размножения у одной особи).
3. Следующий шаг – дифференциация камер сначала по форме, а затем и по функции (рис. 1): флотирующие, выводковые (ВК), зародышевые (ЗК) и слои камер разного строения (Miliolata, Spirillinata, Rotaliata) у глубоко дифференцированных (ГД) Р, что сравнимо с дифференциацией клеток многоклеточных и образованием тканей. Возникновение М (рис. 1, 2) кардинально изменило план строения организма, привело к усложнению его структуры.
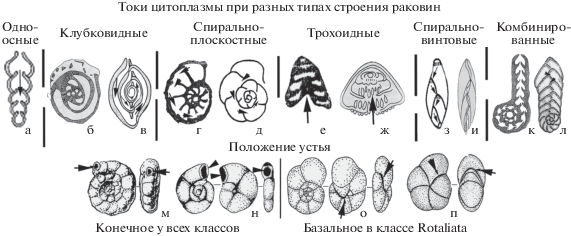

4. Тип строения Р определяет тип токов цитоплазмы (Ц) внутри организма (рис. 2а–л), обеспечивающий сообщение начальных камер с последующими и с внешней средой, способствуя активизации физиологических процессов. Этот путь наиболее длинный и сложный у клубковидных Р (рис. 2б, 2в), наиболее прямой – у одноосных (рис. 2а). Трохоидные Р (рис. 2е, 2ж) классов Spirillinata и Rotaliata более компактны и прочны, а открытая пупочная область создает наиболее короткий и прямой путь циркуляции Ц.
5. Влияет на токи Ц и положение устья (У). Оно конечное (рис. 2а–г, 2м, 2н) на начальных этапах эволюции во всех классах, кроме роталиат, где У имеет базальное положение (рис. 2о, 2п), что привело к глубокому изменению плана строения Р и токов Ц (рис. 2д–2ж). Даже при одинаковом спиральноплоскостном типе Р при базальном положении У этот путь короче (рис. 2г и 2д).
6. По мере возрастания числа камер и сложности их внутреннего строения, путь сообщения Ц отдельных камер через межкамерные форамены (ф) (рис. 3а) и всего организма с внешней средой через У становился недостаточным. Возникновение дополнительных проходов для токов Ц внутри камер (проходы – п (рис 3б), туннели (т) (рис. 3в), столоны (с) (рис. 3г–3е)), часто между очень сложными внутренними элементами эндоскелета (э) (рис. 3д–3е) усложнило и диверсифицировало эти потоки, способствуя лучшей взаимосвязи отдельных частей организма и его связи с внешней средой.
7. В связи с усиленным током Ц в области У возникли опорные внутренние устьевые структуры (УС), имеющие своеобразное, часто довольно сложное строение в разных классах (рис. 4а–4г). Объединение этих внутренних УС от камеры к камере в единую интегрирующую УС (ИУС) привело к значительной дифференциации и компартментализации Ц (рис. 4д–4ж) и явилось следующей ступенью в развитии А у Ф.
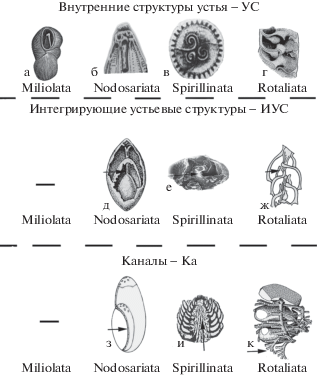
8. Возникновение системы каналов (Ка) (рис. 4з–4к) – ключевой А, в результате которого в организме появляется новая функциональная интегрирующая система. Ка, в отличие от столонов и проходов, ограничены собственными стенками. Они отделяют циркулирующую в них обогащенную 02 эктоплазму от эндоплазмы камер. По функции они сравнимы с кровеносной системой многоклеточных, что поразительно для одноклеточного уровня организации и не встречается ни в одной другой группе протистов. Наиболее сложные системы каналов развиты у роталиат [7], более примитивные – у спириллинат [5, 8, 9].
9. Возникновение параллельно во всех многокамерных классах известковой секреционной монофонтинальной стенки (ссм) со своеобразной ультраструктурой в каждом из них (рис. 1) – один из ключевых А у Ф, так же как возникновение бифонтинальной стенки роталиат (ссб) – известь у них отлагается не по одну сторону над внутренним органическим слоем (вос), а сразу по обе стороны срединной органической мембраны (сом) – (рис. 1). Их стенка обеспечивает наибольшую прочность при наименьшей толщине и весе, обладает наибольшими конструктивными возможностями [5], что создало предпосылки возникновения следующего А (10) – приспособлений к жизни в пелагиали (подкласс Globigerinana) (рис. 1), отразившихся в появлении органелл, обеспечивающих плавучесть.
11. Изменения скелета шли параллельно изменениям живой клетки. Возникновение ядерного диморфизма у милиолят и роталиат (рис. 1) – ключевой А, обеспечивающий дальнейшее развитие. Не случайно именно в этих классах возникли крупные СМ формы с наиболее сложным строением скелета (рис. 1). Дифференциация ядер (Я) на соматические (крупные) и генеративные (мелкие) и, соответственно, ядерных функций сравнима с разделением этих функций в органах многоклеточных. Ядерный аппарат Ф отличает его от всех других типов одноклеточных эукариот (в том числе и от инфузорий [4, 5].
Наибольшее число А произошло в классе Rotaliata, что привело к резкому увеличению числа таксонов, ускорению темпов их эволюции, завоеванию новой экологической ниши – пелагиали [5]. Это самый молодой и высокоорганизованный класс Ф подобно млекопитающим среди позвоночных.
Рассмотренные выше А привели к глубокому изменению плана строения Ф, возрастанию их сложности, поразительной для одноклеточного уровня организации, повышению целостности организма, интенсификации функций и усилению конкурентоспособности, позволили наиболее продвинутым их группам перейти на новый уровень организации и обеспечили пути дальнейшей прогрессивной эволюции. Они происходили в разных классах Ф независимо и параллельно в разное время и имеют своеобразные черты в каждом из них [4]. В отличие от идиоадаптаций, они не связаны напрямую с влиянием окружающей среды. Их можно рассматривать как своеобразный путь эволюции и попытку преодолеть ограничения одноклеточного уровня за счет поразительного увеличения сложности организации. Переход на новый, многоклеточный уровень смог осуществиться при возникновении определенных условий, когда возможности одноклеточного уровня организации оказались исчерпанными.
Список литературы
Groussin M., Pawlowski J., Yang Z. Bayesian relaxed clock estimation of divergent time in Foraminifera // Molecular phylogenetic and evolution. 2011. № 61. P. 157–166.
Северцов А.Н. Морфологические закономерности эволюции. М.–Л: АН СССР, 1939. 610 с.
Михалевич В.И. Макросистема фораминифер // Известия РАН: Серия биол.,1998. № 2. С. 266–271.
Михалевич В.И. Основы зоологии. Protista. Фораминиферы. Петербург: Наука, 2000. С. 525–637.
Mikhalevich V.I. Post-Cambrian Testate Foraminifera as a System in its Evolution. New York: Nova Science Publishers, 2013. 444 p.
Pawlowski J., Holzmann M., Tyszka J. New supraordinal classification of Foraminifera: Molecules meet morphology // Marine micropaleontology. 2013. № 100. P. 1–10.
Hottinger L. Comparative anatomy in selected larger Foraminifera. In: Hedley R.H., Adams C.G. (Eds). Foraminifera. London, 1978. P. 203–206.
Rigaud S., Blau J., Martini R., Rettori R. Taxonomy and phylogeny of Trocholinidae (Involutinina) // Journ. Foraminiferal. Reseach. 2013. V. 43. № 4. P. 317–339.
Schlagintweit F., Rigaud S., Wilmsen M. Cenomanian trocholinids of northern Cantabria // Facies, 2015. № 61. P. 12–27.
Дополнительные материалы отсутствуют.
Инструменты
Доклады Российской академии наук. Науки о жизни