

Доклады Российской академии наук. Науки о жизни, 2023, T. 508, № 1, стр. 58-62
ПЕРВАЯ НАХОДКА Eothenomys (ARVICOLINAE, CRICETIDAE, RODENTIA) В ПЛЕЙСТОЦЕНЕ ВЬЕТНАМА
Академик РАН А. В. Лопатин 1, *
1 Палеонтологический институт им. А.А. Борисяка Российской академии наук
Москва, Россия
* E-mail: alopat@paleo.ru
Поступила в редакцию 27.09.2022
После доработки 10.10.2022
Принята к публикации 10.10.2022
- EDN: MPLHYT
- DOI: 10.31857/S2686738922600704
Аннотация
Из среднеплейстоценового пещерного местонахождения Там-Хай в северном Вьетнаме (провинция Лангшон) по изолированным зубам определена южноазиатская полевка Eothenomys eleusis (Thomas, 1911). Эта первая находка Arvicolinae в плейстоцене Вьетнама и первая ископаемая находка представителей рода Eothenomys за пределами Китая.
Единственный представитель подсемейства Arvicolinae во Вьетнаме – южноазиатская полевка, обитающая на самом севере страны, в горных лесах провинции Лаокай западнее Красной реки (Хонгха). Эта изолированная популяция Eothenomys обычно относится [1, 2] к подвиду E. melanogaster confinii Hinton, 1923 (= E. miletus confinii [3, 4]). Также вьетнамские полевки рассматривались в составе вида E. mucronatus (Allen, 1912) [4], который синонимизируется с E. melanogaster (Milne-Edwards, 1871) [5] или E. miletus (Thomas, 1914) [6]. В настоящее время по морфологическим и молекулярно-генетическим данным типичная форма E. miletus confinii считается [5, 7] идентичной качинской полевке E. cachinus (Thomas, 1921) (= E. eleusis cachinus [8]), тогда как вьетнамская популяция Eothenomys рассматривается в составе E. melanogaster [5], E. miletus [6] или E. eleusis (Thomas, 1911) [8]. Для вьетнамской популяции характерны сравнительно крупные размеры черепа и стабильное наличие четырех лингвальных выходящих углов (lingual salient angles) на последнем верхнем коренном зубе M3 [5]; при этом для E. melanogaster типично наличие трех лингвальных выходящих углов, а для E. eleusis (включая E. eleusis miletus) – четырех [5, 8–10].
Ископаемые находки Eothenomys melanogaster sensu lato известны в Китае с нижнего плейстоцена [11–13]. В настоящей статье описаны первые ископаемые находки Arvicolinae во Вьетнаме – изолированные зубы E. eleusis из среднего плейстоцена пещеры Там-Хай, расположенной в уезде Биньзя провинции Лангшон. Они собраны экспедиционным отрядом Палеонтологического института им. А.А. Борисяка РАН (ПИН) в марте 2021 г. в ходе работ Совместного Российско-Вьетнамского Тропического научно-исследовательского и технологического центра (Тропический центр, Ханой). Пещера Там-Хай (Thẩm Hai) входит в пещерный комплекс Там-Кхюиен (Thẩm Khuyên), известный совместными находками ранних людей и гигантопитеков и датированный средним плейстоценом (Там-Хай: 300–200 тыс. лет, 250 тыс. лет; Там-Кхюиен: 475 ± 125 тыс. лет) [14, 15]. Материал происходит из обособленной северной галереи пещеры (21°55′52.5″ с.ш., 106°25′4″ в.д.) [см. 16].
Описанный материал хранится в коллекции ПИН, Москва. Терминология строения зубов по [17], систематика Arvicolinae – по [8]. Изображения получены с помощью сканирующего электронного микроскопа Tescan Vega 3 XMU в ПИН.
Отряд Rodentia Bowdich, 1821
Семейство Cricetidae Fischer, 1817
Подсемейство Arvicolinae Gray, 1821
Триба Clethrionomyini Hooper et Hart, 1962
Подтриба Eothenomyina Kryštufek et Shenbrot, 2022
Род Eothenomys Miller, 1896
Eothenomys eleusis (Thomas, 1911)
Microtus (Eothenomys) melanogaster eleusis: [18, с. 50].
Голотип – Британский музей, Лондон, BM № 11.9.8.111, взрослый самец; Китай, Северная Юньнань; современный.
Описание (рис. 1, 2). Зубы бескорневые, с обильным цементом во входящих углах. Эмаль толстая, равномерной толщины на передних и задних сторонах треугольных призм, несколько тоньше в глубине входящих углов.
Рис. 1.
Eothenomys eleusis (Thomas, 1911): а, б – экз. ПИН, № 5826/556, левый M3: а – с окклюзиальной стороны, б – с буккальной стороны; в, г – экз. ПИН, № 5826/555, левый M1: в – с окклюзиальной стороны, г – с лингвальной стороны; д – экз. ПИН, № 5826/557, левый M2 с окклюзиальной стороны; е – экз. ПИН, № 5826/558, левый M3 с окклюзиальной стороны; Вьетнам, провинция Лангшон, пещера Там-Хай; средний плейстоцен.

Рис. 2.
Eothenomys eleusis (Thomas, 1911), строение жевательной поверхности коренных зубов: а – экз. ПИН, № 5826/556, левый M3; б – экз. ПИН, № 5826/555, левый M1; в – экз. ПИН, № 5826/557, левый M2; г – экз. ПИН, № 5826/558, левый M3; Вьетнам, провинция Лангшон, пещера Там-Хай; средний плейстоцен. Обозначения: AC – передняя непарная петля, AL – передняя петля, BRA1–BRA4 – буккальные входящие углы, BSA1–BSA4 – буккальные выходящие углы, LRA1–LRA5 – лингвальные входящие углы, LSA1–LSA5 – лингвальные выходящие углы, PC – задняя непарная петля (пятка), PL – задняя петля, T1–T7 – треугольные петли.
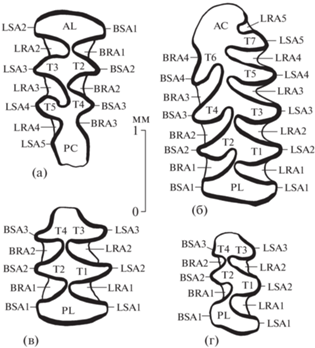
Последний верхний моляр M3 с широкой симметричной передней петлей (AL), четырьмя лингвальными и тремя буккальными выходящими углами и сильно удлиненной задней непарной петлей (пяткой), составляющей более 30% от общей длины зуба по жевательной поверхности. Треугольные петли попарно слиты (T2–T3, T4–T5), соответствующие входящие углы (BRA1–LRA2, BRA2–LRA3) противопоставлены. LRA3 намного глубже BRA2 и сильнее протягивается дистально. Наиболее дистальный лингвальный выходящий угол LSA5 едва выражен, но обособляющий его неглубокий лингвальный входящий угол LRA4 заполнен цементом, в отличие от противолежащей бесцементной буккальной складки, представляющей собой зачаточный буккальный входящий угол BRA3 (рис. 1а, 1б, 2а).
Передний нижний моляр M1 удлиненный (рис. 1в, 1г, 2б), с крупной передней непарной петлей (AC), тремя буккальными (BRA3, BRA2, BRA1) и пятью лингвальными (LRA5, LRA4, LRA3, LRA2, LRA1) входящими углами, заполненными цементом, и умеренно широкой задней петлей (PL) с противолежащими выходящими углами (BSA1–LSA1). Наиболее передний лингвальный входящий угол LRA5 сравнительно неглубоко вдается в переднюю непарную петлю (по глубине составляет приблизительно треть других входящих углов), обособляя переднюю из лингвальных треугольных петель – T7. Лингвальному входящему углу LRA4 противолежит неглубокая бесцементная борозда (зачаточный буккальный входящий угол BRA4), обозначающая обособление передней буккальной треугольной петли T6, противолежащей лингвальной треугольной петле T5. Более дистальные входящие углы чередующиеся (BRA3/LRA3, BRA2/LRA2, BRA1/LRA1), а треугольные петли попарно слитые (T4–T3, T2–T1), с противопоставленными выходящими углами (BSA3–LSA3, BSA2–LSA2).
M2 сравнительно короткий, с тремя одинаково широкими петлями (рис. 1д, 2в). Петли поперечно вытянутые, T4–T3 и T2–T1 попарно слитые, входящие и выходящие углы противопоставленные (BRA2–LRA2, BRA1–LRA1; BSA3–LSA3, BSA2–LSA2, BSA1–LSA1).
M3 мелкий, узкий, задняя петля сравнительно широкая (рис. 1е, 2г). Петли слабо расширены поперечно, передние входящие и все выходящие углы противопоставлены (BRA2–LRA2; BSA3–LSA3, BSA2–LSA2, BSA1–LSA1), тогда как BRA1 располагается заметно мезиальнее, чем LRA1, и направлен антеролингвально, а не поперечно, обозначая чередование T2 и T1.
Размеры (L – наибольшая длина, W – наибольшая ширина, в мм): M3 (экз. ПИН, № 5826/556): L – 2.05, W – 0.95, длина задней непарной петли – 0.65; M1 (экз. ПИН, № 5826/555): L – 2.65, W – 1.2; M2 (экз. ПИН, № 5826/557): L – 1.5, W – 1.0; M3 (экз. ПИН, № 5826/558): L – 1.3, W – 0.65.
Сравнение и замечания. Согласно современным данным [8], род Eothenomys включает три рецентных вида: E. melanogaster (Milne-Edwards, 1871) в южном Китае севернее р. Янцзы, E. colurnus (Thomas, 1911) в юго-восточном Китае южнее р. Янцзы и на о. Тайвань и E. eleusis (Tho-mas, 1911) в юго-западной части Китая и соседних областях Индии, Мьянмы, Вьетнама и Таиланда. E. eleusis рассматривается как политипический вид с пятью отчетливыми подвидами, помещенными в три подвидовые группы. Подвидовая группа eleusis включает E. eleusis eleusis (восточная часть видового ареала к югу от р. Янцзы в Китае, а также северный Вьетнам и северный Таиланд), E. eleusis miletus (Сычуань и Юньнань между р. Меконг и Красной рекой) и E. eleusis fidelis Hinton, 1923 (южная Сычуань между р. Ялунцзян и Красной рекой и соседние области Юньнани). Еще два подвида – E. eleusis cachinus (Thomas, 1921), обитающий на северо-западе Юньнани, севере Мьянмы и северо-востоке Индии, и E. eleusis shimianensis Liu in Liu et al., 2019, известный только из уезда Шимянь китайской провинции Сычуань, по отдельности образуют собственные подвидовые группы – cachinus и shimianensis соответственно [8].
Морфотип M3 с четырьмя лингвальными и тремя буккальными выходящими углами свойственен E. eleusis shimianensis (100%), E. eleusis eleusis (90%), E. eleusis miletus (90%), E. eleusis fidelis (90%), встречается у E. eleusis cachinus (35%) и крайне редок у E. melanogaster (2%) [7]. Морфотип M3 с четырьмя выходящими углами с буккальной и лингвальной сторон представлен у E. eleusis cachinus (50%), E. eleusis eleusis (10%) и E. eleusis fidelis (10%), но отсутствует у E. eleusis miletus, E. eleusis shimianensis, E. colurnus и подавляющего большинства (99%) экземпляров E. melanogaster [7].
Размеры, морфотип M3 с четырьмя лингвальными и тремя буккальными выходящими углами и строение других зубов позволяют относить полевку из среднего плейстоцена пещеры Там-Хай к E. eleusis.
Новые данные о современном таксономическом разнообразии Eothenomys [7, 8, 19] предполагают необходимость ревизии видовой идентификации ископаемых находок, отнесенных ранее к E. melanogaster (например, [11]).
Материал. Из местонахождения Там-Хай четыре изолированных зуба: M3 (экз. ПИН, № 5826/556), M1 (экз. ПИН, № 5826/555), M2 (экз. ПИН, № 5826/557) и M3 (экз. ПИН, № 5826/558).
Результаты филогеографического анализа [20] демонстрируют, что ареал Eothenomys melanogaster sensu lato (примерно соответствует объему подрода Eothenomys в понимании [7] и объему рода Eothenomys в понимании [8]) достиг современных пределов около 488 тыс. л.н. Дальнейшее развитие горного рельефа привело к возникновению адаптаций к средне- и высокогорным ландшафтам и холодному климату, последующей изоляции и генетической дифференциации отдельных высокогорных популяций во время потеплений (концепция “небесных островов”), что стало причиной формирования ряда таксонов видового и подвидового ранга [7, 20]. Современный дизъюнктивный ареал и таксономическое разнообразие Eothenomys сформировались под влиянием чередования теплых и холодных фаз плейстоцена. Предполагается быстрое обособление трех клад, представители которых обитают ныне в разных регионах Китая: юго-восточной клады (юго-восточное побережье Китая и о. Тайвань), центральной (горное обрамление Сычуаньской впадины и юго-восточный край Тибетского нагорья) и юго-западной (Юньнань-Гуйчжоуское нагорье и южная часть Сино-Тибетских гор) [20].
По современным представлениям [8], юго-восточная клада соответствует виду E. colurnus, центральная – E. melanogaster, а юго-западная – E. eleusis. Предполагаемый порядок ветвления клад в эволюционном сценарии [7]: E. colurnus (E. melanogaster (E. cachinus + E. shimianensis) (E. miletus (E. eleusis + E. fidelis))).
Описанные остатки представляет собой первую ископаемую находку Eothenomys за пределами Китая. Среднеплейстоценовая форма из Вьетнама имеет выраженные примитивные признаки коренных зубов (простое строение M3, чередующиеся T5/T6 на M1 и Т1/T2 на M3), в целом соответствующие наименее продвинутым морфотипам E. eleusis.
Присутствие E. eleusis в среднеплейстоценовом местонахождении Там-Хай предполагает возможность очень давнего обособления рецентной вьетнамской популяции Eothenomys, что позволяет ставить вопрос о ее подвидовой самостоятельности. Кроме того, локализация плейстоценовых находок в провинции Лангшон, северо-восточнее Красной реки, на значительном удалении от области обитания современной вьетнамской популяции в провинции Лаокай, свидетельствует о длительной и сложной истории E. eleusis в данном регионе.
Список литературы
Osgood W.H. Mammals of the Kelley-Roosevelts and Delacour Asiatic expeditions // Field Museum of Natural History. Zoological Series. 1932. V. 18. № 10. P. 193–339.
Кузнецов Г.В. Млекопитающие Вьетнама. М.: Т-во научных изданий КМК, 2006.
Wang Y.-X., Li C.-Y. Genus Eothenomys Miller, 1896. In: Luo Z.-X., Chen W., Gao W., eds. Fauna Sinica. Mammalia. V. 6. Rodentia. Part III: Cricetidae. Beijing: Science Press, 2000. P. 388–462.
Kaneko Y. Morphological variation and geographical and altitudinal distribution in Eothenomys melanogaster and E. mucronatus (Rodentia, Arvicolinae) in China, Taiwan, Burma, India, Thailand, and Vietnam // Mammal Study. 2002. V. 27. P. 31–63.
Wilson D.E., Reeder D.M., eds. Mammal species of the World: a taxonomic and geographic reference. 3 ed. Baltimore, Maryland: The Johns Hopkins University Press, 2005.
Pardiñas U.F.J., Myers P., León-Paniagua L. et al. Cricetidae (true hamsters, voles, lemmings and New World rats and mice). In: Wilson D.E., Lacher T.E., Mittermeier R.A., eds. Handbook of the mammals of the World. V. 7. Rodents II. Barcelona: Lynx Ed., 2017. P. 204–535.
Liu S.-Y., Chen S.-D., He K. et al. Molecular phylogeny and taxonomy of subgenus Eothenomys (Cricetidae: Arvicolinae: Eothenomys) with the description of four new species from Sichuan, China // Zoological Journal of the Linnean Society. 2019. V. 186. P. 569–598.
Kryštufek B., Shenbrot G.I. Voles and lemmings (Arvicolinae) of the Palaearctic Region. Maribor: University of Maribor Press, 2022.
Allen G.M. Mammals of China and Mongolia // The natural history of Central Asia. V. 11. Part 2. New York: American Museum of Natural History, 1940. P. 621–1350.
Corbet G.B., Hill J.E. Mammals of the Indomalayan Region: systematic review. Oxford: Oxford University Press, 1992.
Zheng S. Quaternary rodents of Sichuan-Guizhou Area, China. Beijing: Science Press, 1993.
Zheng S. Rodentia. In: Zheng S., ed. Jianshi hominid site. Beijing: Science Press, 2004. P. 121–181, 387–394.
Jin C.Z., Zhang Y.Q., Wei G.B. et al. Rodentia Bowdich, 1821. In: Jin C., Liu J., eds. Paleolithic site – the Renzidong Cave, Fanchang, Anhui Province. Beijing: Science Press, 2009. P. 166–220, 401–412.
Olsen J.W., Ciochon R.L. A review of evidence for postulated Middle Pleistocene occupations in Viet Nam // Journal of Human Evolution. 1990. V. 19. P. 761–788.
Ciochon R., Vu The Long, Larick R. et al. Dated co-occurrence of Homo erectus and Gigantopithecus from Tham Khuyen Cave, Vietnam // Proceedings of the National Academy of Sciences. 1996. V. 93. № 7. P. 3016–3020.
Лопатин А.В. Новый вид Typhlomys (Platacanthomyidae, Rodentia) из среднего плейстоцена северного Вьетнама // Доклады Российской академии наук. Науки о жизни. 2021. Т. 501. № 1. С. 505–510.
Тесаков А.С. Биостратиграфия среднего плиоцена – эоплейстоцена Восточной Европы (по мелким млекопитающим). Москва: Наука, 2004.
Thomas O. On mammals collected in the provinces of Sze-chuan and Yunnan, western China, by Mr. Malcolm Anderson, for the Duke of Bedford’s Exploration // Abstracts of the Proceedings of the Zoological Society of London. 1911. № 100. P. 48–50.
Tang M., Chen Z., Wang X. et al. A summary of phylogenetic systematics studies of Myodini in China (Rodentia: Cricetidae: Arvicolinae) // Acta Theriologica Sinica. 2021. V. 41. № 1. P. 71–81.
Lu X., Cheng J., Meng Y. et al. Disjunct distribution and distinct intraspecific diversification of Eothenomys melanogaster in South China // BMC Evolutionary Biology. 2018. V. 18. Art. 50. https://doi.org/10.1186/s12862-018-1168-3
Дополнительные материалы отсутствуют.
Инструменты
Доклады Российской академии наук. Науки о жизни