

Доклады Российской академии наук. Науки о жизни, 2022, T. 502, № 1, стр. 60-65
GIGANTOPITHECUS BLACKI (PRIMATES, PONGINAE) ИЗ ПЕЩЕРЫ ЛАНГЧАНГ (СЕВЕРНЫЙ ВЬЕТНАМ): ПОСЛЕДНИЕ ГИГАНТОПИТЕКИ В ПОЗДНЕМ ПЛЕЙСТОЦЕНЕ?
Академик РАН А. В. Лопатин 1, *, Е. Н. Мащенко 1, 2, Ле Суан Дак 3
1 Палеонтологический институт им. А.А. Борисяка Российской академии наук
Москва, Россия
2 Совместный Российско-Вьетнамский тропический научно-исследовательский и технологический центр
Ханой, Вьетнам
3 Институт тропической экологии Совместного Российско-Вьетнамского тропического научно-исследовательского и технологического центра
Ханой, Вьетнам
* E-mail: alopat@paleo.ru
Поступила в редакцию 20.07.2021
После доработки 21.09.2021
Принята к публикации 21.09.2021
- EDN: HLZJVH
- DOI: 10.31857/S2686738922010140
Аннотация
Из верхнеплейстоценовых отложений пещеры Лангчанг в северном Вьетнаме (провинция Тханьхоа) описаны зубные остатки (целый правый m2 с корнями и фрагмент дистальной части коронки левого m2) Gigantopithecus blacki von Koenigswald, 1935. Это первая находка гигантопитеков в верхнем плейстоцене Вьетнама и, по-видимому, вторая – в верхнем плейстоцене в целом (с учетом материалов из пещеры Шуантань в южном Китае). Вероятно, вымирание G. blacki было приурочено к переходу от среднего к позднему плейстоцену, и экземпляры из пещеры Лангчанг принадлежат к одной из самых поздних, реликтовых, популяций этого вида.
Гигантопитек Gigantopithecus blacki von Koenigswald, 1935 – крупнейший примат за всю историю отряда. Этот вид гигантских человекообразных обезьян, родственный орангутану, был довольно широко распространен в плейстоцене на территории южного Китая [1], северного Вьетнама [2] и северного Таиланда [3]. Ископаемые остатки гигантопитеков приурочены к отложениям карстовых пещер и представлены в основном изолированными зубами и фрагментами челюстей. Для нижнего и среднего плейстоцена (возраст местонахождений от 2.0 до 0.3 млн лет) находки гигантопитеков довольно обычны [1], а для верхнего плейстоцена – единичны [4, 5] и альтернативно датируются концом среднего плейстоцена [1, 6]. Поэтому значительный интерес представляет описанная в настоящей статье находка двух изолированных зубов G. blacki в позднеплейстоценовом местонахождении Лангчанг (Lang Trang) в северном Вьетнаме (провинция Тханьхоа, уезд Батхыок; географические координаты 20°20'29'' с.ш., 105°12'21'' в.д.) в ассоциации с остатками других млекопитающих. Фаунистический комплекс пещеры Лангчанг датирован началом позднего плейстоцена [см. 7, 8].
Описанный материал происходит из основного зала пещеры, обозначенного как Лангчанг II [9], из костеносной брекчии в юго-восточной части пещеры [см. 9, рис. 2, 3]. Образцы хранятся в Палеонтологическом институте им. А.А. Борисяка РАН (ПИН) в Москве. Измерения в мм, проведены с помощью цифрового штангенциркуля с точностью 0.01 мм и окуляр-микрометра стереомикроскопа Leica MZ6 с точностью 0.05 мм. Внутреннее строение зубов исследовано на рентгеновском микротомографе Neoscan N80 в ПИН, поверхность эмали изучена с помощью сканирующего электронного микроскопа Tescan Vega II XMU в ПИН.
Рис. 1.
Gigantopithecus blacki von Koenigswald, 1935: а–е – экз. ПИН, № 5792/439, правый m2: а – с окклюзиальной стороны, б – с корневой стороны, в – с буккальной стороны, г – с лингвальной стороны, д – с мезиальной стороны, е – с дистальной стороны; ж–л – экз. ПИН, № 5792/490, фрагмент дистальной части левого m2: ж – с окклюзиальной стороны, з – с буккальной стороны, и – с лингвальной стороны, к – с мезиальной стороны (на сломе видна толщина эмали), л – с дистальной стороны; Вьетнам, провинция Тханьхоа, уезд Батхыок, пещера Лангчанг; низы верхнего плейстоцена.
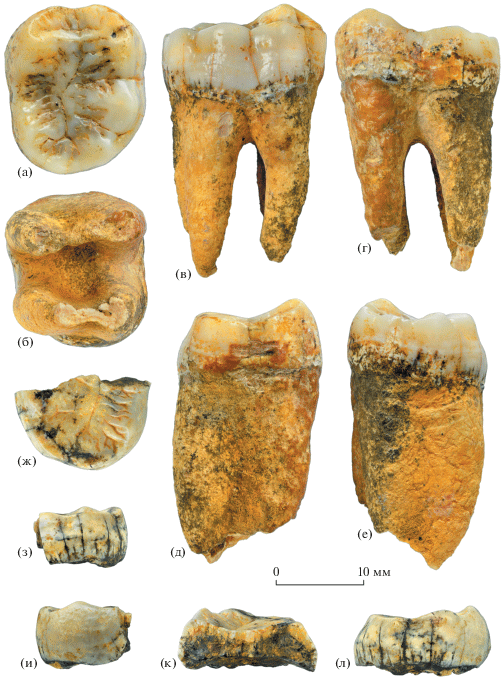
Рис. 2.
Gigantopithecus blacki von Koenigswald, 1935, экз. ПИН, № 5792/439, правый m2, томограмма на уровне передних главных бугорков, демонстрирующая толщину эмали; Вьетнам, провинция Тханьхоа, уезд Батхыок, пещера Лангчанг; низы верхнего плейстоцена.

Рис. 3.
Gigantopithecus blacki von Koenigswald, 1935, экз. ПИН, № 5792/490, фрагмент дистальной части левого m2 с увеличенным участком поверхности эмали с микроцарапинами и ямками; Вьетнам, провинция Тханьхоа, уезд Батхыок, пещера Лангчанг; низы верхнего плейстоцена.

Отряд Primates Linnaeus, 1758
Семейство Hominidae Gray, 1825
Подсемейство Ponginae Elliot, 1913
Род Gigantopithecus von Koenigswald, 1935
Gigantopithecus blacki von Koenigswald, 1935
Gigantopithecus blacki: [10, с. 874, рис. 14 ].
Голотип – Научно-исследовательский институт и музей естественной истории Зенкенберга (Франкфурт-на-Майне, Германия), коллекция Густава Генриха Ральфа фон Кёнигсвальда (GHRvK), № CA 733, изолированный правый m3; Китай, плейстоцен.
Описание (рис. 1–3). Второй нижний постоянный коренной зуб m2 (экз. ПИН, № 5792/439) относительно высококоронковый, длинный и узкий, с отчетливым букколингвальным пережимом посередине длины и заметным дистальным сужением. Зуб относительно слабо стерт, сохранились все элементы рельефа окклюзиальной поверхности. Основные бугорки массивные, гребни низкие и сглаженные. Мезиальная контактная фасетка четкая, дистальная контактная фасетка слабо выражена.
Протоконид и метаконид одинаково крупные, их вершины связаны широким и мощным, но низким в средней части поперечным гребнем. Спереди от этого гребня находится поперечно вытянутая широкая и довольно глубокая мезиальная ямка, а дистально протягивается длинная продольная борозда. Высокий мезиальный краевой гребень ограничивает мезиальную ямку спереди и боковыми концами соединяется с вершинами протоконида и метаконида. Постпротокристид и прегипокристид разделены глубокой поперечной бороздой, расположенной на уровне букколингвального пережима. Постметакристид отделен от преэнтокристида глубокой долинкой, также располагающейся на уровне букколингвального пережима. У лингвального выхода долинки находится очень мелкий дополнительный бугорок, продолжающийся буккально невысоким эмалевым гребнем.
Талонид несколько уже тригонида, талонидный бассейн длинный, узкий и сравнительно неглубокий. Гипоконид немного меньше протоконида. Гипоконулид еще меньше, расположен у заднего края окклюзиальной поверхности несколько буккальнее средней линии коронки. Этот бугорок обособлен от гипоконида короткой направленной дистобуккально бороздой (у ее выхода расположен маленький дополнительный бугорок), а от энтоконида – дистальной порцией длинной продольной борозды. Энтоконид меньше и ниже гипоконида, приблизительно равен по размерам гипоконулиду. Между энтоконидом и гипоконулидом имеется относительно крупный, низкий и плоский дополнительный шестой бугорок (tuberculum sextum), отделенный от соседних бугорков короткими бороздками. В месте соединения этих бороздок с окончанием продольной борозды располагается мелкая треугольная дистальная ямка. Складчатость эмали талонидного бассейна образует мелкие вторичные бугорки и короткие гребни, придающие окклюзиальной поверхности коронки характерный мелкобугорчатый облик.
Корни длинные, массивные, продольно сжатые (рис. 1б–1г); дистальный корень длиннее мезиального.
Экз. ПИН, № 5792/490 (фрагмент дистальной части коронки среднестертого m2) отличается от экз. ПИН, № 5792/439 более низкими бугорками талонида, сильнее выраженной складчатостью талонидного бассейна в области энтоконида, а также более четкой дистальной контактной фасеткой.
Эмаль обоих моляров толстая, особенно в области вершин основных бугорков (рис. 1к, 2). Поверхность эмали с микроцарапинами и ямками, что характерно для зубов гигантопитеков (рис. 3).
Размеры в мм. Экз. ПИН, № 5792/439, правый m2: мезиодистальный диаметр (MD) – 19.11; букколингвальный диаметр (BL) – 16.62; ширина талонида (BT) – 15.85; высота по протокониду (HP) – 8.00; высота по метакониду (HM) – 8.09; высота по гипокониду (HH) – 9.00; высота по энтокониду (HE) – 7.50; высота по гипоконулиду (HHl) – 7.70; длина мезиального корня – 20.71; длина дистального корня – 21.40; толщина эмали по томограмме (методика и терминология измерения по [11]): линейная буккальная бугорковая толщина (BCT, по протокониду) – 2.50, линейная лингвальная бугорковая толщина (LCT, по метакониду) – 2.18, толщина в середине окклюзиального бассейна (MOB) – 1.73.
Экз. ПИН, № 5792/490, фрагмент дистальной части левого m2: BT – 15.55; HH – 5.90; HE – 7.00; HHl – 5.60; толщина эмали на сломе (рис. 1к): по гипокониду – 1.7, по преэнтокристиду – 1.2, по средней линии талонидного бассейна – 1.35.
Замечания. Позднемиоценовый Gigantopithecus bilaspurensis Simons et Chopra, 1969 из Индии в настоящее время считается младшим синонимом Indopithecus giganteus (Pilgrim, 1915) [12]. Обнаруженные на Яве (Индонезия) два фрагмента нижних челюстей крупного примата неопределенного геологического возраста, соответствующие по челюстным и зубным промерам G. blacki, рассматриваются как Gigantopithecus sp. [13, 14].
Метрические показатели зубов из местонахождения Лангчанг (экз. ПИН, № 5792/439: MD – 19.11, BL – 16.62; площадь, MD×BL – 317.61 мм2) приблизительно соответствуют средним значениям для серий m1/2 G. blacki (минимум – среднее – максимум: MD – 15.90–19.77–26.92 мм, 205 экз.; BL – 13.31–17.47–24.67 мм, 201 экз.; MD×BL – 222.1–349.06–647.2 мм2, 194 экз.) [1]. По всему комплексу одонтологических признаков экземпляры из Лангчанга полностью отвечают G. blacki. Размеры и форма зубных корней у экз. ПИН, № 5792/439 – характерные для G. blacki [1, 15], так же как толщина эмали и характер ее микростирания у обоих изученных экземпляров [11, 16]. Не отмечено морфологических признаков, отличающих образцы из Лангчанга от материала из других местонахождений гигантопитеков и показывающих эволюционные вариации у представителей вида разных хронологических интервалов.
Распространение. Нижний – низы верхнего плейстоцена Китая, средний – низы верхнего плейстоцена Вьетнама.
Материал. Из пещеры Лангчанг два изолированных зуба: целый правый m2 с корнями (экз. ПИН, № 5792/439) и фрагмент дистальной части коронки левого m2 (экз. ПИН, № 5792/490).
В Китае Gigantopithecus blacki известен из 18 местонахождений (в областях Хубэй, Гуйчжоу, Хайнань, Гуанси, Чунцин), расположенных на территории от р. Янцзы до Южно-Китайского моря [1, 14; и др.]. Считается, что вид вымер в конце среднего плейстоцена [16]. Фактически, позднеплейстоценовые находки G. blacki в Китае представлены лишь пятью зубами из пещеры Шуантань (Shuangtan) на горе Мулань в провинции Хубэй [5]. Позднеплейстоценовый возраст этого местонахождения был обоснован совместным присутствием остатков человека Homo sapiens Linnaeus, 1758 и азиатского слона Elephas maximus Linnaeus, 1758 [4]. В ряде современных публикаций для местонахождения в пещере Шуантань без каких-либо комментариев указывается среднеплейстоценовый возраст [1, 6], но при этом некоторые из авторов позднеплейстоценовой датировки продолжаютее придерживаться и в настоящее время [14].
Из пещеры Лангчанг известны остатки более 30 видов млекопитающих фауны Stegodon–Ailuropoda, характерной для второй половины среднего плейстоцена и начала позднего плейстоцена [8, 17]. Возраст лангчангской фауны определялся в интервалах 385–185 тыс. л.н. [9, 18] или 80–60 тыс. л.н. [17]; в последнее время он уверенно соотносится с началом позднего плейстоцена – 100–80 тыс. л.н. [7, 8]. В ее составе много рецентных видов [8, 18], но отсутствуют вымершие формы, свойственные среднему плейстоцену (теперь исключая G. blacki). Среди парнокопытных присутствуют виды, характерные для болотистых местообитаний и мангровых зарослей (бородатая свинья Sus barbatus Müller, 1838, малый канчиль Tragulus kanchil (Raffles, 1821), свиной олень Axis porcinus (Zimmermann, 1780), водяной олень Hydropotes inermis Swinhoe, 1870) [8]. Комплекс брюхоногих моллюсков (определение П.Ю. Пархаева, ПИН) включает наземные и пресноводные формы и один прибрежно-морской вид – Ellobium aurismidae (Linnaeus, 1758). Последний в настоящее время населяет прибрежные мангры и эстуарии рек. Присутствие этого вида позволяет синхронизировать время накопления изученных отложений с эпизодом крупной морской трансгрессии, когда море подходило к району пещеры (в настоящее время он удален от морского побережья более чем на 90 км, его высота над уровнем моря составляет около 60 м). В целом мы полагаем, что возраст изученной фауны из местонахождения Лангчанг близок времени наиболее высокого в позднем плейстоцене подъема уровня моря (MIS 5e, ~125 тыс. л.н.) [19] и, таким образом, соответствует самому началу позднего плейстоцена.
Судя по географическому и стратиграфическому распространению ископаемых находок, уже в среднем плейстоцене ареал гигантопитека сократился до небольшого района на смежных территориях южного Китая и северного Вьетнама [16]; к концу среднего плейстоцена он мог распасться на отдельные рефугиумы. Можно предположить, что вымирание вида было приурочено к переходу от среднего к позднему плейстоцену, и образцы из пещеры Лангчанг принадлежат к одной из самых поздних, реликтовых, популяций гигантопитеков.
Список литературы
Zhang Y., Harrison T. Gigantopithecus blacki: a giant ape from the Pleistocene of Asia // American Journal of Physical Anthropology. 2017. V. 162 (S63). P. 153–177.
Ciochon R., Vu The Long, Larick R., et al. Dated co-occurrence of Homo erectus and Gigantopithecus from Tham Khuyen Cave, Vietnam // Proceedings of the National Academy of Sciences. 1996. V. 93. № 7. P. 3016–3020.
Bocherens H., Schrenk F., Chaimanee Y., et al. Flexibility of diet and habitat in Pleistocene South Asian mammals: implications for the fate of the giant fossil ape Gigantopithecus // Quaternary International. 2017. V. 434. Pt A. P. 148–155.
Jin C., Takai M., Zhang Y., Kono R.T. Sequence of Gigantopithecus faunas, from Chongzuo, Guangxi, South China // The 2012 Annual Meeting of the Paleontological Society of Japan, June 29–July 1, 2012, Nagoya, Aichi Prefecture. Abstracts with programs. Paleontological Society of Japan, 2012. P. 18.
Takai M., Zhang Y., Kono R.T., Jin C. Changes in the composition of the Pleistocene primate fauna in southern China // Quaternary International. 2014. V. 354. P. 75–85.
Ortiz A., Zhang Y., Jin C., et al. Morphometric analysis of fossil hylobatid molars from the Pleistocene of southern China // Anthropological Science. 2019. V. 127. № 2. P. 109–121.
Suraprasit K., Jaeger J.-J., Chaimanee Y., et al. The Middle Pleistocene vertebrate fauna from Khok Sung (Nakhon Ratchasima, Thailand): biochronological and paleobiogeographical implications // ZooKeys. 2016. V. 613. P. 1–157.
Лопатин А.В., Мащенко Е.Н., Вислобокова И.А., и др. Плейстоценовые млекопитающие из пещеры Лангчанг (Вьетнам): новые данные // Доклады Российской академии наук. Науки о жизни. 2021. Т. 496. № 1. С. 5–9.
Ciochon R.L., Olsen J.W. Paleoanthropological and archaeological discoveries from Lang Trang caves: a new Middle Pleistocene hominid site from northern Viet Nam // Bulletin of the Indo-Pacific Prehistory Association. 1991. V. 10. P. 59–73.
Koenigswald G.H.R von. Eine fossile Säugetierfauna mit Simia aus Südchina // Proceedings of the Koninklijke Akademie van Wetenschappen. 1935. V. 38. № 8. P. 872–879.
Dean M.C., Schrenk F. Enamel thickness and development in a third permanent molar of Gigantopithecus blacki // Journal of Human Evolution. 2003. V. 45. P. 381–387.
Cameron D. The taxonomic status of the Siwalik late Miocene hominid Indopithecus (= Gigantopithecus) // Himalayan Geology. 2001. V. 22. P. 29–34.
Noerwidi S., Siswanto S., Widianto H. Giant primate of Java: a new Gigantopithecus specimen from Semedo // Berkala Arkeologi. 2016. T. 36. № 2. P. 141–160.
Insani H., Takai M. Mainland versus island adaptation: paleobiogeography of Sunda shelf Primates revisited. In: Ono R., Pawlik A., eds. Pleistocene archaeology – migration, technology, and adaptation. London: IntechOpen; 2020. P. 1–22.
Kupczik K., Dean M.C. Comparative observations on the tooth root morphology of Gigantopithecus blacki // Journal of Human Evolution. 2008. V. 54. № 2. P. 196–204.
Zhao L.X., Zhang L.Z. New fossil evidence and diet analysis of Gigantopithecus blacki and its distribution and extinction in South China // Quaternary International. 2013. V. 286. P. 69–74.
Long V.T., De Vos J., Ciochon R.L. The fossil mammal fauna of the Lang Trang caves, Vietnam, compared with Southeast Asian fossil and recent mammal faunas: the geographical implications // Bulletin of the Indo-Pacific Prehistory Association. 1996. V. 14. P. 101–109.
Ciochon R.L. Divorcing hominins from the Stegodon-Ailuropoda fauna: new views on the antiquity of hominins in Asia. In: Fleagle J.G., Shea J.J., Grine F.E., et al., eds. Out of Africa I: the first hominin colonization of Eurasia. Dordrecht, Heidelberg, London, New York: Springer; 2010. P. 111–126.
Pedoja K., Shen J.-W., Kershaw S., Tang C. Coastal Quaternary morphologies on the northern coast of the South China Sea, China, and their implications for current tectonic models: a review and preliminary study // Marine Geology. 2008. V. 255. P. 103–117.
Дополнительные материалы отсутствуют.
Инструменты
Доклады Российской академии наук. Науки о жизни