

Доклады Российской академии наук. Науки о жизни, 2022, T. 507, № 1, стр. 510-514
ЭНТРОПИЯ И ЭФФЕКТИВНОСТЬ РОСТА У МОРСКИХ ВИДОВ РЫБ АРКТИКИ
1 Мурманский морской биологический институт Российской академии наук (ММБИ РАН)
Мурманск, Россия
* E-mail: karamushkol@yahoo.com
Поступила в редакцию 30.06.2022
После доработки 10.07.2022
Принята к публикации 11.07.2022
- EDN: BPVICP
- DOI: 10.31857/S2686738922060105
Аннотация
На основании собственных экспериментальных данных и литературных сведений исследованы механизмы процессов обмена энергией у рыб, обитающих в высокоширотных районах Арктики. Показано, что потребление энергии в расчете на единицу роста становится тем экономнее, чем медленнее они растут (низкая удельная скорость роста). В этом случае процесс роста у рыб при более низких температурах становится все более эффективным, и они могут расти с минимальными затратами энергии. При низких температурах внутренние процессы замедляются и отходы накапливаются медленнее, что приводит к высвобождению меньшего количества лишнего тепла, которое генерируется метаболическими процессами и ежедневной деятельностью. У пойкилотермных животных эта стратегия использования низкоэнтропийной энергии и “сбрасывания” высокоэнтропийной теплоты при низких температурах весьма эффективна.
При исследовании энергетики пойкилотермных водных организмов главным определяющим термодинамическим параметром является температура, которая, влияя на скорость и регуляцию биохимических реакций, в значительной степени изменяет интенсивность метаболизма в целом и соотношение между отдельными метаболическими путями. Следовательно, влияние температуры будет отражаться и на величине метаболического потока, который определяет общие энергетические затраты организма. В результате этого воздействие низких температур в значительной степени может изменять качественные и количественные показатели метаболических и продукционных процессов у морских животных, обитающих в высокоширотных районах Арктики, где условия существования видов изменяются в результате значительных пространственно-временных колебаний абиотических и биотических факторов.
В целом эффекты, вызываемые изменениями температуры, обусловлены восприимчивостью биохимических процессов, в том числе функций ферментов, и тем самым направляются адаптационными возможностями организма. Выявлению основных принципов организации таких адаптаций придается большое значение, поскольку их главные “консервативные” цели состоят в приспособлении интенсивности энергетического обмена, сохранении надлежащей структуры макромолекул, и, что еще очень важно, в регуляции клеточного метаболизма [1]. И хотя для каждого функционального компонента характерны свои специфические реакции на температуру, разным биохимическим системам свойственны некоторые общие особенности температурных эффектов и способов адаптации.
Живой организм – это открытая биологическая система, которая никогда не находится в равновесии и функционирует за счет свободной энергии (полезной части внутренней энергии), при этом постоянно совершая работу против навязываемого внешними условиями равновесия [2]. Как известно, общая энергия, получаемая организмом, равна:
т.е. внутренняя энергия ∆H равна сумме свободной ∆G и связанной энергии T∆S, и где ∆G – это свободная энергия активации (полезная часть внутренней энергии, являющаяся функцией состояния); ∆H – энтальпия активации (внутренняя или суммарная энергия); ∆S – изменение энтропии, T∆S – связанная энергия, которая не может быть использована для полезной работы (обесцененная часть внутренней энергии), и которая способна переходить только в теплоту и рассеиваться. Свободная энергия активации ∆G для той или иной метаболической реакции не есть постоянная величина, она может изменяться в соответствии с температурными условиями обитания животных. Вклад ∆H и ∆S в ∆G различен для разных химических реакций и, что особенно важно, для одних и тех же реакций, катализируемых несколько различающимися ферментами.Особое значение для температурных эффектов имеют две энергетические характеристики – это энтальпия (∆H – тепловая функция, внутренняя или суммарная энергия) и энтропия (∆S). Энтальпия – определенное качество вещества, указывающее на уровень энергии, характеризующий среднюю скорость движения реакционноспособных молекул, т.е. – это доступная для преобразования в теплоту энергия при сопутствующих значениях давления и температуры. Именно с величиной ∆H связаны значительные изменения скорости реакции при относительно небольших сдвигах температуры. Такие изменения взаимоотношений между скоростью реакции, сдвигом температуры и энтальпией активации можно считать одним из важнейших способов биохимической адаптации к температуре [1].
Все это становится весьма актуальным при исследовании процессов метаболизма и роста у эктотермных животных, обитающих в высокоширотных районах Арктики в относительно узком диапазоне низких температур и имеющих низкую скорость энергетического обмена и ограниченный аэробный метаболический объем. При вопросе о том, как холодноводные эктотермные организмы могут поддерживать максимальные аэробные скорости активного обмена, возникают важные соображения: во-первых, не могут ли рассматриваться низкие температуры, как фактор “давления” на их энергетику, и второй момент касается взаимоотношения биоэнергетической специфики с процессами роста и воспроизводства у рыб при низких температурах. Изучение особенностей метаболизма и роста у полярных морских видов рыб на уровне процессов обмена энергией и составило предмет настоящего сообщения.
Энергетический обмен. Эффективность роста и энтропия
В течение своей жизни организму необходимо столько потребить энергетических ресурсов, сколько он “сбрасывает” в окружающую себя среду, в основном в виде тепловых отходов, вырабатываемых его жизнеобеспечивающими системами. При этом следует отметить, что в качественном отношении не все типы энергии эквивалентны, поскольку они различны по происхождению [3]. Обесцененная энергия, отдаваемая живым организмом окружающей среде, находится в состоянии значительной неупорядоченности и обладает высокой энтропией. Поэтому такую энергию невозможно использовать для какой-либо полезной работы. Анализ связи между теплотой и энтропией выводит нас на следующий механизм: организм, потребляя высококачественную энергию, использует ее для поддержания и развития своих упорядоченных структур, производя при этом уже низкокачественную энергию в виде тепловых и других отходов. Под потоком теплоты подразумевается не само вещество, а определенное качество этого вещества. Поток теплоты и поток энтропии очень тесно связаны и, как говорят физики, “энтропия путешествует на волне теплоты”. Когда энергия течет через систему, она уносит прочь энтропию, поддерживая, таким образом, или даже порождая, порядок на своем пути.
Функция под названием “энтропия” позволяет выявить различия между свободной энергией активации (полезной части внутренней энергии) и связанной энергией (диссипированной, обесцененной части внутренней энергии), которая теряется необратимо [4]. Ключевое слово в этом объяснении – “необратимость” (центральное свойство всякого развития), поскольку именно необратимые процессы обусловливают производство энтропии внутри организма. И. Пригожиным было показано, что разные формы энергии продуцируют качественно различающиеся изменения энтропии в открытой биологической системе, и энтропия связана именно с внутренней энергией (это важный момент, так как в обратимом процессе параллель между энтропией и энергией заканчивается). Если обозначить dS – полное приращение энтропии за интервал времени dt, deS – относящееся к обмену энергией с внешней средой, а diS – к производству энтропии вследствие необратимых процессов внутри системы, то уравнение Пригожина имеет вид:
при этом увеличение внутренней энтропии организма не изменяет знак при обмене тепла с окружающей средой, величина приращения энтропии diS может быть только положительной. А составляющая deS может быть как положительной, так и отрицательной. Таким образом, для открытой биологической системы любые изменения энтропии не равнозначны, в этом и заключается смысл уравнения dS/dt = deS/dt + diS/dt, [5]. Вероятностное изменение внутренней энтропии diS отличается от изменения deS, обусловливаемого колебаниями таких факторов, как температура, конвекция, излучение. Например, низкие температуры ведут к снижению deS, а значит и к увеличению свободной энергии.В живых организмах используется такой механизм энергетического сопряжения, что системы, переносящие энергию, заряжаются в одних условиях, а разряжаются в других, и в первом случае система удаляется от того равновесия, к которому она будет возвращаться во втором [1]. Такой анализ показывает нам, что все анаболические или связанные с выполнением клеткой работы реакции сопряжены с катаболическими реакциями, приводящими к синтезу АТФ (аденозинтрифосфата). Химическая основа сопряжения таких реакций как раз и состоит в переносе энергии между этими блоками в форме АТФ. Сопряжение осуществляется при участии структурных элементов клетки, при этом, несомненно, биосинтетический процесс различных специфических клеточных компонентов в организме сопровождается генерированием энтропии, интенсивность которой определяется пространственной структурой макромолекул синтезируемого продукта. И чем сложнее структура этого продукта, тем в большей степени уменьшается энтропия при синтезе (diS), но при этом наблюдается больший прирост диссипативной энтропии (deS) [4].
При низких температурах обитания и сезонной ограниченности кормовых ресурсов пойкилотермные организмы высокоширотных районов Арктики растут значительно медленнее по сравнению с умеренными и тропическими видами. Проводимые нами исследования с тремя видами камбал – Liopsetta glacialis полярной камбалой, Pleuronectes platessa морской камбалой и Scophthalmus maeoticus черноморским калканом, показали, что у полярной камбалы, которая проводит значительную часть года при отрицательных придонных температурах [6], удельная скорость роста (Cw) самая низкая (рис. 1).
Рис. 1.
Изменение скорости роста Cw с массой тела W у рыб семейства Pleuronectidae, обитающих в разных областях Мирового океана.
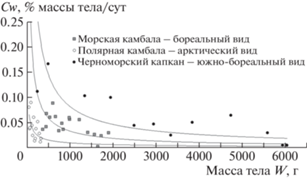
Несомненно, значительное влияние на изменение потока энтропии оказывают такие важнейшие функциональные процессы клеточного метаболизма, как биосинтез, рост, интеграция. Рост организмов в основном поддерживается процессами, протекающими с потреблением большого количества энергии – это белковый рост и сохранение баланса между синтезом и распадом белков [7]. Поэтому усиление энергообмена, связанного с метаболическим выделением теплоты в результате переваривания и абсорбции пищи (так называемое специфическое динамическое действие, СДД), вызывает увеличение скорости синтеза и оборота белка и тем самым отражает энергетические расходы, связанные с ростом, что описывается уравнением: Q = rP, где P – энергетические затраты на рост организма, r – коэффициент пропорциональности, описывающий энергетическое содержание единицы роста. Исследовать эту зависимость можно на основании свойств и взаимодействий двух процессов – энергетического обмена и роста, и предположить, что у рыб усиление интенсивности синтеза белка объясняет большую часть расходов энергии на СДД пищи. Результаты наших экспериментальных исследований показывают, что абсолютное увеличение энергетических затрат (максимальные значения пищевого обмена) на специфическое динамическое действие пищи для рыб, обитающих при низких температурах, ниже, чем для рыб умеренных и тропических широт. Однако это приводит к большей продолжительности СДД и тем самым снижает удельную скорость потребления пищи. Анализ рассчитанных нами годовых рационов рыб в природных условиях подтверждает этот вывод: потребление пищи у рыб высоких широт ниже, чем у рыб, обитающих в средних и низких широтах (табл. 1). Основываясь на полученных данных, следует отметить, что такая минимизация потребления энергетических ресурсов в течение года и может являться одним из механизмов адаптации к жизни у пойкилотермных организмов, обитающих в высокоширотных районах Арктики.
Таблица 1.
Годовые рационы арктических и бореальных видов рыб в природных условиях
| Возраст, лет | Годовой рацион, % массы тела | ||||
|---|---|---|---|---|---|
| Атлантическая треска (Северное море, t = 8°) | Атлантическая треска (Баренцево море, t = 2°) | Сайка (Арктика) | Ледяная треска (Арктика) | Морская камбала (Баренцево море) | |
| 1–2 | 758.9 | 524.8 | 400.9 | – | 409.6 |
| 2–3 | 540.8 | 402.3 | 330.7 | – | 334.8 |
| 3–4 | 464.9 | 364.0 | 337.5 | – | 280.1 |
| 4–5 | 388.6 | 322.1 | 313.1 | – | 279.5 |
| 5–6 | 355.1 | 292.6 | 276.4 | 269.8 | 181.7 |
| 6–7 | 337.7 | 270.6 | 276.0 | 238.1 | 178.8 |
| 7–8 | 328.4 | 253.3 | – | – | 167.8 |
| 8–9 | 319.5 | 239.3 | – | 219.1 | 163.6 |
| 9–10 | 310.3 | 227.7 | – | 207.9 | 159.6 |
Рассматривая биохимические адаптации в их взаимосвязи, мы отмечаем, как два раздельных, но энергетически сопряженных этапа метаболического процесса могут привести к перестройкам других особенностей организма в целом. Например, нами ранее показаны низкие скорости энергетического обмена у арктических видов рыб [8]. Этот факт позволяет нам полагать, что адаптивной стратегией в экстремальных высокоширотных условиях для пойкилотермных организмов может быть экономия энергетических затрат на поддерживающие жизнедеятельность функции, обеспечивающая освобождение части энергии для роста. При низких температурах у рыб такая экономия энергии может быть максимальной. Поскольку усвоенная организмом энергия пищи, необходимая для роста, может быть получена только двумя путями: либо за счет подавления других энергоемких функций, включенных в базальные энергетические затраты на поддержание жизнедеятельности, либо путем увеличения эффективности преобразования пищи в энергию ковалентных связей в молекулах белков, жиров и углеводов в процессе роста организма.
Биоэнергетические исследования количественных закономерностей соотношений соматического, генеративного и энергетического обмена мы также проводили и на некоторых представителях семейства тресковых – это Boreogadus saida сайка, Gadus morhua атлантическая треска и Gadus macrocephalus тихоокеанская треска. В этих работах на основе собственных экспериментальных измерений скорости роста при различных температурах и в режиме кормления до насыщения (при максимальных рационах) показано, что для всех трех видов тресковых удельная скорость роста с повышением температуры начинает быстро расти (рис. 2). С приближением к температурному оптимуму значения скорости роста достигают максимума и выходят на низкое плато. После чего кривая зависимости резко снижается под влиянием высоких температур, особенно у арктического вида – сайки.
Рис. 2.
Изменение скорости роста Cw с температурой у рыб семейства Gadidae, обитающих в разных областях Мирового океана. Все значения рассчитаны по нашим экспериментальным данным при кормлении в режиме ed libitum, для трески из Берингова моря определяли по [9].

Арктические виды рыб, например сайка, обитают большую часть года в воде, покрытой льдом, с отрицательной температурой. Поэтому температурный оптимум роста, как и следовало ожидать, сдвигается в направлении от арктического к бореальному виду, т.е. сдвиг происходит в сторону более высоких температур (рис. 2). А температуры, превышающие оптимальные для роста, приводят, как правило, к снижению эффективности конвертирования пищи [10], что может оказывать существенное влияние на способность эффективно использовать энергетические ресурсы на процессы роста и размножения. Следует отметить, что при низких температурах (около 0°С) скорость весового роста Cw у арктического вида – сайки, выше, чем у бореальных холодноводных рыб. Что указывает на более высокую степень эффективности прироста соматических и генеративных тканей у арктических видов.
Таким образом, анализ механизмов, лежащих в основе процессов обмена энергией у рыб, обитающих в высокоширотных районах Арктики, показывает, что потребление энергии в расчете на единицу роста становится тем экономнее, чем медленнее они растут (низкая удельная скорость роста). То есть процесс роста у рыб при более низких температурах становится все более эффективным, и они могут расти с минимальными затратами энергии. А поскольку при низких температурах все внутренние процессы замедляются и отходы накапливаются медленнее, то организм высвобождает меньше лишнего тепла, генерируемого его метаболическими процессами и ежедневной деятельностью. В этом и заключается эффективная стратегия использования низкоэнтропийной энергии и “сбрасывания” высокоэнтропийной теплоты при низких температурах у пойкилотермных животных. Организм и окружающая его среда, таким образом, участвуют в термодинамическом действии – организм выпускает излишек теплоты, что приводит к снижению энтропии, тогда как среда принимает эту теплоту и повышает свою энтропию. То есть рост энтропии среды компенсирует снижение энтропии, благодаря внутренним процессам биосинтеза при низких температурах, – и тем самым обеспечивает рост суммарной энтропии.
И в заключение хотелось бы отметить, что все живое находится в глубокой связи не только между собой, но также со звездами, планетами и другими упорядоченными структурами: жизнь – это еще одно средство, при помощи которого Вселенная высвобождает энтропийный потенциал, запертый в веществе [11].
Список литературы
Hochachka P.W., Somero G.N. Biochemical Adaptation: Mechanism and Process in Physiological Evolution. Oxford; N.Y.: Oxford Univ. Press, 2002. 466 p.
Бауэр Э.С. Теоретическая биология. // М.-Л.: Изд-во ВИЭМ, 1935. 206 с.
Иванов К.П. Новые биологические проблемы в энергетике живых систем // Успехи современной биологии. 2016. Т. 136. № 6. С. 586–592.
Пригожин И. Введение в термодинамику необратимых процессов. М.: Иностранная литература, 1960. 150 с.
Пригожин И., Стенгерс И. Порядок из хаоса. Новый диалог человека с природой. М.: Прогресс, 1986. 432 с.
Андрияшев А.П. Рыбы северных морей СССР. М.; Л.: Изд-во АН СССР, 1954. 556 с.
Fraser K.P.P., Rogers A.D. Protein metabolism in marine animals: the underlying mechanism of growth // Adv. Mar. Biol. 2007. V. 52. P. 267–362.
Карамушко Л.И., Шатуновский М.И. Адаптивная значимость различных форм метаболических процессов у морских видов рыб высоких широт // Успехи современной биологии. 2018. Т. 138. № 1. С. 12–17.
Laurel B.J., Spencer M., Iseri P., et al. Temperature-dependent growth and behavior of juvenile Arctic cod (Boreogadus saida) and co-occurring North Pacific gadids // Polar Biology. 2016. V. 39. iss. 6. P. 1127–1135.
Jobling M. Fish Bioenergetics. London: Chapman & Hall., 1994. 309 p.
Грин Б. До конца времен: Сознание, материя и поиск смысла в меняющейся Вселенной / Брайан Грин; Пер. с англ. – М.: Альпина нон-фикшн, 2021. 548 с.
Дополнительные материалы отсутствуют.
Инструменты
Доклады Российской академии наук. Науки о жизни