

Доклады Российской академии наук. Науки о жизни, 2021, T. 500, № 1, стр. 402-406
ПЛЕЙСТОЦЕНОВЫЕ ЛИСИЦЫ (Vulpes, Canidae, Carnivora) ИЗ ПЕЩЕРЫ ТАВРИДА, КРЫМ
Д. О. Гимранов 1, 2, *, С. Бартолини-Лученти 3, 4, А. В. Лавров 5, Б. А. Вахрушев 6, академик РАН А. В. Лопатин 5
1 Институт экологии растений и животных
УрО РАН
Екатеринбург, Россия
2 Уральский федеральный университет им. Б.Н. Ельцина
Екатеринбург, Россия
3 Флорентийский университет, Отдел наук о Земле
Флоренция, Италия
4 Отдел геологии и палеонтологии музея естественной истории Флорентийского университета
Флоренция, Италия
5 Палеонтологический институт
им. А.А. Борисяка РАН
Москва, Россия
6 Крымский федеральный университет
им. В.И. Вернадского
Симферополь, Россия
* E-mail: djulfa250@rambler.ru
Поступила в редакцию 20.05.2021
После доработки 19.06.2021
Принята к публикации 19.06.2021
Аннотация
Из раннеплейстоценового местонахождения пещера Таврида (Крым, поздний виллафранк, возраст 1.8–1.5 млн. лет) описаны фрагмент нижней челюсти и четыре изолированных зуба ископаемых лисиц: Vulpes alopecoides (Del Campana, 1913), Vulpes cf. vulpes (Linnaeus, 1758) и Vulpes sp. На основании размерных и морфологических характеристик установлено, что фрагмент нижней челюсти и два M1 принадлежат V. alopecoides. Строение m1 Vulpes cf. vulpes сходно с V. vulpes и V. alopecoides, но размеры намного превосходят известные пределы изменчивости последнего вида. Строение m1 Vulpes sp. является уникальным среди плейстоценовых представителей рода Vulpes, хотя по размерам зуб соответствует m1 V. alopecoides.
В составе раннеплейстоценового комплекса хищных млекопитающих из пещеры Таврида в Крыму, открытой в 2018 г. [1, 2], представлены семейства Felidae, Ursidae, Hyaenidae и Canidae [3–5]. Костные остатки лисиц (род Vulpes, Canidae) были собраны в ходе раскопок в 2018 и 2020 г. Они представлены фрагментом нижней челюсти и четырьмя изолированными зубами (рис. 1–3), которые хранятся в Палеонтологическом институте им. А.А. Борисяка РАН (ПИН), Москва, и Институте экологии растений и животных УрО РАН (ИЭРЖ), Екатеринбург. Описания и измерения зубов проведены по ранее предложенной методике [6]. Измерения (L – длина, W – ширина) в мм, проведены с помощью цифрового штангенциркуля с точностью 0.01 мм. Сравнение размеров зубов проиллюстрировано диаграммами (рис. 4).
Рис. 1.
Vulpes alopecoides (Del Campana, 1913): а–в – экз. ИЭРЖ, № 727/98, левый М1: а – с окклюзиальной стороны, б – с мезиальной стороны, в – с дистальной стороны; г–е – экз. ИЭРЖ, № 727/97, правый М1: г – с окклюзиальной стороны, д – с мезиальной стороны, е – с дистальной стороны; Крым, местонахождение Таврида; нижний плейстоцен. Обозначения: buccin – буккальный цингулюм, Hyc – гипокон, Mec – метакон, mescin – мезиальный цингулюм, Pac – паракон, Parstl – парастиль, postprcr – постпротокриста, Prc – протокон, Tlbas – бассейн талона (гипокона), Trbas – бассейн тригона.
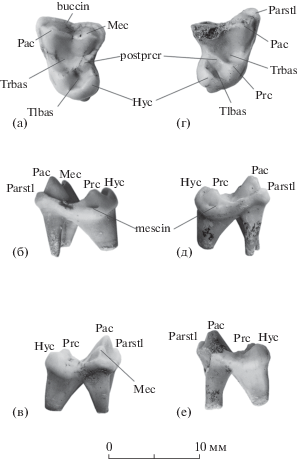
Рис. 2.
Vulpes alopecoides (Del Campana, 1913), экз. ПИН, № 5644/92, фрагмент левой нижнечелюстной кости: а – с буккальной стороны, б – с лингвальной стороны, в – с окклюзиальной стороны; Крым, местонахождение Таврида; нижний плейстоцен. Обозначения: buccind – буккальный цингулид, Encd – энтоконид, Encnd – энтоконулид, Hycd – гипоконид, Mecd – метаконид, Pacd – параконид, PC1 – первый задний дополнительный бугорок, PC2 – второй задний дополнительный бугорок, Prcd – протоконид, Tldbas – бассейн талонида.
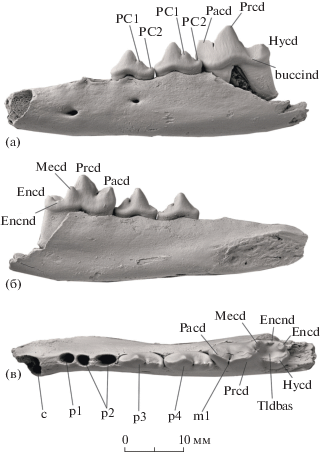
Рис. 3.
Лисицы из пещеры Таврида: а–в – Vulpes cf. vulpes Linnaeus, 1758, экз. ПИН, № 5644/88, левый m1: а – с окклюзиальной стороны, б – с буккальной стороны, в – с лингвальной стороны; г–е – Vulpes sp., экз. ПИН, № 5644/87, правый m1: г – с окклюзиальной стороны, д – с буккальной стороны, е – с лингвальной стороны; Крым, местонахождение Таврида; плейстоцен. Обозначения: posthycr – постгипокристид, Tvcrd – поперечный кристид; др. см. рис. 2.
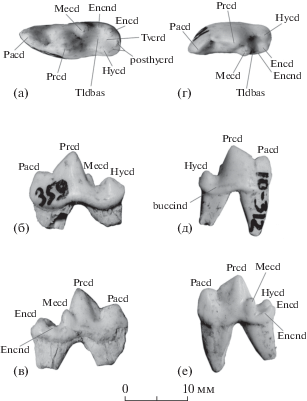
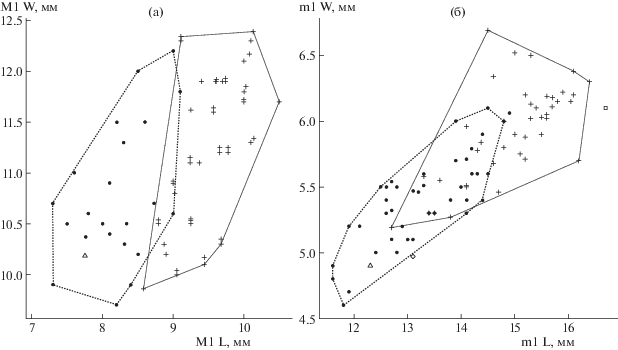
Рис. 4.
Сравнение размеров зубов некоторых представителей рода Vulpes: а – отношение длины (L) к ширине (W) М1 у V. alopecoides (Del Campana, 1913) (круг, данные по [6]), современных V. vulpes Linnaeus, 1758 Евразии (крест) и V. alopecoides (экз. ИЭРЖ, № 727/98) из нижнего плейстоцена пещеры Таврида (треугольник); б – отношение длины (L) к ширине (W) m1 у V. alopecoides (круг, данные по [6]), современных V. vulpes Евразии (крест), V. alopecoides (экз. ПИН, № 5644/92) из плейстоцена пещеры Таврида (треугольник), Vulpes sp. (экз. ПИН, № 5644/87) из плейстоцена пещеры Таврида (ромб) и Vulpes cf. vulpes (экз. ПИН, № 5644/88) из плейстоцена пещеры Таврида (квадрат).
Два изолированных первых верхних моляра на основании размеров и строения отнесены к Vulpes alopecoides (Del Campana, 1913). Строение коронок и корней левого М1 (экз. ИЭРЖ, № 727/98) и правого M1 (экз. ИЭРЖ, № 727/97) идентично (рис. 1), их размеры имеют очень близкие значения (экз. ИЭРЖ, № 727/97: L – 7.61, W – 10.13; экз. ИЭРЖ, № 727/98: L – 7.60, W – 10.18). Зубы найдены недалеко друг от друга в одном слое на одном участке пещеры, они нестертые и имеют одинаковый светло-желтый цвет. На основании этих данных мы предполагаем, что левый и правый М1 принадлежали одной особи. Зубы трехкорневые, вытянутые букколингвально. Паракон несколько больше метакона, имеется небольшой, но хорошо различимый парастиль. Метастиль не развит. Имеется четко выраженный буккальный цингулюм. Протокон хорошо развит, протоконуль и метаконуль отсутствуют, имеется развитая постпротокриста. Гипокон небольшой по площади, высокий, равен по высоте протокону. Имеется прерывистый мезиальный цингулюм, дистальный цингулюм отсутствует. Бассейн тригона больше и глубже, чем бассейн талона.
Найденные М1 (экз. ИЭРЖ, №№ 727/97, 98) имеют морфологические признаки, характерные для V. alopecoides: зубы вытянуты букколингвально, паракон М1 больше метакона, протокон хорошо развит, имеется развитая постпротокриста и крупный гипокон с глубоким бассейном, также имеется мезиальный цингулюм [6]. Размеры М1 входят в пределы изменчивости таковых у V. alopecoides (рис. 4).
Фрагмент левой горизонтальной ветви нижней челюсти (экз. ПИН, № 5644/92) на основании размеров и строения m1 также отнесен к V. alopecoides. У этого экземпляра (рис. 2) утрачены резцовая часть и восходящая ветвь, а тело нижней челюсти обломано на уровне контакта m1 и m2. Переднее подбородочное отверстие расположено на уровне альвеолы переднего корня р2, заднее подбородочное отверстие – на уровне заднего корня р3. Сохранились р3, р4 и m1 и альвеолы клыка, p1 и p2. Первый премоляр p1 однокорневой. Передний корень р2, р3 и р4 немного меньше заднего. Премоляры узкие и имеют по два задних дополнительных бугорка. Второй задний дополнительный бугорок на р3 и р4 цингулярный.
Челюсть принадлежит взрослой особи, так как вершины главных бугорков на зубах (протоконида премоляров и m1, параконида, метаконида, гипоконида и энтоконида m1) заметно стерты (рис. 2). Особенности морфологии m1 соответствуют таковым V. alopecoides. Он двухкорневой, передний корень больше заднего. Разница в высоте параконида и протоконида не очень значительна. Длина протоконида (4.4 мм) ненамного больше длины параконида (3.9 мм). Метаконид большой и четко отделен от протоконида. Талонид занимает 33% длины зуба. Поперечный кристид (трансверсальный гребень) не выражен. Буккальный цингулид выражен на уровне от задней части протоконида до задней части гипоконида. Гипоконид и энтоконид крупные, последний меньше по размеру, бассейн талонида развитый, закрыт лингвально энтоконулидом [6]. Размеры m1 (экз. ПИН, № 5644/92): L – 12.30, W – 4.89 (см. рис. 4).
Левый m1 (экз. ПИН, № 5644/88) по строению коронки не отличается от V. alopecoides и V. vulpes Linnaeus, 1758 (рис. 3a–3в). По длине и ширине (L – 16.69, W – 6.08) он значительно больше средних размеров V. alopecoides и ближе к таковым современных V. vulpes (рис. 4). Зуб двухкорневой, нестертый. Параконид значительно ниже протоконида. Протоконид (6.6 мм в длину) немного длиннее параконида (5.4 мм). Метаконид довольно крупный и четко отделен от протоконида. Талонид занимает 30% длины зуба. Гипоконид крупный. Энтоконид высокий, крупный, немного уступает по размерам гипокониду. Бассейн талонида хорошо развит, закрыт лингвально отчетливым энтоконулидом. Задний гребень гипоконида (постгипокристид) раздваивается в своем основании. Имеется четкий поперечный кристид (рис. 3а).
Морфологические характеристики левого m1 (экз. ПИН, № 5644/88) не выходят за пределы изменчивости V. alopecoides и V. vulpes: метаконид достаточно большой и четко отделен от протоконида, гипоконид крупный, энтоконид высокий, но уступает по размеру гипокониду, бассейн талонида развитый, закрыт лингвально энтоконулидом, имеется поперечный кристид [6]. Размеры зуба сходны с таковыми современной обыкновенной лисицы (V. vulpes). Учитывая ограниченное число экземпляров и возможность смешения разновозрастных плейстоценовых материалов, мы не рассматриваем эту находку (экз. ПИН, № 5644/88) как самое раннее свидетельство появления V. vulpes в геологической летописи, но определяем ее систематическое положение как Vulpes cf. vulpes.
Правый m1 (экз. ПИН, № 5644/87; L – 13.10, W – 4.97) по размерам близок к m1 V. alopecoides (рис. 4), но строение этого зуба не имеет аналогов среди известных представителей Vulpes (рис. 3г–3е). Зуб нестертый, двухкорневой, передний корень больше заднего. Разница в высоте параконида и протоконида не так значительна, как у экз. ПИН, № 5644/88. Длина протоконида (6.1 мм) в 1.5 раза больше длины параконида (4.1 мм). Метаконид большой и четко отделен от протоконида. Тригонид удлинен, талонид редуцирован и занимает 22% длины зуба. Гипоконид очень крупный, несколько смещен центрально. Энтоконид сильно редуцированный, смещен в дистолингвальный угол талонида. Бассейн талонида маленький. Крохотный энтоконулид не закрывает бассейн талонида с лингвальной стороны. Поперечный кристид отсутствует. Буккальный цингулид выражен на уровне от задней части протоконида до задней части гипоконида.
Пропорции основных бугров и морфология правого m1 (экз. ПИН, № 5644/87) резко отличаются от свойственных для V. alopecoides и других древних лисиц [см. 6]. Особенно значимыми признаками являются увеличенная длина протоконида по сравнению с параконидом и короткий талонид. Также важной характеристикой является крупный гипоконид, смещенный несколько центрально. Еще одним существенным признаком является рудиментарный энтоконид. Три перечисленных признака m1 (экз. ПИН, № 5644/87) указывают на гиперкарниворную специализацию этой ископаемой лисицы. Ни один из описанных ранее плейстоценовых видов лисиц не имеет такого комплекса морфологических особенностей m1. Подобная гиперкарнивория нижних хищнических зубов известна для V. qiuzhudingi Wang et al., 2014 и V. rooki Bartolini Lucenti, 2021 из раннего плиоцена Китая [7, 8]. V. qiuzhudingi также имеет очень крупный гипоконид, который, в отличие от экз. ПИН, № 5644/87, занимает значительную площадь талонидного бассейна m1. В этом отношении экземпляр из Тавриды ближе к V. rooki, у которого энтоконид редуцирован не так сильно, как у V. qiuzhudingi. При этом m1 V. qiuzhudingi и V. rooki значительно крупнее; кроме того, у этих двух видов сильнее редуцированы метаконид и энтоконид.
Таким образом, морфологические особенности описанного m1 (экз. ПИН, № 5644/87) не позволяют отнести его к какому-либо известному виду древних лисиц. Объем доступного для описания материала явно недостаточен для выделения нового вида, поэтому данная форма рассматривается здесь как Vulpes sp.
Образцы из пещеры Таврида, отнесенные к представителям рода Vulpes, не очень многочисленны, но их морфологические и морфометрические особенности достаточно интересны. До недавнего времени считалось, что в раннем – среднем плейстоцене Европы существовали три вида лисиц: V. praecorsac Kormos, 1932, V. praeglacialis (Kormos, 1932) и V. alopecoides (Del Campana, 1913). Новейшие исследования [6] показали, что эти виды демонстрируют морфологические характеристики, соответствующие внутривидовой изменчивости современных видов Vulpes; на этом основании был сделан вывод, что они должны быть объединены в один вид V. alopecoides.
Присутствие V. alopecoides в отложениях пещеры Таврида соответствует определению возраста фаунистического комплекса в интервале 1.8–1.5 млн л.н. (поздний виллафранк, MNQ18–MNQ19) [см. 1, 9, 10]. Ранее костные остатки лисицы, сходной с V. alopecoides, были обнаружены на Кавказе в пещере Кударо 1 (Южная Осетия, средний плейстоцен) [11, 12]. Они были отнесены к V. praeglacialis, т.е. к виду, который ныне считается младшим синонимом V. alopecoides [6].
Находка V. alopecoides в нижнеплейстоценовых отложениях пещеры Таврида является первым достоверным свидетельством обитания этого вида в Крыму. Экз. ПИН, № 5644/87 может представлять собой своеобразный и неизвестный в настоящее время вид рода Vulpes из раннего плейстоцена Евразии.
Список литературы
Лопатин А.В., Вислобокова И.А., Лавров А.В., и др. Пещера Таврида – новое местонахождение раннеплейстоценовых позвоночных в Крыму // Доклады Академии наук. 2019. Т. 485. № 3. С. 381–385.
Zelenkov N.V., Lavrov A.V., Startsev D.B., et al. A giant early Pleistocene bird from Eastern Europe: unexpected component of terrestrial faunas at the time of early Homo arrival // Journal of Vertebrate Paleontology. 2019: e1605521.
Лавров А.В., Гимранов Д.О., Старцев Д.Б., Зайцев Б.А. Саблезубые кошки (Carnivora, Felidae, Machairodontinae) из нижнего плейстоцена пещеры Таврида, Крым // Доклады РАН. Науки о жизни. 2020. Т. 495. № 1. С. 567–571.
Гимранов Д.О., Лавров А.В., Старцев Д.Б., и др. Первая находка этрусского медведя (Ursus etruscus, Ursidae, Carnivora) в Крыму (пещера Таврида, ранний плейстоцен) // Доклады РАН. Науки о жизни. 2020. Т. 491. № 1. С. 130–133.
Лавров А.В., Гимранов Д.О., Старцев Д.Б., Лопа-тин А.В. Гигантская гиена Pachycrocuta brevirostris (Hyaenidae, Carnivora) из нижнего плейстоцена пещеры Таврида, Крым // Доклады РАН. Науки о жизни. 2021. Т. 496. № 1. С. 10–14.
Bartolini Lucenti S., Madurell-Malapeira J. Unraveling the fossil record of foxes: an updated review on the Plio-Pleistocene Vulpes spp. from Europe // Quaternary Science Reviews. 2020. V. 236. 106296.
Wang X., Tseng Z., Li Q., et al. From ‘third pole’ to north pole: a Himalayan origin for the arctic fox // Proceedings of the Royal Society B. 2014. V. 281. № 1787: 20140893.
Bartolini Lucenti S. A new large-sized Pliocene fox (Carnivora, Canidae) from Yushe basin (Shanxi, China) // Rivista Italiana di Paleontologia e Stratigrafia. 2021. V. 127. № 1. P. 133–147.
Вислобокова И.А., Титов В.В., Лавров A.В., и др. О находке большерогого оленя рода Arvernoceros в пещере Таврида в Крыму // Доклады Академии наук. 2019. Т. 487. № 5. С. 596–599.
Вислобокова И.А., Титов В.В., Лавров A.В., и др. Раннеплейстоценовые винторогие антилопы (Artiodactyla, Bovidae) из пещеры Таврида (Крым, Россия) // Палеонтологический журнал. 2020. № 1. С. 78–88.
Baryshnikov G. Pleistocene Canidae (Mammalia, Carnivora) from Paleolithic sites in Kudaro caves in the Caucasus // Russian Journal of Theriology. 2012. V. 11. № 2. P. 77–120.
Барышников Г.Ф. Крупные млекопитающие кударской плейстоценовой фауны Кавказа. СПб.: Наука, 2020 (Тр. Зоол. ин-та РАН. Прил. 9).
Дополнительные материалы отсутствуют.
Инструменты
Доклады Российской академии наук. Науки о жизни