

Доклады Российской академии наук. Науки о жизни, 2022, T. 506, № 1, стр. 387-391
РАЗРАБОТКА ИНСТРУМЕНТАЛЬНОГО МЕТОДА ОПРЕДЕЛЕНИЯ ТИПОВ ПОВЕДЕНИЯ БЕЛУХИ (DELPHINAPTERUS LEUCAS) ПО ПАРАМЕТРАМ УСКОРЕНИЯ
О. И. Лямин 1, 2, *, Е. А. Назаренко 1, академик РАН В. В. Рожнов 1
1 Институт проблем экологии и эволюции
им. А.Н. Северцова Российской академии наук
Москва, Россия
2 Калифорнийский университет в г. Лос-Анджелес
Лос-Анджелес, США
* E-mail: oilyamin@yahoo.com
Поступила в редакцию 06.07.2022
После доработки 10.07.2022
Принята к публикации 10.07.2022
- EDN: MCJCPE
- DOI: 10.31857/S2686738922050183
Аннотация
Апробирован неинвазивный инструментальный метод описания поведения белухи (Delphinapterus leucas) по параметрам двигательной активности, которую характеризовали средним и стандартным отклонением ускорения хвостового стебля животного. Было установлено, что с помощью алгоритма “дерева решений” могут быть выделены как минимум два типа поведения белух – спокойное зависание у поверхности воды и активное плавание по бассейну. Точность определения (в зависимости от параметра) составляла для спокойного зависания в среднем 81–97%, а для активного плавания – 54–99%. Различия в точности классификации спокойного зависания по 3Д-ускорению и результирующему ускорению были минимальными. В будущем точность может быть повышена использованием других параметров ускорения, уменьшением интервала анализа и оптимизацией алгоритмов классификации.
Неинвазивные инструментальные методы оценки поведения животных имеют большой потенциал для исследований в области их экологической физиологии и поведенческой экологии, а также для мониторинга состояния здоровья животных [1, 2]. Одним из параметров, который характеризует поведение, является двигательная активность. Для ее описания используются акселерометры, регистрирующие ускорение в трех перпендикулярных направлениях. Последние два десятилетия их все чаще применяют в исследованиях поведения морских млекопитающих (китообразных и ластоногих), в частности пищевого поведения [3], реакции на антропогенный шум [4], состояния покоя и сна [5, 6]. Белуха – один из наиболее популярных объектов исследований среди китообразных, в том числе как индикатор состояния арктических экосистем [7]. В природных условиях для изучения поведения этого вида использовали регистраторы глубины погружений и акустической активности, спутниковые метки [7], а в условиях аквариумов и вольерного содержания – методы видеорегистрации [8, 9]. Акселерометрию при исследовании белух применяли крайнее редко [10].
Задача данного исследования состояла в том, чтобы оценить возможность описания поведения белух по параметрам их двигательной активности.
МАТЕРИАЛЫ И МЕТОДЫ
Исследование выполнено на белухе (Delphinapterus leucas) на Утришской морской станции Института проблем экологии и эволюции им. А.Н. Северцова РАН. На момент проведения экспериментов белуха (самец, возраст не менее 10 лет, вес не менее 600 кг) содержалась на станции три года. За три недели до начала эксперимента животное было помещено в круглый бассейн диаметром 6 м и глубиной 1.8 м. На хвостовой стебель белухи установили мягкий обруч с акселерометром (X16-mini, www.gcdataconcepts.com, вес 30 г), который регистрировал ускорение в трех направлениях: ростро-каудальном, дорзо-вентральном и проксимально-дистальном (рис. 1). Вес обруча составлял 500 г (<0.08% массы белухи) и, судя по поведению, не беспокоил животное. Белуху кормили два раза в день.

Эксперимент продолжался 4 дня, в течение которых поведение белухи регистрировали круглосуточно двумя видеокамерами параллельно с двигательной активностью. В поведении визуально выделяли спокойное зависание у поверхности воды (СЗП) и несколько видов активного поведения, включая плавание по бассейну (АП), игру в струе воды при заливе бассейна, а также активность на месте.
Для установления корреляции между визуально определенным типом поведения и параметрами ускорения были использованы видеозаписи и данные акселерометрии третьего дня. Ускорение по каждой из трех осей оцифровывали с частотой 5 гц. За каждый минутный интервал рассчитывали среднее и стандартное отклонение ускорения по каждой из осей (3Д-ускорение), а также результирующее ускорение – корень из суммы квадратов по всем трем осям (общая двигательная активность, ОДА). Корреляцию между типами поведения и параметрами ускорения устанавливали программными средствами библиотеки “rpart” языка программирования R. Процедура реализует метод “дерево решений”, последовательно разбивая данные на группы и добиваясь минимальной суммы квадратов внутригрупповых отклонений [11]. Эффективность классификации оценивали для всех четырех дней (10 ч в день: 2 ч подряд с разрывом в 2 ч; периоды тренировки и смены воды не учитывали) по четырем параметрам, которые применяются для оценки работы систем искусственного интеллекта, а именно: точность (способность определять события данного типа и показатель ложных срабатываний), полнота (показатель пропуска событий и совпадения с экспертной оценкой), специфичность (показатель точности определения событий другого класса), а также по среднему значению для всех трех параметров [12].
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
Поведение белухи было представлено преимущественно СЗП, на которое приходилось 83.0 ± ± 2.5% (n = 4) времени. Суммарная длительность активного поведения составляла 8.4 ± 1.9% времени. Значительная его часть представляла собой активное плавание по бассейну (АП). В остальное время белуха дрейфовала у поверхности или заныривала на месте.
На рис. 2 показана динамика ускорения белухи в течение 7 ч параллельно с визуально определенными типами поведения. В СЗП двигательная активность варьировала от повторяющихся движений хвостового стебля до практически полной неподвижности. Ускорение (по всем трем осям) и ОДА регулярно увеличивались и уменьшались с периодом 4–8 мин, что соответствовало характерному для белух чередованию задержек дыхания (белуха опускала голову под воду) и регулярных вдохов-выдохов (белуха поднимала дыхало над водой для каждого вдоха); вариабельность ускорения в это время была низкой. Такой паттерн активности регистрировали до 10:20, когда в бассейне начинался слив (смена) воды. В СЗП выделялись два эпизода с минимальным ускорением и стандартным отклонением (03:50–04:10 и 05:50–06:40); параллельно снижались те же параметры ОДА. Такие эпизоды могли быть глубоким сном [13]. Таким образом, акселерометрия позволяет идентифицировать у белухи периоды покоя с разным уровнем двигательной активности и оценить продолжительность задержек дыхания.
Рис. 2.
Динамика параметров ускорения и типы поведения у белухи. Ускорение – средние значения (Xm, Ym, Zm,) и стандартное отклонение (Xsd, Ysd, Zsd) по трем осям и результирующие значения (ОДАm и ОДАsd; м/c2, относительные величины). Типы поведения определялись визуально (Видео) и классификатором по 3Д-ускорению и ОДА. Все параметры рассчитаны для одноминутных интервалов. АП – активное плавание, С – активность во время залива бассейна, З – заныривание, СЗП – спокойное зависание у поверхности, ЛА – активность во время слива воды.
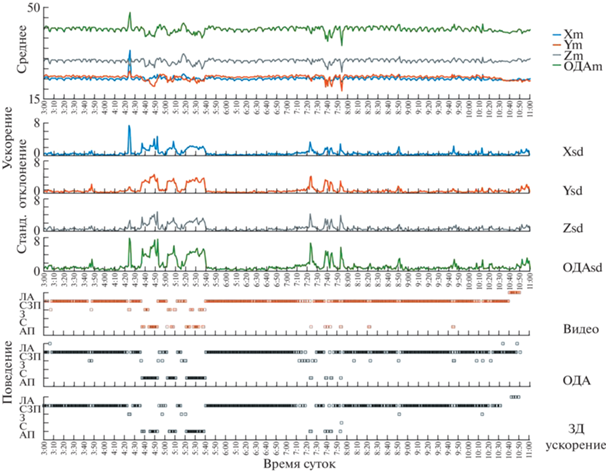
АП регистрировалось в периоды 04:20–05:40 и 07:20–08:00 (рис. 2). АП характеризовалось большим ускорением и большой его вариабельностью по всем трем осям. Параллельно увеличивались ОДА и его вариабельность. Несколько эпизодов заныриваний совпали с “взрывным” ростом ускорения только по вертикальной оси (например, в 04:30). Это означает, что и другие типы активного поведения белух могут быть выделены по 3Д-ускорению.
По трехкомпонентной оценке (точность, полнота и специфичность), общая эффективность классификации СЗП по 3Д-ускорению и ОДА была одинаковой – в среднем 90% (табл. 1). При этом показатель точности определения СЗП составил 97% (ложных срабатываний всего 3%), полноты – по 3Д-ускорению 89%, по ОДА 94% (пропусков меньше 11%), а специфичность была выше 80% (способность определять поведение, которое не было СЗП). Точность оценки реального количество СЗП по ОДА достигла 98% всего СЗП, определенного визуально, что было даже больше, чем по 3Д-ускорению (92%; p = 0.003, парный Т-тест).
Таблица 1.
Результаты классификации двух категорий поведения белухи (спокойного зависания у поверхности воды и активного плавания) визуально и по параметрам ускорения и метрики оценки эффективности классификации
| Параметр | Спокойное зависание у поверхности (СЗП) | Активное плавание (АП) | ||
|---|---|---|---|---|
| 3Д | ОДА | 3Д | ОДА | |
| Визуальная оценка | 3.5 ± 1.3 | – | ||
| Классификатор | 77.0 ± 3.0 | 81.7 ± 2.7 * | 4.2 ± 1.3 | 5.5 ± 1.9 |
| Классификатор/Визуально | 92.1 ± 1.7 | 97.8 ± 1.5 * | 131.2 ± 17.1 | 160.5 ± 5.2 |
| Точность/precision (Т) | 96.8 ± 0.9 | 96.1 ± 1.0 | 66.3 ± 5.3 | 53.6 ± 1.5 |
| Полнота/recall (П) | 89.1 ± 1.3 | 94.0 ± 0.9* | 84.7 ± 6.7 | 85.8 ± 2.4 |
| Специфичность/specificity (С) | 84.5 ± 4.9 | 80.5 ± 6.1 | 98.5 ± 0.5 | 97.4 ± 0.8 |
| Общая оценка (ТПС/3) | 90.1 ± 1.7 | 90.2 ± 2.2 | 83.1 ± 2.6 | 78.9 ± 0.9 |
Данные представлены как среднее и ошибка измерений для 4 дней. Расчеты сделаны для 3Д-ускорения и ОДА. * – значимость отличий между средними не менее 0.01 (парный Т-тест). Формулы расчета параметров оценки: Т = ИП/(ИП + ЛП), П = = ИП/(ИП + ЛО), С = ИО/(ИО+ЛП), С = (Т + П + С)/3. ИП – истинно положительный результат, ИО – истинно отрицательный, ЛП – ложно положительный, ЛО – ложно отрицательный.
Некоторые из использованных параметров оценки зависят от числа событий [12]. Тем не менее значимой корреляции между количеством СЗП в разных сутках (79–89% времени регистрации) и метриками оценки не было (p > 0.05, коэффициент корреляции Пирсона). Таким образом, эпизоды СЗП у белух могут быть достаточно надежно определены как по 3Д-ускорению, так и ОДА.
АП у белухи определялось хуже, чем СЗП: в среднем на 7–10% (табл. 1). Главным образом это было связано с уменьшением точности (ростом числа ложных срабатываний на 34% для ЗД-ускорения и на 46% для ОДА). Способность классификатора определять общее количество АП также ухудшилась: по 3Д-ускорению превышение реального количества АП составило 46%, а по ОДА – 80%. В то же время специфичность классификации АП по сравнению с СЗП увеличилась примерно на 15%.
В наших условиях количество АП у белухи было в среднем в 20 раз меньше, чем СЗП, а межсуточные различия составляли 7 раз. Мы не исключаем, что точность определения АП была бы выше при большей продолжительности АП, например, если бы белуха находилась в более просторном бассейне или в открытом море, и где она была бы более активна. В то же время значимая корреляция была обнаружена только между количеством АП и значением параметра специфичность (Rxy = –0.988, p = 0.01) при оценке по ОДА.
Таким образом, мы установили, что, используя алгоритм “дерева решений”, эпизоды СЗП у белухи могут быть определены как по параметрам 3Д-ускорения, так и ОДА (что упрощает расчеты) с надежностью 81–97% в зависимости от параметра оценки. СЗП – один из основных типов поведения китообразных, продолжительность которого характеризует скорость метаболизма, уровня стресса и состояния здоровья животного [14]. Для описания поведения животных предсказательная точность классификатора более 90% считается высокой [4, 15]. В то же время выявление у белухи АП было менее точным (56–98%), а оценка суммарной длительности – завышенной. Такие результаты могли определяться достаточно “простыми” параметрами оценки двигательной активности (среднее и стандартное отклонение ускорения) и небольшим количеством АП. Немаловажно, что в нашем эксперименте была использована и простая модель акселерометра. Будущие исследования должны быть направлены на оптимизацию алгоритмов классификации, включая использование других параметров движения (например, доминирующей частоты в ускорении), сокращение интервала анализа (от 1 мин до 10–20 с), а также применение более чувствительных акселерометров.
Список литературы
Whitford M., Klimley A.P. // Anim. Biotelemetry. 2019. V. 7. P. 1–26.
Wilmers C.C., Nickel B., Bryce C.M. et al. // Ecology. 2015. V. 96. № 7. P. 1741–1753.
Tennessen J.B., Holt M., Hanson M.B. et al. // J. Exp. Biol. 2019. V. 222 (3). jeb191874.
Holt M.M., Tennessen J.B., Hanson M.B. et al.// Marine Envir. Research. 2021. V. 170. № 105429.
Miller P.J.O., Aoki K., Rendell L.E. et al. // Current Biology. 2008. V. 18. № 1. P. R21–R23.
Mitani Y., Andrews R.D., Sato K. et al. // Biol. Lett. 2010. V. 6. № 2. P. 163–166.
O'Corry-Crowe G. In. Encyclopedia of Marine Mammals. 2009. P. 108–111.
Lyamin O.I., Shpak O.V., Nazarenko E.A. et al. // Physiol. Behav. 2002. V. 76. № 2. P. 265–270.
Lyamin O.I., Ostras D.A., Mukhametov L.M. et al. // Proceedings of Meetings on Acoustics. 2016. V. 27. № 050002.
Castellote M., Mooney A., Andrews R. et al. // PLoS ONE. 2021. V. 16. № 11. e0260485.
James G., Witten D., Hastie T. et al. An Introduction to Statistical Learning with Applications in R. Springer Science. New York. 2013. P. 426.
Hossin M., Sulaiman M.N. // Intern. J. Data Mining & Knowledge Management Process. 2015. V. 5. № 2 (1).
Lyamin O.I., Manger P.R., Ridgway S.H. et al. // Neurosci. Biobehav. Review. 2008. V. 32. № 8. P. 1451–1484.
Qasem L., Cardew A., Wilson A. et al. // PLoS ONE. 2012. V. 7. e31187.
Graf P.M., Wilson R.P., Qasem L. et al. // PLoS One. 2015. V. 10. № 8: e0136751.
Дополнительные материалы отсутствуют.
Инструменты
Доклады Российской академии наук. Науки о жизни