

Доклады Российской академии наук. Науки о жизни, 2021, T. 500, № 1, стр. 437-442
ОСОБЕННОСТИ ЦИКЛА СОН–БОДРСТВОВАНИЕ И ЦИРКАДНОЙ АКТИВНОСТИ МАЛОГО ОЛЕНЬКА (Tragulus kanchil)
О. И. Лямин 1, 2, 3, *, Д. М. Сигал 3, Р. В. Евсигнеев 4, Е. А. Назаренко 1, 2, академик РАН В. В. Рожнов 1, 2
1 Институт проблем экологии и эволюции
им. А.Н. Северцова Российской академии наук
Москва, Россия
2 Совместный Российско-Вьетнамский Тропический научно-исследовательский и технологический центр
Ханой, СРВ
3 Калифорнийский университет в г. Лос-Анджелес
Лос-Анджелес, США
4 Самарский Университет
Самара, Россия
* E-mail: oilyamin@yahoo.com
Поступила в редакцию 18.05.2021
После доработки 23.05.2021
Принята к публикации 23.05.2021
Аннотация
Впервые изучены особенности сна и циркадной ритмики у малого оленька (Tragulus kanchil) – самого мелкого (вес 1.5–2.2 кг) представителя базальной группы (Tragulidae) парнокопытных, которые сформировались 40–50 млн. лет назад. В условиях, приближенных к естественным, на 6 животных выполнено 30 сут видеозаписи поведения и 15 сут полисомнографических исследований. Оленьки были активны менее 20% времени суток и 60–80% времени проводили в состоянии покоя. На медленноволновый сон (МС) приходилось 49.7 ± 3.7%, на парадоксальный (РЕМ) сон – 1.7 ± 0.3% времени суток. Большую часть МС (87.0 ± 4.4% времени) глаза у животных были открыты. Наибольшее количество МС и РЕМ сна регистрировалось в дневное время (09–16 ч) и в первой половине ночи (20–02 ч), наибольшая активность – в сумеречные часы (04–06 и 18–19 ч). Мы предполагаем, что основные параметры сна оленьков определяются в первую очередь экологическими факторами, такими как температурные условия и воздействие хищников, а также размерами тела и особенностями физиологии оленьков.
Копытные – одна из наиболее интересных групп для сравнительных исследований особенностей сна животных. Эти млекопитающие обитают в разных биотопах, ведут преимущественно стадный образ жизни, среди них много одомашненных видов. Большинство электрофизиологических исследований сна были выполнены на домашних животных [1, 2] и лишь в трех работах исследовали сон диких парнокопытных [3–5]. Общая продолжительность сна у исследованных видов варьирует в диапазоне от 12 до 43% от времени суток, тогда как количество парадоксального сна (РЕМ сон, сокращение от rapid eye movement) у всех копытных относительно небольшое – от 1.2 до 7.2% от времени суток [1–5].
Оленьки (Tragulidae, Artiodactyla) – самые мелкие среди копытных млекопитающих. Они обитают в тропических лесах Южной Азии и Африки. Оленьки появились 40–50 млн. лет назад и в течение этого времени практически не изменились [6]. Это единственная сохранившаяся из известных базальных групп парнокопытных. У оленьковых, которых называют “живыми ископаемыми”, имеется ряд морфофизиологических и поведенческих признаков, которые считаются “архаичными” для парнокопытных [7]. Учитывая филогенетическое положение, особенности и размер животных, а также их распространение, оленьки представляют особый интерес для сравнительных исследований сна, в частности понимания роли экологических и эволюционных факторов в формировании параметров сна и проверки ряда гипотез о функции сна.
Задача данного исследования состояла в изучении сна и циркадной ритмики у малого оленька (Tragulus kanchil).
МАТЕРИАЛЫ И МЕТОДЫ
Две серии экспериментов были выполнены на 6 взрослых самках оленька (вес 1.7–2.2 кг, длина тела 42–48 см) в национальном парке Бузямап (Вьетнам). В первой серии методом видеосъемки исследовали особенности поведения оленьков (всего 14 дней у 3 животных). Во второй серии провели полисомнографические исследования с применением телеметрии (15 сут регистрации у 4 животных). Всем животным под общей анестезией были вживлены электроды для регистрации электроэнцефалограммы (ЭЭГ) коры головного мозга, электромиограммы шейной мускулатуры и электроокулограммы. Во время экспериментов животные находились по одному в вольере размером 3.5 × 3.5 м с естественной растительностью, в нем находился домик (укрытие) размером 0.9 × × 0.7 × 0.7 м, в котором животные проводили большую часть дневного времени. Режим освещения (рассвет в 06 ч, закат в 18 ч) и шума были естественными. В качестве корма животным два раза в день давали листья кустарников (в 06–07 ч и в 15–16 ч). Кроме того, с деревьев в вольер падали спелые плоды кешью. Электрополиграммы и видеозаписи поведения были обработаны в 20-сек эпохах анализа.
План исследований был одобрен комиссией по биоэтике Института проблем экологии и эволюции им. А.Н. Северцова РАН и Совместного Российско-Вьетнамского Тропического научно-исследовательского и технологического центра.
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
Большую часть дневного времени оленьки находились в укрытии, ночного – в вольере. Две трети времени суток (в среднем 64.0 ± 2.1% в серии 1 и 66.7 ± 4.7% в серии 2) животные были в состоянии покоя в характерной позе, которая напоминала позу сфинкса: оленьки лежали, оперевшись на грудину, конечности подогнуты под туловище, шея вертикально, голова направлена вперед (рис. 1). Примерно 16% времени (18.3 ± ± 3.2% в серии 1 и 14.0 ± 3.4% в серии 2) оленьки стояли. Меньше 2% времени (2.0 ± 0.9% и 1.3 ± ± 0.3%) оленьки лежали, вытянув голову вперед и положив на землю. На периоды высокой активности (передвижение по вольеру и кормление) приходилось 15.7 ± 4.1% времени в серии 1 и 18.0 ± 2.6% в серии 2. Таким образом, продолжительность основных форм поведения у интактных и оперированных оленьков была одинаковой, и более 80% времени животные были неактивны.
Рис. 1.
(а) Динамика стадий сна и бодрствования и состояния глаз у малого оленька в течение одного дня. Состояние правого (П) и левого (Л) глаза: О – открыт, З – закрыт. (б) Электрополиграмма АБ, СБ, МС и РЕМ сна длительностью 20 мин. Электроэнцефалограмма (ЭЭГ), электроокулограмма (ЭОГ), электромиограмма (ЭМГ) и ускорение (УСК) во время эпизода, обозначенного пунктирной линией на (а). Стрелки над шкалой времени – время фотографирования. (в–е). Фотографии: АБ (в), МС в позе сфинкса (глаза открыты, отражают ИК-свет) (г, д), РЕМ сон в позе лежа (голова на земле, направленный на камеру глаз закрыт) (е). Темное время суток с 18 до 06 ч.

Электрополиграммы активного и спокойного бодрствования (АБ и СБ), медленноволнового сна (МС) и РЕМ сна у оленьков не отличались от таковых у других копытных (рис. 1б). На бодрствование приходилось 46.2 ± 2.9% времени суток, т.е. меньше половины всего дня. Почти 60% этого времени занимало СБ, во время которого в ЭЭГ на фоне низкоамплитудной (характерной для АБ и СБ) активности регистрировались вспышки с частотой от 5 до 14 Гц.
В ночное время оленьки спали в основном на открытом пространстве вольера, днем – как правило, в домике. МС характеризовался высокоамплитудными медленными волнами в ЭЭГ с частотой ниже 6 Гц и сниженным мышечным тонусом (рис. 1). На МС приходилось 49.7 ± 3.7% времени суток. Практически весь МС (96.2 ± 2.2%) регистрировался в позе сфинкса и лишь небольшое количество – стоя (3.7 ± 2.2%). Большую часть МС глаза у оленьков были открыты. Состояние одного или двух глаз было определено для 47.6 ± ± 13.4% эпох МС. В среднем 87.0 ± 4.4% этого времени глаза были открыты. В дневное время МС сопровождался частым (3–5 раз/сек) поверхностным дыханием (тепловая одышка) и обильным слюноотделением.
РЕМ сон характеризовался практически непрерывными движениями глаз, еще более низким, по сравнению с МС, мышечным тонусом (вплоть до атонии) и низкоамплитудной ЭЭГ, характерной для состояния бодрствования (рис. 1б). У всех оленьков в ЭЭГ увеличивалась активность в диапазоне тета-ритма (частота 6–7 Гц). В РЕМ сне глаза могли быть закрыты или открыты. Часто веки закрывались и открывались параллельно вздрагиваниям глаз. Продолжительность РЕМ сна у разных животных варьировала от 1.2 до 2.4% времени суток и составляла в среднем 1.7 ± 0.3%, или 3.2% от общего количества сна. Всего 35.3 ± 6.4% РЕМ сна было зарегистрировано в позе сфинкса и 64.7 ± 6.4% в позе лежа (голова на поверхности земли). Длительность эпизодов составляла 2.0 ± ± 0.2 мин и варьировала от 20 сек до 8 мин. В день регистрировалось 12.3 ± 1.5 эпизодов РЕМ сна.
Жвачка (ЖВ, ритмичное пережевывание пищи) регистрировалась у оленьков в СБ и МС. Всего на ЖВ приходилось 17.0 ± 3.7% времени суток: 68.3 ± 9.1% всей ЖВ регистрировалось во время МС и 26.8 ± 7.1% – в СБ. Как и у других копытных [1, 2], ЖВ у оленьков никогда не совпадала по времени с РЕМ сном.
Эпизоды бодрствования и сна у оленьков регистрировались в течение всего дня (рис. 2). В обеих сериях животные были наиболее активны перед рассветом (04–06 ч, до 60% времени) и сразу после заката (18–19 ч, 36%), наименее активны – утром (07–09 ч, 10%) и во второй половине дня (13–17 ч, 8%). Наибольшее количество времени животные проводили в МС в утренние часы (07–09 ч, до 71%), наименьшее – перед рассветом (05–06 ч, 15% времени). В период с 9 ч утра до 5 ч утра следующих суток среднее почасовое количество МС постепенно снижалось. Почасовое количество АБ и МС у оленьков значимо зависело от времени суток (АНОВА с повторениями, p < 0.001 и p = 0.008 соответственно). Эпизоды РЕМ сна совпадали с периодами наибольшего количества МС (08–16 и 19–02 ч).
Рис. 2.
Усредненное почасовое количество АБ, МС и РЕМ сна у оленьков (среднее ± S.E.M.). (а) Серия 1 – поведенческие исследования (3 животных, 4–5 дней у каждого), АБ. (б–г) Серия 2 – электрофизиологические исследования (4 животных, 1 день у каждого). (б) АБ, (в) МС, (г) РЕМ сон. Темное время суток с 18 до 06 ч.

В результате исследований мы установили, что стадии МС и РЕМ сна у оленьков хорошо дифференцированы, как и у других плацентарных млекопитающих. При этом общее количество МС (49.7% от времени суток или почти 12 ч) у самого маленького из копытных оказалось самым большим среди всех исследованных копытных, а количество РЕМ сна (1.7% от времени суток или 25 мин) – сопоставимым с величинами у других видов [1–5, 8] (рис. 3). Рекордное количество МС у оленьков согласуется с общей тенденцией увеличения продолжительности сна у копытных с уменьшением веса (рис. 3а). Считается, что более крупные травоядные животные должны тратить больше времени на поиск и поедание пищи, что сокращает время для сна. Разнообразная диета оленьков (листья, плоды, грибы, а иногда и насекомые) и небольшие потребности в количестве корма [6] оставляют им больше времени для сна. С другой стороны, низкий уровень активности и большое количество МС делают оленьков также менее заметными для диких кошачьих, для которых мелкие копытные составляют значительную долю добычи. Поэтому главная стратегия противодействия хищникам у оленьков – это затаивание [6, 7]. Наконец, высокая дневная температура воздуха в тропическом лесу (больше 30°С) и узкая термонейтральная зона у оленьков (15–30°С) также могут быть связаны с большим количеством МС и СБ. Интенсивная двигательная активность оленьков в таких условиях приводит к гипертермии [9]. Таким образом, большая продолжительность МС у оленьков согласуется с представлением о сне как о состоянии адаптивной неподвижности, которое снижает или исключает активность животных, когда она нецелесообразна [8, 10].
Рис. 3.
Соотношение между количеством МС (а) и РЕМ сна (б) и массой тела у копытных. Темно- и светло-синие кружки – дикие и домашние парнокопытные (Artiodactyla) соответственно. Треугольники – непарнокопытные (Perissodactyla). Данные о количестве сна из работ [1–5, 8]. R2 – коэффициент корреляции, p – уровень значимости.
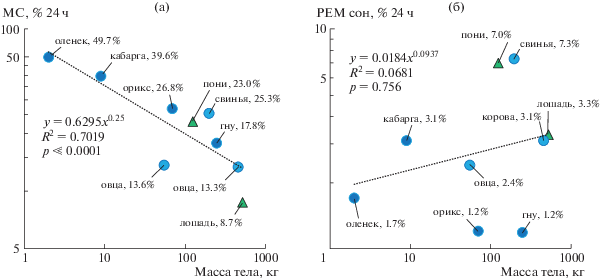
Небольшое количество РЕМ сна у копытных также часто связывают с тем, что они являются добычей для диких кошек [6]. РЕМ сон сопровождается атонией или сниженным мышечным тонусом, и высокими порогами пробуждения [8, 11, 12]. Оба фактора могут замедлять реакцию животных на опасность. Продолжительность РЕМ сна у оленьков, как и двух других диких видов, исследованных в условиях максимально приближенных к естественным, была самой низкой среди копытных (рис. 3б). С другой стороны, наибольшее количество РЕМ сна было зарегистрировано у домашних животных – пони и свиней [2], одним из основных принципов селекции которых было формирование спокойного отношения к человеку. Следовательно, небольшое количество РЕМ сна у оленьков согласуется с данными о связи между продолжительностью РЕМ сна и защищенностью животных, а также уровнем бдительности во время сна.
Считается, что оленьковые ориентируются в первую очередь на обоняние и слух, тогда как стадные копытные для обнаружения хищников используют зрение и коллективную бдительность [6, 7, 13]. Мы установили, что большую часть времени оленьки спали на открытом пространстве с открытыми глазами. Поэтому есть основания предполагать, что оленьки способны обрабатывать зрительную информацию во время МС, что также может способствовать более высокой бдительности. Интересно, что у пони и свиньи при рекордной для копытных продолжительности РЕМ сна глаза во время МС были закрыты [1, 2]. В совокупности эти данные поддерживают идею о том, что МС с открытыми глазами и небольшое количество РЕМ сна – признаки более настороженного состояния животных.
Небольшое количество РЕМ сна у оленьков может быть связано также с высокой температу-рой воздуха и особенностями терморегуляции животных. В отличие от МС, РЕМ сон характеризуется нарушением терморегуляции. Так, учащенное дыхание у оленьков (один из способов понижения температуры тела) регистрировалось во время СБ и МС, но прекращалось за некоторое время до начала эпизода РЕМ сна и восстанавливалось уже после пробуждения. Во время РЕМ сна у животных также растет температура мозга [14]. Хотя мы не обнаружили у оленьков отличий в количестве и длительности эпизодов дневного и ночного РЕМ сна, но все наиболее продолжительные эпизоды РЕМ сна (4–8 мин) были зарегистрированы у них до 9 ч утра или после 16 ч, т.е. вне наиболее жаркого времени суток. К тому же у оленьковых отсутствует система проточного охлаждения поступающей к мозгу крови, которая есть у других видов парнокопытных (так называемая “чудесная сеть” [15]). Таким образом, сниженное количество РЕМ сна может способствовать не только повышению бдительности животных, но и снижению риска перегрева мозга.
Учитывая, что хищники в тропическом лесу могут быть активны в разное время суток [6, 13], сумеречная (крепускулярная) активность оленьков может быть стратегией максимально увеличить активность в периоды наиболее комфортного (с точки зрения температурных условий) времени суток. При этом полифазный сон, а также снижение степени цикличности в виде чередования эпизодов МС и РЕМ сна, делает поведение оленьков пластичным, изменяющимся в зависимости от внешних условий (например, режима активности хищников).
Таким образом, проведенное нами исследование особенностей сна и циркадной ритмики малого оленька в условиях, максимально приближенных к естественным, показало, что эти самые мелкие копытные большую часть времени суток находятся в состоянии покоя и МС, причем МС сопровождается преимущественно открытым состоянием глаз. Всего на МС у оленьков приходится около половины времени суток (наибольшее количество среди исследованных копытных), а РЕМ сон очень краток (как у диких видов). Другие важные особенности оленьков – сумеречный характер активности, полифазный сон, сниженная цикличность стадий сна. Мы не обнаружили каких-либо признаков связи между параметрами сна у оленьков и их филогенетическим статусом. Полученные нами данные предполагают, что особенности цикла сон–бодрствование у оленьков определяются в первую очередь экологическими факторами, такими как характер питания, температурные условия и воздействие хищников, а также размерами тела и особенностями физиологии оленьков.
Список литературы
Ruckebusch Y. // Anim Behav. 1972. V. 20. P. 637–643.
Dallaire A., Ruckebusch Y. // Can. J. Comp. Med. 1974. V. 38. № 1. P. 65–67.
Соколов В.Е., Мухаметов Л.М., Приходько В.И., и др. // Доклады Академии Наук СССР. 1988. Т. 302. № 4. С. 1005–1009.
Davimes J.G., Alagaili A.N., Bhagwandin A. et al. // Sleep. 2018. V. 41. № 5.
Malungo I.B., Gravett N., Bhagwandin A., et al. // IBRO Neuroscience Reports. 2021. V. 10. P. 142–152.
Meijaard E. In: Handbook of the Mammals of the World. V. 2. Hoofed Mammals. Madrid: Lynx Editions, 2011. P. 320–335.
Janis C. In: Living fossils. Springer-Verlag: New York,1984. P. 87–94.
Siegel J.M. // Nature. 2005. V. 437. № 7063. P. 1264–1271.
Whittow G.C., Scammell C.A., Leong M. et al. // Comp. Biochem. Physiol. Part A. 1977. V. 56. № 1. P. 23–26.
Siegel J.M. // Nature Rev Neurosci. 2009. V. 10. № 10. P. 747–753.
Twyver V.H., William G. // Behavioral biology. 1972. V. 7. № 2. P. 205–215.
Ermis U., Krakow K., Voss U. // J. Sleep Res. 2010.V. 19. № 3. P. 400–406.
Рожнов В.В. Опосредованная хемокоммуникация в социальном поведении млекопитающих. М.: Т-во научных изданий КМК. 2011. 288 с.
Kovalzon V.M. // Physiol Behav. 1973. V. 10. № 4. P. 667–670.
Fukuta K., Kudo H., Sasaki M., et al. // J. Anat. 2007. V. 210. № 1. P. 112–116.
Дополнительные материалы отсутствуют.
Инструменты
Доклады Российской академии наук. Науки о жизни