

Экология, 2019, № 1, стр. 30-39
Влияние эколого-географических факторов в широтно-долготном градиенте на систему “красной” крови автохтона Кавказа – гудаурской полевки (Chionomys gud)
З. Х. Боттаева a, *, Ф. А. Темботова a, М. М. Емкужева a, З. А. Берсекова a, А. Х. Чапаев a
a Институт экологии горных территорий им. А.К. Темботова РАН
360051 КБР, Нальчик, ул. Инессы Арманд, 37а, Россия
* E-mail: zulya_bot@mail.ru
Поступила в редакцию 11.07.2017
После доработки 15.06.2018
Принята к публикации 05.06.2018
Аннотация
Впервые на примере типично горного вида мелких млекопитающих – гудаурской полевки (Chionomys gud Satunin, 1909) – показатели кроветворной ткани и периферической крови исследованы не в высотном, а широтно-долготном градиенте. В условиях приморского климата западной части и континентального климата центральной части Кавказа на одной высоте (1800 м над ур. м.) у вида выявлена сходная кислородная емкость крови, но механизмы ее обеспечения различны. Полученные данные свидетельствуют в пользу того, что наиболее оптимальны для вида условия Западного Кавказа.
При изучении механизмов адаптации животных к условиям гор, в том числе и на Кавказе, в центре внимания исследователей на протяжении более века и до настоящего времени остается высотная поясность (вертикальная зональность). Однако горные ландшафты как в целом, так и отдельные их элементы несут глубокие следы влияния широтных зон, и спектральные отрезки одного и того же пояса могут различаться даже больше, чем соседние высотные пояса одного и того же макросклона. Горные хребты Кавказа располагаются в пределах пяти широтных зон, соответственно столько же и типов поясности, которые в зависимости от близости к океану и другим крупным источникам влаги, направления воздушных течений делятся на приморский и континентальный подтипы. Особенности поясного спектра, обусловленные региональными условиями (ориентация хребтов, местный климат, орография и др.), отражает вариант поясности [1, 2].
Механизмы приспособления к горным условиям у типично горных животных имеют существенные отличия в сравнении с таковыми горных популяций широко распространенных видов. Так, если у первой группы процесс адаптации происходит на тканевом уровне, то у второй – на органном [3–13].
Особую роль в реализации приспособительных реакций организма в изменяющихся условиях среды обитания выполняет система крови, поскольку характер обменных процессов в тканях существенно зависит от обеспеченности тканей необходимым количеством кислорода. Регуляция кислородного гомеостаза организма осуществляется при участии “красной” крови посредством изменения уровня эритропоэза и кислородно-транспортных свойств гемоглобина [14].
Исследования системы крови млекопитающих в условиях Кавказа ранее проводились с учетом лишь высотной поясности [15–19]. Изменчивость в неоднородных условиях с учетом секторности, т.е. горизонтальной, или широтной, неоднородности поясных спектров, практически не изучена. Отсутствуют подобные исследования и в других горных системах. В этой связи нами были начаты исследования системы крови и кроветворения млекопитающих в горных условиях с учетом горизонтальной зональности. Объектом для настоящей работы был выбран типично горный вид мелких млекопитающих – гудаурская полевка (Chionomys gud Satunin, 1909). Это автохтон Кавказа, распространение которого охватывает в основном альпийский и субальпийский пояса до 3000–3500 м над ур. м. [20]. Вид – стенотопный, строго приуроченный к каменистым местообитаниям, в силу чего его распространение имеет прерывистый характер, структура ареала мозаична и в ней выделяются группы популяций, в той или иной степени изолированные друг от друга. Оледенения Большого Кавказа в плейстоцене не смогли вытеснить вид на равнину, но частично понизили границы его распространения, что, по мнению Н.К. Верещагина [21], и обусловило его появление на высоте, не характерной для вида (600 м над ур. м.), – в Пятигорье на изолированном участке (гора Развалка).
Помимо распространения, ландшафтной и биотопической приуроченности вида, разными исследователями также изучены вопросы, касающиеся популяционной структуры, динамики численности, особенностей размножения и развития, морфологических особенностей, кариотипов и таксономического статуса [22–28]. Современные исследования Ch. gud, посвященные уточнению таксономического статуса, направлены на установление генетических связей между популяциями вида и его происхождение [29–31]. Наименее изучены вопросы, затрагивающие особенности физиологии вида.
Ранее нами была показана изменчивость иммунологических показателей (“белая” кровь) Ch. gud в условиях секторальной неоднородности ландшафтов [32]. Сведения о показателях кислородтранспортной функции крови (“красная” кровь) вида, имеющиеся в литературе, обрывочны и фрагментарны, так как они исследовались лишь в условиях Центрального Кавказа и в сравнительном аспекте не изучались [7, 33]. Отсутствуют данные и по кроветворной функции костного мозга вида.
Мы предположили, что в разных вариантах поясности Западного и Центрального Кавказа, обусловленных секторальной неоднородностью, между популяциями Ch. gud в системе “красной” крови будут обнаружены различия, связанные с действием различных природно-климатических факторов, и в первую очередь температуры и влажности. Эти предположения основаны на том, что температура и влажность являются ведущими факторами и тесно взаимосвязаны между собой. Они достаточно надежно оцениваются количественно и с их взаимодействием коррелирует большинство экологических явлений в природе. В горах условия обитания животных осложняются как из-за резких перепадов температуры и влажности, так и понижения атмосферного давления и связанного с ним низкого парциального давления кислорода.
На наш взгляд, изменчивость системы крови, выявленная у гудаурской полевки в разных вариантах поясности, позволит оценить влияние разных природно-климатических условий на механизмы обеспечения организма кислородом, а также охарактеризовать эти условия как благоприятные или неблагоприятные для вида. Выявление оптимальных условий существования может быть полезным при изучении таких важных вопросов, как происхождение и эволюция вида.
Цель настоящей работы – сравнительное изучение показателей кроветворной ткани костного мозга и периферической крови Ch. gud в разных вариантах поясности среднегорий Западного и Центрального Кавказа, обусловленных секторальной неоднородностью, а также оценка влияния климатических факторов на эти показатели.
МАТЕРИАЛ И МЕТОДЫ
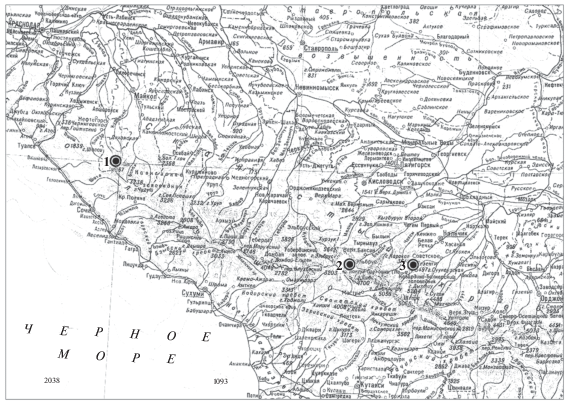
Исследования проводили на Западном Кавказе в условиях кубанского варианта поясности (Адыгея, окр. плато Лагонаки, 44°04′ с.ш., 40°00′ в.д.) и на Центральном Кавказе (Кабардино-Балкария) в условиях эльбрусского (окр. п. Эльбрус, 43°15′ с.ш., 42°38′ в.д.) и терского (окр. с. Безенги, 43°09′ с.ш., 43°12′ в.д.) вариантов в пределах субальпийского пояса на высоте 1800 м над ур. м. (рис. 1).
Рис. 1.
Места отлова Ch. gud на Западном и Центральном Кавказе: 1 – плато Лагонаки (кубанский вариант поясности, приморский подтип); 2 – окр. п. Эльбрус (эльбрусский вариант, континентальный подтип); 3 – окр. с. Безенги (терский вариант, континентальный подтип).
Согласно типизации поясных спектров, разработанной А.К. Темботовым [1, 2], кубанский вариант поясности выделяется в пределах приморского подтипа западно-северокавказского (степного) типа поясности. Благодаря влиянию воздушных масс Средиземно-Черноморского бассейна и Атлантического океана, климат здесь влажный и мягкий. Влияние влажных воздушных масс резко ослабевает к востоку. Эльбрусский и терский варианты относятся к континентальному подтипу восточно-северокавказского (полупустынного) типа поясности, который резко отличается от западно-северокавказского по степени ксерофитизации ландшафтов. Высокогорные осевые хребты с вечными снегами усиливают здесь континентальность климата. Ряд климатических характеристик районов исследования приведен в табл. 1. Использовали данные интернет-ресурса [34] и собственные данные, полученные с помощью метеостанции (Vantage Pro2 6152 Davis, США).
Таблица 1.
Некоторые климатические характеристики районов исследования в разных вариантах поясности среднегорий (1800 м над ур. м.) Западного и Центрального Кавказа
| Показатели | Западный Кавказ (приморский подтип поясности) | Центральный Кавказ (континентальный подтип поясности) | |
|---|---|---|---|
| окр. пл. Лагонаки (кубанский вариант) | окр. п. Эльбрус (эльбрусский вариант) | окр. с. Безенги (терский вариант) | |
| Среднегодовая температура, °С | 5.7 | 3.8 | 5.2 |
| Среднегодовое количество осадков, мм | 1485 | 920 | 930 |
| Атмосферное давление, мм рт. ст. | 650 | 600 | 620 |
| Парциальное давление кислорода, мм рт. ст. | 136.1 | 125.6 | 129.8 |
Исследовали эритроидный ряд клеток костного мозга и периферической крови 102 особей гудаурской полевки: в условиях кубанского варианта поясности – 14 самцов и 21 самка, эльбрусского – 12 самцов и 16 самок, терского – 15 самцов и 24 самки. В работе использовали только половозрелых животных, которых определяли по состоянию генеративных органов [35].
Отлов животных проводили в летний период (июль–август) с 2013 по 2015 г. Площадь участков исследований составила: в кубанском варианте поясности – 4.6 га, эльбрусском – 3.8 га, терском – 5.0 га. В качестве орудий лова применяли живоловушки. Отловленных животных доставляли в лабораторию, где сразу же после небольшого наркоза (эфир) получали кровь при вскрытии кровеносных сосудов и костный мозг из диафиза бедренной кости для дальнейших исследований.
Препараты крови и костного мозга окрашивали комбинированным методом по Май–Грюнвальду и Романовскому–Гимза. Эритропоэтическую функцию костного мозга изучали по морфологии клеток эритроидного ряда с дифференциацией на эритробласты, базофильные, полихроматофильные и оксифильные нормобласты. Количество гемоглобина (г/л) определяли унифицированным гемиглобинцианидным методом на гемоглобинометре фотометрическом МиниГЕМ-540 (Россия), гематокритное число (об. %) – микрометодом на гематокритной центрифуге СМ-70 (Латвия). Подсчет количества эритроцитов (млн в 1 мкл) проводили в камере Горяева. Диаметр эритроцитов (мкм) измеряли прямым микрометрическим методом с помощью винтового окуляр-микрометра МАВ 1-16х на сухих окрашенных препаратах крови. Вычисляли следующие эритроцитарные индексы: среднее содержание гемоглобина в эритроците (пг), средний объем эритроцита (мкм3), среднюю концентрацию гемоглобина в эритроците (%) [36, 37].
Статистическую обработку данных проводили с использованием пакета прикладных программ Statistica-10. Различия гематологических показателей между выборками животных из разных вариантов поясности выявляли с помощью дисперсионного и дискриминантного анализов, половых различий показателей – t-критерия Стъюдента. Различия считали значимыми при p < 0.05.
РЕЗУЛЬТАТЫ
С помощью дискриминантного анализа установлено, что выборки гудаурской полевки из разных вариантов поясности Западного и Центрального Кавказа по изученным показателям системы “красной” крови существенно различаются (табл. 2). Наиболее сходными являются популяции терского и эльбрусского вариантов. Согласно классификационной матрице, 77.94% животных соответствуют априорной классификации. Наибольшее соответствие отмечено в кубанской выборке (96.00%), несколько меньше – в терской (72.72%) и минимальное – в эльбрусской (61.96%) (рис. 2). Пол животных не влияет ни на один из изученных показателей системы крови, однако данные по самцам и самкам приведены отдельно (табл. 3).
Таблица 2.
Оценка различий между выборками Ch. gud из разных вариантов поясности среднегорий Западного и Центрального Кавказа
| Вариант поясности | 1 | 2 | 3 |
|---|---|---|---|
| Кубанский (1) | 45.61 | 25.21 | |
| Эльбрусский (2) | 13.61 | 8.77 | |
| Терский (3) | 7.12 | 2.73 |
Рис. 2.
Диаграмма рассеяния. Расположение выборок Ch. gud, полученных на основании классификации по показателям системы крови в координатах дискриминантного анализа (эллипсы – 95%-ный доверительный интервал): ⚪ – кубанский вариант, ◻ – эльбрусский вариант, ▲ – терский вариант.
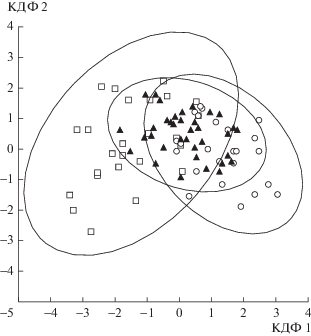
Таблица 3.
Показатели эритроидного ряда костного мозга и периферической крови Ch. gud в разных вариантах поясности среднегорий Западного и Центрального Кавказа (X ± m)
| Показатели | Пол | Западный Кавказ (приморский подтип) |
Центральный Кавказ (континентальный подтип) |
Уровень значимости | F | p | |||
|---|---|---|---|---|---|---|---|---|---|
| Кубанский вариант (1) |
Эльбрусский вариант (2) | Терский вариант (3) |
1–2 | 1–3 | 2–3 | ||||
| Общее количество эритроидных клеток, % | ♂♂ ♀♀ |
19.26 ± 1.031 18.35 ± 0.750 |
23.92 ± 1.738 25.23 ± 0.745 |
24.19 ± 0.789 25.70 ± 0.893 |
0.011 0.000 |
0.005 0.000 |
0.873 0.695 |
5.50 23.37 |
0.009 0.000 |
| Эритробласт, % | ♂♂ ♀♀ |
2.26 ± 0.157 2.61 ± 0.147 |
4.08 ± 0.364 3.75 ± 0.296 |
2.51 ± 0.108 2.82 ± 0.128 |
0.000 0.000 |
0.399 0.427 |
0.000 0.001 |
19.23 9.40 |
0.000 0.000 |
| Базофильный нормобласт, % | ♂♂ ♀♀ |
8.14 ± 0.627 7.11 ± 0.523 |
7.83 ± 0.333 7.54 ± 0.348 |
7.95 ± 0.432 7.99 ± 0.212 |
0.658 0.423 |
0.774 0.077 |
0.856 0.373 |
0.10 1.64 |
0.903 0.204 |
| Полихроматофильный нормобласт, % | ♂♂ ♀♀ |
35.51 ± 0.821 35.30 ± 0.809 |
37.40 ± 1.083 36.25 ± 0.732 |
38.77 ± 1.019 38.09 ± 0.647 |
0.201 0.395 |
0.023 0.007 |
0.335 0.076 |
2.88 4.14 |
0.071 0.021 |
| Оксифильный нормобласт, % | ♂♂ ♀♀ |
52.75 ± 1.079 52.97 ± 1.186 |
49.04 ± 1.631 51.34 ± 0.826 |
49.68 ± 1.191 49.79 ± 0.773 |
0.061 0.246 |
0.095 0.016 |
0.732 0.239 |
2.26 3.11 |
0.121 0.053 |
| Гемоглобин, г/л | ♂♂ ♀♀ |
142.09 ± 2.918 139.55 ± 1.870 |
140.30 ± 1.660 139.75 ± 1.955 |
136.00 ± 1.795 136.69 ± 1.482 |
0.576 0.940 |
0.072 0.245 |
0.207 0.274 |
1.81 0.88 |
0.183 0.422 |
| Количество эритроцитов, млн | ♂♂ ♀♀ |
8.70 ± 0.313 8.76 ± 0.163 |
10.21 ± 0.236 9.92 ± 0.288 |
9.05 ± 0.171 9.26 ± 0.184 |
0.000 0.000 |
0.337 0.071 |
0.006 0.037 |
8.55 7.79 |
0.001 0.001 |
| Гематокрит, об. % | ♂♂ ♀♀ |
45.08 ± 0.839 44.10 ± 0.740 |
45.20 ± 0.629 44.83 ± 0.534 |
43.36 ± 0.364 43.67 ± 0.464 |
0.901 0.443 |
0.069 0.615 |
0.057 0.241 |
2.42 0.71 |
0.106 0.498 |
| Диаметр эритроцитов, мкм | ♂♂ ♀♀ |
5.84 ± 0.032 5.82 ± 0.041 |
5.50 ± 0.032 5.49 ± 0.037 |
5.64 ± 0.034 5.65 ± 0.025 |
0.000 0.000 |
0.000 0.000 |
0.007 0.002 |
25.34 21.69 |
0.000 0.000 |
| Среднее содержание гемоглобина в эритроците, пг |
♂♂ ♀♀ |
16.63 ± 0.408 16.09 ± 0.222 |
13.78 ± 0.192 14.01 ± 0.319 |
14.79 ± 0.214 14.68 ± 0.201 |
0.000 0.000 |
0.000 0.000 |
0.031 0.076 |
23.82 19.71 |
0.000 0.000 |
| Средний объем эритроцитов, мкм3 | ♂♂ ♀♀ |
51.95 ± 1.092 50.46 ± 0.615 |
44.42 ± 0.902 45.04 ± 1.349 |
48.05 ± 0.860 47.57 ± 0.649 |
0.000 0.000 |
0.007 0.010 |
0.015 0.048 |
14.93 10.68 |
0.000 0.000 |
| Средняя концентрация гемоглобина в эритроците, % |
♂♂ ♀♀ |
31.83 ± 0.337 32.00 ± 0.399 |
31.03 ± 0.305 31.21 ± 0.506 |
31.23 ± 0.409 31.02 ± 0.146 |
0.111 0.153 |
0.227 0.057 |
0.724 0.746 |
1.46 2.17 |
0.251 0.126 |
Эритропоэз. Эритроидный ряд клеток кроветворной ткани костного мозга Ch. gud представлен эритробластами и нормобластами всех последовательных стадий созревания – базофильными, полихроматофильными и оксифильными. Общее количество эритроидных клеток по отношению ко всем ядерным элементам в костном мозге у животных в условиях кубанского варианта поясности на Западном Кавказе существенно ниже, чем в обоих вариантах Центрального Кавказа (эльбрусском и терском). Следовательно, эритропоэтическая активность костного мозга полевок на Центральном Кавказе значительно выше. У центральнокавказских особей в условиях эльбрусского варианта на фоне высокого содержания эритроидных клеток в костном мозге отмечено значительное количество эритробластов – наиболее молодых клеток эритроидного ряда. Содержание базофильных нормобластов у животных всех трех популяций на одном уровне. У полевок терского варианта больше полихроматофильных нормобластов, несколько меньше данный показатель у животных в эльбрусском варианте и значительно меньше – в кубанском. Количество наиболее зрелых форм – оксифильных нормобластов, наоборот, выше у полевок на Западном Кавказе (см. табл. 3).
Периферическая кровь. Содержание гемоглобина в крови Ch. gud во всех трех вариантах в целом невысокое и существенно не различается. Также практически не различается и гематокритная величина. При этом количество и размеры эритроцитов, а также насыщенность их гемоглобином сильно варьируют (см. табл. 3).
На Центральном Кавказе в условиях эльбрусского варианта у полевок обнаружено большое количество мелких эритроцитов (как по диаметру, так и по объему) с меньшим содержанием в них гемоглобина, в то время как на Западном Кавказе, наоборот, у животных более крупные клетки в меньшем количестве, но в большей степени насыщенные гемоглобином. Популяция терского варианта поясности по этим параметрам занимает промежуточное положение. По среднему диаметру и среднему объему эритроцитов, а также среднему содержанию гемоглобина в эритроците существенная разница обнаружена как при сравнении западнокавказской популяции с центральнокавказскими, так и центральнокавказских между собой.
Выявленные различия подтверждаются и при попарном сравнении выборок. Западнокавказскую популяцию существенно отличает от обеих центральнокавказских общее количество эритроидных клеток в костном мозге, а от эльбрусской популяции еще и количество эритробластов (табл. 4). По показателям периферической крови самцов различает средний диаметр эритроцитов, самок – среднее содержание гемоглобина в эритроцитах. Центральнокавказские популяции различимы между собой по количеству эритробластов. Также в качестве различающего признака выступает среднее содержание гемоглобина в эритроците у самцов и количество эритроцитов у самок (табл. 5).
Таблица 4.
Дискриминирующие признаки эритроидного ряда костного мозга Ch. gud в разных вариантах поясности среднегорий Западного и Центрального Кавказа
| Показатели | F | p | Показатели | F | p |
|---|---|---|---|---|---|
| самцы | самки | ||||
| Кубанский вариант–эльбрусский вариант | |||||
| F (1.19) = 22.575, p < 0.0001 | F (1.30) = 28.854, p < 0.0000 | ||||
| Эритробласты | 22.57 | 0.000 | Общее количество эритроидных клеток | 31.98 | 0.000 |
| Эритробласты | 7.15 | 0.012 | |||
| Процент дискриминации | 85.71 | Процент дискриминации | 90.62 | ||
| Кубанский вариант–терский вариант | |||||
| F (1.22) = 14.868, p < 0.0009 | F (1.37) = 43.577, p < 0.0000 | ||||
| Общее количество эритроидных клеток |
14.87 | 0.001 | Общее количество эритроидных клеток |
43.58 | 0.000 |
| Процент дискриминации | 79.16 | Процент дискриминации | 87.50 | ||
| Терский вариант–эльбрусский вариант | |||||
| F (1.20) = 20.205, p < 0.0002 | F (1.34) = 11.326, p < 0.0019 | ||||
| Эритробласты | 21.20 | 0.000 | Эритробласты | 11.33 | 0.002 |
| Процент дискриминации | 82.61 | Процент дискриминации | 72.50 | ||
Таблица 5.
Дискриминирующие признаки периферической крови Ch. gud в разных вариантах поясности среднегорий Западного и Центрального Кавказа
| Показатели | F | p | Показатели | F | p |
|---|---|---|---|---|---|
| самцы | самки | ||||
| Кубанский вариант–эльбрусский вариант | |||||
| F (1.18) = 46.820, p < 0.0000 | F (1.27) = 26.359, p <0.0000 | ||||
| Диаметр эритроцитов | 46.82 | 0.000 | Среднее содержание гемоглобина в эритроцитах | 26.36 | 0.000 |
| Процент дискриминации | 88.46 | Процент дискриминации | 90.62 | ||
| Кубанский вариант–терский вариант | |||||
| F (1.18) = 20.565, p < 0.0003 | F (1.32) = 21.263, p < 0.0001 | ||||
| Диаметр эритроцитов | 20.56 | 0.000 | Среднее содержание гемоглобина в эритроцитах | 21.26 | 0.000 |
| Процент дискриминации | 79.31 | Процент дискриминации | 77.78 | ||
| Терский вариант–эльбрусский вариант | |||||
| F (1.16) = 10.236, p < 0.0056 | F (1.25) = 4.308, p < 0.0484 | ||||
| Среднее содержание гемоглобина в эритроцитах | 10.24 | 0.006 | Количество эритроцитов | 4.31 | 0.048 |
| Процент дискриминации | 83.33 | Процент дискриминации | 66.67 | ||
ОБСУЖДЕНИЕ РЕЗУЛЬТАТОВ
Изменчивость эритропоэтической активности костного мозга Ch. gud сопровождается колебаниями показателей периферической крови (см. табл. 3). Сопряженная изменчивость костного мозга и периферической крови описана ранее на примере рыжей полевки [38]. Согласно этим данным, число эритроцитов имеет прямую связь с содержанием клеток эритроидного ростка костного мозга и обратную – с содержанием базофильных и полихроматофильных нормобластов.
Известно, что в процессах регуляции эритропоэза как в норме, так и при различных стрессовых ситуациях происходит разрушение (эритродиерез) определенной части эритроцитов, продукты распада которых затем стимулируют эритропоэз. Разрушение эритроцитов является неотъемлемой частью эритропоэза. Оба эти процесса генетически связаны между собой, и усиление регенерации крови на определенном этапе сопровождается повышением эритродиереза [39]. Наблюдаемая у гудаурской полевки в условиях Центрального Кавказа высокая эритропоэтическая активность свидетельствует о том, что обитание вида в условиях континентального климата требует более напряженного функционирования системы крови, чем в условиях приморского. В результате эритроциты изнашиваются быстрее, и уровень эритродиереза в крови повышается. Хотя общее количество эритроидных клеток в костном мозге полевок обеих центральнокавказских популяций находится на одном уровне, высокое количество эритробластов у животных эльбрусского варианта свидетельствует о большей стимуляции у них эритропоэза, чем у животных терского варианта.
Уровень эритроцитов в крови Ch. gud в трех вариантах поясности существенно различается. Известно, что колебания уровня эритроцитов периферической крови, связанные с видовыми особенностями или вызванные внешними воздействиями, находятся в обратной корреляционной зависимости с активностью каталазы крови. Различным уровням эритроцитов, находящихся в пределах физиологической нормы, соответствуют определенные уровни активности глюкозо-6-фосфат-дегидрогеназы. При относительно низком уровне эритроцитов отмечается относительно высокая гемолитическая стойкость красных кровяных клеток. При их высоком уровне в крови индивидуальный темп распада гемоглобина выше [40]. Следовательно, меньшее количество эритроцитов у полевок Западного Кавказа может быть связано с их большей гемолитической стойкостью по сравнению с таковыми Центрального Кавказа. В таком случае наименее устойчивы клетки животных эльбрусского варианта поясности.
Выявленные нами близкие значения гематокритной величины у всех трех популяций Ch. gud обусловлены обратной зависимостью количества и размеров эритроцитов: чем больше количество клеток, тем меньше их объем и диаметр, и наоборот. Уменьшение диаметра эритроцитов, по мнению А.Д. Панцхава [41], может быть связано с опорожнением кровяных депо и выходом в циркуляцию старых форм – мелких эритроцитов. Соответственно эритроциты меньших размеров, обнаруженные у животных эльбрусского варианта поясности, могут свидетельствовать о выходе в кровоток старых эритроцитов.
Рассмотрим выявленные различия в системе крови гудаурской полевки из разных вариантов поясности в связи с природно-климатическими особенностями этих вариантов. В настоящее время считается общепринятым мнение о комплексном действии факторов в природных условиях, что определяет специфичные условия отдельной горной системы [4, 42–45], при этом ведущая роль отводится низкому парциальному давлению кислорода, т.е. гипоксии.
Как видно из табл. 1, разница по парциальному содержанию кислорода в изученных районах незначительна. По-видимому, она не является значимой для гудаурской полевки, о чем свидетельствует кислородная емкость крови: содержание гемоглобина, гематокритное число и средняя концентрация гемоглобина в эритроците у изученных групп животных не отличаются (см. табл. 3).
Существенно различается среднегодовой уровень осадков на Западном и Центральном Кавказе (см. табл. 1), косвенно свидетельствующий о влажности воздуха, менее значительны различия по среднегодовой температуре. В соответствии с различиями климатических характеристик трех вариантов поясности можно попытаться объяснить выявленные различия в системе крови полевок. Так, при сравнении показателей животных двух популяций (терской и эльбрусской) из континентального подтипа поясности в условиях, характеризующихся разным температурным режимом и сходным уровнем среднегодовых осадков (влажности), обнаружены различия в составе клеток костного мозга, количестве и размерах эритроцитов периферической крови и степени насыщенности их гемоглобином. Соответственно можно полагать, что эти различия определяются температурой.
На Западном Кавказе (плато Лагонаки) выпадает в 1.5 раза больше осадков, чем на Центральном (окр. с. Безенги и п. Эльбрус). При этом среднегодовая температура в окр. плато Лагонаки выше почти на 2°С, чем в окр. п. Эльбрус. Данные особенности климата, вероятно, обусловливают более значительные изменения механизмов поддержания оптимальной кислородной емкости крови. В условиях континентального подтипа поясности кислородная емкость крови поддерживается за счет усиления процесса эритропоэза и увеличения количества мелких эритроцитов, в приморском подтипе – за счет продуцирования костным мозгом качественно иных эритроцитов – более крупных с большим содержанием в них гемоглобина, но в меньшем количестве.
Полученные данные по системе “красной” крови Ch. gud дают основание полагать, что наиболее благоприятными для вида являются условия Западного Кавказа, в пользу чего свидетельствует и проведенный ранее нами анализ состояния иммунной системы вида в тех же условиях [32]. Наши выводы согласуются с утверждением Н.К. Верещагина [21], согласно которому оптимум ареала гудаурской полевки расположен в субальпийском поясе Западного Кавказа.
ЗАКЛЮЧЕНИЕ
Сравнительный анализ комплекса гематологических показателей Ch. gud в условиях Западного и Центрального Кавказа позволил выявить различия в системе “красной” крови количественно-качественного характера, свидетельствующие о том, что у вида на одной высоте (1800 м над ур. м.) в широтно-долготном градиенте, при отсутствии различий в кислородной емкости крови, механизмы обеспечения тканей организма кислородом существенно различаются.
На Западном Кавказе у Ch. gud эритропоэтическая функция костного мозга выражена в меньшей степени, чем на Центральном, возможно потому, что в условиях теплого и влажного климата эритроциты в крови животных более устойчивы, дольше живут и соответственно нет необходимости в продуцировании костным мозгом большого количества эритроцитов. Для удовлетворения кислородного запроса организма в этих условиях достаточно меньшего количества клеток, но качественно иных – более крупных с большим содержанием гемоглобина. В отличие от животных приморского подтипа поясности у животных континентального подтипа обнаружено повышенное количество клеток эритроидного ряда как в костном мозге, так и в периферической крови, свидетельствующее о некотором напряжении в функционировании системы крови. При одинаково высоком уровне эритропоэза костного мозга у полевок в условиях терского и эльбрусского вариантов поясности, характеризующихся различным сочетанием температуры и влажности, наблюдаются существенные различия в парциальной эритрограмме костного мозга. Значительное количество эритробластов – наиболее молодых клеток эритроидного ряда у животных в эльбрусском варианте поясности, не характерное для особей двух других вариантов, свидетельствует о том, что напряжение усиливается при сочетании низких температуры и влажности. На напряженное функционирование системы крови в этих условиях указывает и высокое содержание в периферической крови эритроцитов меньших размеров и с меньшим количеством в них гемоглобина.
При такой значительной изменчивости показателей “красной” крови Ch. gud кислородная емкость крови у особей всех трех вариантов поясности остается на одном уровне: содержание гемоглобина в крови, гематокрит и средняя концентрация гемоглобина в эритроците существенно не различаются. Это свидетельствует о том, что разница в парциальном давлении кислорода в изученных районах не существенна для данного вида. Выявленная изменчивость показателей, направленная на поддержание стабильной концентрации гемоглобина в крови в разных природно-климатических условиях, способствует оптимизации дыхательной функции крови.
Таким образом, суммируя полученные данные, можно заключить, что основными факторами, определяющими выявленные особенности системы крови гудаурской полевки в широтно-долготном градиенте гор Западного и Центрального Кавказа, характеризующихся различными природно-климатическими условиями, на одной высоте, на наш взгляд, могут быть температура и влажность среды или их различные сочетания. Наиболее оптимальным для вида является теплый и влажный климат кубанского варианта приморского подтипа поясности, так как в этих условиях на уровне системы крови проявляются признаки, наиболее выгодные с точки зрения энергозатрат.
Изучение изменчивости системы крови млекопитающих в горах в предложенном аспекте с учетом секторности, на наш взгляд, может существенно дополнить сведения о механизмах обеспечения организма кислородом в горных условиях и расширить представления об адаптационных возможностях животных.
Список литературы
Соколов В.Е., Темботов А.К. Млекопитающие Кавказа: Насекомоядные. М.: Наука, 1989. 548 с.
Темботов А.К., Шебзухова Э.А., Темботова Ф.А. и др. Проблемы экологии горных территорий: Учеб. пос. для студ. Майкоп: Изд-во АГУ, 2001. 186 с.
Шварц С.С., Смирнов В.С., Добринский Л.Н. Метод морфофизиологических индикаторов в экологии наземных позвоночных. Свердловск: Урал. фил. АН СССР, 1968. 387 с.
Большаков В.Н. Пути приспособления мелких млекопитающих к горным условиям. М.: Наука, 1972. 199 с.
Большаков В.Н., Ковальчук Л.А. Адаптационные возможности окислительного метаболизма тканей и системы крови сибирского лемминга // Докл. АН СССР. 1984. Т. 274. № 2. С. 493–496.
Темботова Э.Ж. Экологический анализ системы красной крови малого суслика (Cittelus pygmaeus Pall.) в условиях высотной поясности Северного Кавказа: Автореф. дис. … канд. биол. наук. Свердловск, 1975. 18 с.
Темботова Э.Ж., Улигова З.Н., Барагунова Е.А. и др. Показатели крови некоторых млекопитающих Кавказа // Фауна, экология и охрана животных Северного Кавказа: Межвед. сб. науч. тр. Нальчик: КБГУ, 1980. С. 65–81.
Пантелеев П.А., Терехина А.Н., Варшавский А.А. Экогеографичекая изменчивость грызунов. М.: Наука, 1990. 374 с.
Morrison P., Kerst K., Reynafarje C., Ramos J. Hematocrit and hemoglobin levels in same perinron rodents from high low altitude // International J. of Biometeorology. Springer Berlin. Heidelberg, 1963. V. 7. № 1. P. 51–58.
Wyckoff Susan M., Frasse Barbara A. Hematological adaptation to hypoxia in Peromuscus and Microtus at high and low altitude // Trans. III. State Acad. Sci. 1990. № 3–4. P. 197–205.
Zongkun Zhu, Muyan He, Baolan Zhan et al. Comparison of hypobaric hypoxia reference in hedgehogs, squirrels and rats // Beijing Daxuebao = Acta Sci. Natur. Univ. Perenensis. 1993. V. 29. № 2. P. 184–190.
Ostrojic H., Cifuentes V., Monge C. Hemoglobin affinity in Andean rodent // Biol. Res. 2002. V. 35 (1). P. 27–30.
Storz F. Jay. Hemoglobin function and physiological adaptation to hypoxia in high-altitude mammals // Journal of Mammalogy. 2007. V. 88 (1). P. 24–31.
Ковальчук Л.А. Эколого-физиологические аспекты адаптации к условиям техногенных экосистем. Екатеринбург: УрО РАН, 2008. 216 с.
Барбашова З.И., Тавровская Т.В., Тараканова О.И. и др. Сравнительная оценка механизмов адаптации к гипоксии у горных и равнинных сусликов Северного Кавказа. 1. Состояние эритрона // Экология. 1976. № 2. С. 63–70.
Темботов А.К., Темботова Э.Ж., Берсекова З.А., Емкужева М.М. Влияние градиента высоты местности на гематологические показатели одного из широкораспространенных видов грызунов – малой лесной мыши (Muridae, Rodentia) на Центральном Кавказе // Млекопитающие горных территорий: Мат-лы междунар. конф. М.: КМК, 2005. С. 169–174.
Темботова Э.Ж., Емкужева М.М., Темботова Ф.А. Эколого-физиологический анализ эритрона домовой мыши (Mus musculus L.) в условиях высотной поясности Центрального Кавказа // Млекопитающие горных территорий: Мат-лы междунар. конф. М.: КМК, 2007. С. 315–323.
Емкужева М.М. Сравнительный анализ адаптивных реакций системы крови и интерьерных признаков дикоживущих и синантропных грызунов семейства Muridae к условиям гор центральной части Северного Кавказа: Автореф. дис. … канд. биол. наук. Саратов, 2013. 19 с.
Барагунова Е.А., Лампежева Р.М., Сабанова Р.К. и др. Сравнительная характеристика гематологических и морфофизиологических показателей водяной полевки (Arvicola terrestris rufescens Sat.) горной и предгорной популяций Центрального Кавказа // Современные проблемы науки и образования. 2017. № 3. С. 148–155.
Громов И.М., Поляков И.Я. Фауна СССР. Млекопитающие. Полевки (Microtinae). Л.: Наука, 1977. Т. III. Вып. 8. 504 с.
Верещагин Н.К. Млекопитающие Кавказа. М.; Л.: Изд-во АН СССР, 1959. 702 с.
Курятников Н.Н. К размножению гудаурской полевки на Центральном Кавказе // Вопросы экологии и биологии животных северных склонов Центрального Кавказа. Орджоникидзе, 1976. С. 105–113.
Курятников Н.Н. Некоторые особенности стациального распределения и биологии гудаурской снежной полевки Chionomys gud Sat. на Центральном Кавказе // Экология животных северных склонов Центрального Кавказа. Орджоникидзе, 1978. С. 26–30.
Добролюбов А.Н. Численность, биотопическая приуроченность и морфологические особенности гудаурской снежной полевки из Тебердинского заповедника // Экология, методы изучения и организация охраны млекопитающих горных областей. Свердловск, 1977. С. 32–33.
Маликов В.Г. Сравнительный анализ постнатального онтогенеза полевок родов Microtus и Chionomys (Rodentia, Cricetidae) // Экология и охрана горных видов млекопитающих. М., 1987. С. 112–113.
Саблина О.В., Раджабли С.И., Маликов В.Г. и др. О систематическом положении полевок рода Chionomys по кариологическим данным // Зоол. журн. 1988. Т. 17. Вып. 3. С. 472–475.
Кулиев Г.Н. Гомология хромосом у представителей Chionomys и Pitymys // V съезд Всесоюзного териологического общества АН СССР. М., 1990. С. 74–76.
Хуламханова М.М. Эколого-биологические особенности гудаурской полевки (Chionomys gud Satunin, 1909) на Кавказе: Автореф. дис. … канд. биол. наук. Махачкала, 2007. 22 с.
Дзуев Р.И., Сижажева А.М., Банникова А.А. Внутривидовая генетическая структура кавказской снеговой полевки (Chionomys gud Satunin, 1909) по результатам анализа митохондриальной ДНК // Изв. КБГУ. Нальчик: Каб.-Балк. ун-т, 2011. Т. I. № 4. С. 35–42.
Сижажева А.М. Молекулярно-генетическое и экологическое разнообразие рода Chionomys на Кавказе: Автореф. дис. … канд. биол. наук. Владикавказ, 2012. 23 с.
Банникова А.А., Сижажева А.М., Маликов В.Г. и др. Генетическое разнообразие рода Chionomys (Mammalia, Arvicolinae) и сравнительная филогеография трех видов снеговых полевок // Генетика. 2013. Т. 49. № 5. С. 649–664.
Боттаева З.Х., Темботова Ф.А., Берсекова З.А. и др. Эколого-географическая изменчивость иммунологических показателей периферической крови гудаурской полевки (Cricetidae, Rodentia) в среднегорьях Западного и Центрального Кавказа // Изв. Самарского научного центра РАН. 2016. Т. 18. № 5. С. 83–88.
Дзуев Р.И., Хуламханова М.М., Сижажева А.М. Молекулярная систематика и эколого-биологические особенности гудаурской полевки (Chionomys gud Satunin, 1909) на Кавказе. Махачкала: Эко-экспресс, 2011. 208 с.
Климатические данные городов по всему миру: http://ru.climat-data.org./.
Карасева Е.В., Телицына А.Ю., Жигальский О.А. Методы изучения грызунов в полевых условиях. М.: Изд-во ЛКИ, 2008. 416 с.
Тодоров И. Клинические лабораторные исследования в педиатрии. София: Медицина и физкультура, 1968. 1063 с.
Анализы. Полный справочник / Под ред. Елисеевой Ю.Ю. М.: Изд-во “Эксмо”, 2006. 768 с.
Тарахтий Э.А., Давыдова Ю.А. Сезонная изменчивость показателей системы крови рыжей полевки (Clethrionomys glareolus) разного репродуктивного состояния // Изв. РАН. Сер. биол. 2007. № 1. С. 14–25.
Горбунова Н.А. Эритродиерез при экстремальных воздействиях и его роль в регенерации крови // Гематология и трансфузиология. 1985. № 2. С. 23–27.
Клиорин А.И., Тиунов Л.А. Функциональная неравнозначность эритроцитов. Л.: Наука, 1974. 148 с.
Панцхава А.Д. Качественные изменения эритроцитов в периферической крови. Тбилиси: Саббсота Сакартвело, 1961. 66 с.
Темботов А.К. География млекопитающих Северного Кавказа. Нальчик: “Эльбрус”, 1972. 245 с.
Барбашова З.И. Акклиматизация к гипоксии и ее физиологические механизмы. М.; Л.: Изд-во АН СССР, 1960. 216 с.
Булатова Н.Н. Особенности высокогорных животных // Тр. Ин-та морфологии животных им. А.Н. Северцова. 1962. Вып. 41. С. 11–43.
Экологическая физиология животных. Ч. III. Физиология животных в различных физико-географических зонах / Под ред. Слонима А.Д. Л.: Наука, 1982. 504 с.
Дополнительные материалы отсутствуют.