

Экология, 2019, № 1, стр. 40-46
Питание бородатой неясыти Strix nebulosa при разной численности жертв в период гнездования
Ю. Э. Кропачева a, *, Н. Г. Смирнов a, **, С. В. Зыков a, М. И. Чепраков a, Н. О. Садыкова a, Г. Н. Бачурин b
a Институт экологии растений и животных УрО РАН
620144 Екатеринбург, ул. 8 Марта, 202, Россия
b Научно-практический центр биоразнообразия
623853 Ирбит, ул. Мира, 56, Россия,
* E-mail: KropachevaJE@yandex.ru
** E-mail: nsmirnov@ipae.uran.ru
Поступила в редакцию 20.06.2018
После доработки 25.06.2018
Принята к публикации 20.06.2018
Аннотация
Исследована изменчивость рациона одного из специализированных миофагов – бородатой неясыти – на Среднем Зауралье при разном уровне численности жертв в различных биотопических условиях. Основными жертвами были серые полевки, альтернативными – бурозубки. В течение гнездового периода на разных участках отмечены два типа изменения рациона: 1) замещение серых полевок луговых биотопов серыми полевками, населяющими лесные местообитания; 2) замещение серых полевок лесных местообитаний бурозубками.
Изменчивость рациона как один из механизмов адаптации сов к меняющимся условиям широко исследуется в орнитологии [1–4]. Данная проблема рассматривается как в географическом, так и во временном аспектах [1, 2, 5, 6]. Питание бородатой неясыти хорошо изучено в Фенноскандии, Белоруссии, США и Канаде [3, 4, 7–9 и др.]. В России питание этого вида описано для нескольких регионов [10–13]. По характеру кормодобывания бородатая неясыть находится ближе к специализированным хищникам по сравнению с серой (Strix aluco) и длиннохвостой (Strix uralensis) неясытями [3, 7, 8, 14]. На большей части ареала ее основными жертвами являются серые полевки, однако в связи с широким циркумбореальным распространением описаны и различия в рационе [3, 4, 7, 10, 11]. Оптимальные местообитания вида – перестойная тайга в сочетании с открытыми пространствами: болотами, гарями, пустошами, лугами [3–5, 7, 10]. Бородатая неясыть добывает жертв, населяющих как лесные, так и открытые биотопы [3, 4, 7, 12, 13]. Как специализированный хищник она зависит от количества предпочитаемой добычи. Для нее характерны миграции или изменения параметров гнездования в зависимости от численности жертв (численный ответ) [4, 6]. Изменение рациона в связи с уменьшением численности предпочитаемых жертв (функциональный ответ) менее характерно для данного вида и мало изучено.
Цель настоящей работы – изучить питание бородатой неясыти в зависимости от численности жертв на примере Среднего Зауралья. Для этого необходимо было оценить рацион сов: 1) гнездившихся на участках с разными биотопическими характеристиками; 2) при разном уровне численности жертв; 3) в разные периоды гнездования.
МАТЕРИАЛ И МЕТОДЫ
Район исследования. Работы проводили с 2015 по 2017 г. в Ирбитском районе Свердловской области, Россия (57°40′ с.ш., 62°30′ в.д.). Результаты сравнения питания бородатой неясыти в 1978 и 2015 гг. опубликованы ранее [12]. Растительность изучаемой территории представлена разновозрастными и разнопородными участками леса, перемежающимися с луговыми территориями. Здесь на территории КОТР СВ-001 “Лесной массив близ пос. Зайково” были установлены искусственные гнезда для бородатой неясыти [12].
Используемые в работе жилые гнезда располагались на участках (территория вокруг гнезд радиусом 1.5 км), различающихся по соотношению разных типов местообитаний. На участке 1 преобладают луга с многочисленными сосново-березовыми колками (3 гнезда). Участок 2 представлен в основном лесными местообитаниями – он располагается в сплошном лесном массиве с зарастающей вырубкой и небольшими лесными полянами (1 гнездо). На участке 3 преобладающими по площади являются лесные местообитания, но присутствуют и луговые территории (2 гнезда). Участок 4 расположен на краю лесного массива, примыкающего к заброшенным сельскохозяйственным угодьям (1 гнездо). Расстояние между гнездами на одном участке 0.8–1.5 км, между гнездами на разных участках – от 2.2 до 9 км.
Анализ погадок. Питание в гнездовой период в 2015 г. изучено у одной пары, в 2016 г. – у трех, в 2017 г. – у шести пар птиц. Свежие погадки собирали под присадами возле гнезд в 2015 и 2016 гг. однократно в мае, в 2017 г. – 4–6 раз в течение гнездования (май и июнь). Собирали все погадки самок, самцов и птенцов. За основу анализа взяты сборы 2017 г., так как в этот год наблюдалось наибольшее число заселенных гнезд (6 шт.), а кормовая база обусловила успешность гнездования. Сравнение питания сов, гнездившихся на разных участках, проведено по совокупности синхронных материалов (мая и июня) одного года (2017 г.), чтобы избежать влияния межгодовых различий. Питание сов, гнездившихся на одном участке в разные годы, сравнивали только для сборов мая. Так как в 2017 г. птенцы вылупились в конце мая, собранные материалы разделили на 2 периода – до конца мая (период высиживания яиц) и в июне (период выкармливания птенцов). Сравнение питания сов в 2017 г. проведено для участков 1, 2, 3. Общая характеристика питания – состав и соотношение жертв – дана по всем имеющимся выборкам 2015–2017 гг.
Костные остатки из погадок были очищены вручную. Грызунов идентифицировали до вида [15, 16], насекомоядных – до рода. По максимальному количеству одноименных остатков (нижних челюстей) определено число особей. Из 372 погадок определен 3191 остаток 1323 особей животных. Виды жертв были сгруппированы по биотопическому и систематическому принципу: 1) серые полевки, обитающие в луговых биотопах (луга, сельхозугодия), – обыкновенная (Microtus arvalis sensu lato) и узкочерепная (Microtus gregalis); 2) серые полевки лесных местообитаний (заболоченные участки, пойменный кочкарник, лесные поляны, зарастающие вырубки) – полевка-экономка (Microtus oeconomus) и темная полевка (Microtus agrestis); 3) лесные полевки – красная (Clethrionomys rutilus) и рыжая (Clethrionomys glareolus); 4) малая лесная мышь (Sylvaemus uralensis); 5) землеройки (Soricidae); 6) другие жертвы, остатки которых в погадках единичны или малочисленны: ондатра (Ondatra zibethicus), лесной лемминг (Myopus schisticolor), водяная полевка (Arvicola terrestris), лесная мышовка (Sicista betulina), большой суслик (Spermophilus major), ласка (Mustela nivalis), лягушки (Rana sp.).
Оценка численности мелких млекопитающих. В 2016 и 2017 гг. были проведены отловы мелких млекопитающих живоловушками с возвратом в среду обитания: в 2016 г. – на участках 1 и 2 однократно, так как к началу июня совы, не завершив гнездование, покинули гнезда; в 2017 г. – на участках 1, 2, 3 дважды – в мае и июне. Ловушки расставляли в пределах 1 км2 вокруг жилых гнезд на 2–4 сут в луговых и лесных биотопах (сухие и заболоченные участки, опушки, заросли кустарников) и проверяли дважды в сутки. Пойманных животных метили ампутацией пальцев. У серых полевок для видовой идентификации снимали прижизненные слепки жевательной поверхности коренных зубов [17]. Для этой процедуры использовали стоматологические силиконы Bisico S1 (базовый) и S4 (коррегирующий). Подсчитано количество особей на 100 ловушко-суток (ос/л-с) на каждом участке – отдельно для лесных и луговых линий отловов. Всего отловлено и идентифицировано 180 особей, которых при описании материала объединили в те же группы, что и при анализе погадок: 1 – обыкновенная и узкочерепная полевки; 2 – экономка и темная полевка; 3 – красная и рыжая полевки; 4 – малая лесная и полевая (Apodemus agrarius) мыши; 5 – землеройки.
Статистический анализ. Численность мелких млекопитающих в разные годы и месяцы одного года сравнивали с помощью t-теста. В качестве повторностей использованы линии отловов. Для сравнения рациона сов с разных участков, в разные годы и периоды гнездования одного года, а также для сравнения результатов отловов на разных участках применяли тест χ2. В эти сравнения включены перечисленные группы видов, составляющие не менее 10% в погадках и/или отловах. Сравнения проведены по средним значениям для гнезд и линий отловов (отдельно лесных и луговых) на одном участке. Отношения между долями отдельных групп жертв в рационе оценены с помощью коэффициента корреляции Спирмена. Средние значения приведены с величиной стандартного отклонения (SD).
РЕЗУЛЬТАТЫ
Численность мелких млекопитающих в 2016–2017 гг. Численность мелких млекопитающих (по результатам отловов) в мае 2016 и 2017 гг. различалась незначимо (t = 2.23, n1 = 4, n2 = 9, p = 0.99), а соотношение групп видов – значимо (χ2 = 16.9, df = 4, p = 0.01). Весной 2016 г. мелкие млекопитающие в отловах в луговых биотопах отсутствовали, в лесных биотопах их численность была 6.25 ± ± 6 ос/100 л-с, преобладали мыши. В мае 2017 г. в луговых биотопах численность составила 3.86 ± ± 1.23 ос/100 л-с, представлены обыкновенная и узкочерепная полевки (рис. 1). В лесных биотопах при численности 2.55 ± 1.97 ос/100 л-с доминировали лесные полевки. В конце мая 2017 г. в отловах отмечены сеголетки серых полевок. В июне по сравнению с маем численность мелких млекопитающих на изучаемой территории значимо выросла (t = 2.31, n1 = 9, n2 = 9, p = 0.05). Тенденция более выражена у полевок, обитающих на лугах, хотя для отдельных биотопов изменения численности незначимы (луг: t = 2.23, n1 = 4, n2 = 4, p = 0.06; лес: t = 1.38, n1 = 5, n2 = 5, p = 0.22). На лугах численность составила 10.48 ± 5.80 ос/л-с, тогда как в лесных местообитаниях 4.86 ± 3.34. Соотношение групп видов значимо не изменилось (χ2 = 0.25, df = 4, p = 0.99) (см. рис. 1).
Рис. 1.
Соотношение групп мелких млекопитающих в отловах: 1 – Microtus arvalis, M. gregalis; 2 – M. oeconomus, M. agrestis; 3 – Clethrionomys sp.; 4 – Sylvaemus uralensis, 5 – Sorex sp.
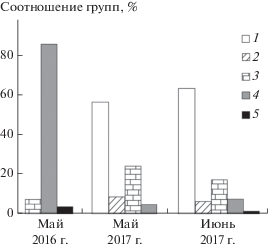
Результаты отловов в 2016 и 2017 гг. на охотничьих участках бородатой неясыти приведены в табл. 1. Различия численности на разных участках в 2017 г. были незначимы (уч. 1 и 2: t = 0.37, n1 = 8, n2 = 2, p = 0.72; уч. 1 и 3: t = 0.46, n1 = 8, n3 = 8, p = = 0.05; уч. 2 и 3: t = 0.44, n2 = 2, n3 = 8, p = 0.05).
Таблица 1.
Число особей мелких млекопитающих на 100 л-с в отловах живоловушками на охотничьих участках бородатых неясытей
| Таксон | 2016 г. | 2017 г. | |||
|---|---|---|---|---|---|
| уч. 1 | уч. 2 | уч. 1 | уч. 2 | уч. 3 | |
| Microtus arvalis sensu lato | 0 | 0 | 2.82 | 0 | 7.78 |
| M. gregalis | 0 | 0 | 3.74 | 0 | 0 |
| M. oeconomus | 0 | 0 | 0.1 | 0 | 0 |
| M. agrestis | 0 | 0 | 0.59 | 1.83 | 0 |
| Clethrionomys rutilus | 0 | 0.5 | 3.0 | 1.67 | 0.24 |
| Cl. glareolus | 0 | 0.5 | 0.29 | 1.5 | 0.36 |
| Sylvaemus uralensis | 2.0 | 8.5 | 0.22 | 1.5 | 0.72 |
| Apodemus agrarius | 0 | 0.5 | 0 | 0 | 0 |
| Sorex sp. | 0 | 0.5 | 0.38 | 0 | 0 |
| Количество особей на 100 л-с (количество отловленных особей) | 2(2) | 10.5(21) | 11.14(71) | 6.5(39) | 9.1(47) |
Состав рациона. Рацион бородатых неясытей в период гнездования в 2017 г состоял в основном из серых полевок (83%), из них примерно равные доли составляли серые полевки, обитающие в луговых (44%) и лесных местообитаниях (39%), далее следуют землеройки (10%), лесные полевки (4%) и остальные виды (3%).
Питание неясытей, гнездившихся на разных участках в 2017 г. Питание сов, гнездившихся на разных участках в 2017 г. (май и июнь), значимо различалось (уч. 1 и 2: χ2 = 25.42, df = 4, p = 0.01; уч. 1 и 3: χ2 = 31.75, df = 4, p < 0.001; уч. 1 и 4: χ2 = = 63.88, df = 4, p <0.001; уч. 2 и 3: χ2 = 72.52, df = 4, p <0.001; уч. 2 и 4: χ2 = 27.0, df = 4, p = 0.01; уч. 3 и 4: χ2 = 37.03, df = 3, p < 0.001). В рационе на участке 1 преобладали экономка и темная полевки (57%), далее следовали обыкновенная и узкочерепная (32%). На участке 2 доминировали экономка и темная полевки (69%), значительна была доля бурозубок (13%) и обыкновенной полевки (11%). На участке 3 доминировала обыкновенная полевка (55%), далее следовали экономка и темная полевки (31%). На участке 4 примерно равные доли составляли бурозубки (39%) и узкочерепная полевка (34%) (табл. 2).
Таблица 2.
Число особей жертв в погадках на разных участках в период гнездования бородатых неясытей (2015–2017 гг.)
| Таксон | 2015 г. | 2016 г. | 2017 г. | ||||
|---|---|---|---|---|---|---|---|
| уч. 1 | уч. 1 | уч. 2 | уч. 1 | уч. 2 | уч. 3 | уч. 4 | |
| Microtus arvalis s. l. | 11 | 13 | 0 | 68 | 13 | 227 | 0 |
| M. gregalis | 25 | 83 | 1 | 82 | 1 | 1 | 14 |
| M. oeconomus | 21 | 40 | 10 | 174 | 42 | 73 | 4 |
| M. agrestis | 7 | 10 | 8 | 94 | 45 | 55 | 1 |
| Clethrionomys rutilus | 2 | 2 | 1 | 8 | 1 | 7 | 2 |
| C. glareolus | 1 | 1 | 0 | 6 | 2 | 7 | 3 |
| Arvicola terrestris | 0 | 1 | 1 | 1 | 0 | 0 | 0 |
| Myopus schisticolor | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Sylvaemus uralensis | 0 | 0 | 1 | 2 | 2 | 0 | 0 |
| Sciurus vulgaris | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Spermophilus major | 0 | 0 | 0 | 0 | 0 | 0 | 1 |
| Sicista betulina | 0 | 1 | 0 | 6 | 1 | 14 | 0 |
| Ondatra zibethicus | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Mustela nivalis | 0 | 0 | 0 | 2 | 0 | 1 | 0 |
| Sorex sp. | 0 | 6 | 31 | 23 | 17 | 25 | 16 |
| Rana sp. | 0 | 0 | 0 | 3 | 2 | 0 | 0 |
| Количество погадок | 14 | 38 | 15 | 154 | 45 | 96 | 10 |
| Количество определенных остатков | 167 | 401 | 130 | 1148 | 307 | 938 | 100 |
| Количество особей | 67 | 157 | 53 | 469 | 126 | 410 | 41 |
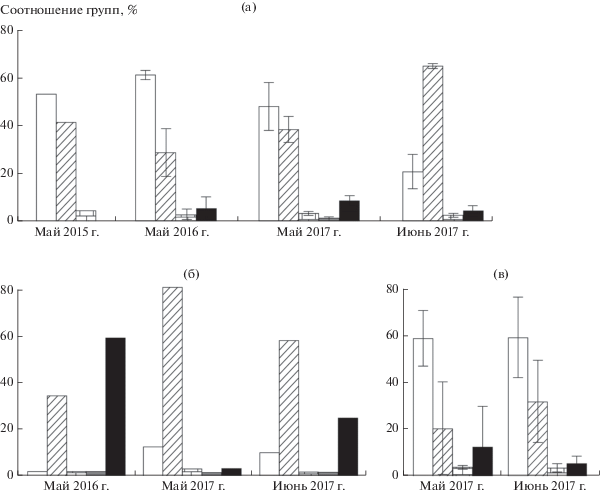
Различия в питании между годами и в течение гнездования одного года. Рацион сов, селившихся на участке 1 в мае разных лет, значимо не различался (2015 и 2017 г.: χ2 = 6.6, df = 4, p = 0.9; 2016 и 2017 г.: χ2 = 3.9, df = 4, p = 0.90) (рис. 2а). В майских выборках 2015 г. зафиксировано 6 видов грызунов, в 2016 г. – 8 видов грызунов и бурозубки, в 2017 г. – 7 видов грызунов и бурозубки. Рационы сов значимо различались в мае и июне 2017 г. (χ2 = 18.66, df = 4, p = 0.01) (см. рис. 2а). В июне было 8 видов грызунов, ласка, бурозубки и лягушки. Питание сов, гнездившихся на участке 2, значимо различалось в разные годы (2016 и 2017 г.: χ2 = 46.91, df = 4, p < 0.001) (рис. 2б). В мае 2016 г. отмечены 6 видов грызунов и бурозубки, в мае 2017 г. – 6 видов грызунов и бурозубки (см. табл. 2). Соотношение групп видов значимо поменялось в июне (χ2 = 13.53, df = 4, p = 0.01): обнаружены 6 видов грызунов, бурозубки и лягушки (см. рис. 2б). В рационе сов, селившихся на участке 3, в мае 2017 г. было 6 видов грызунов, ласка и бурозубки, в июне – 7 видов грызунов и бурозубки. Изменения долей групп жертв незначимы (χ2 = 0.56, df = 4, p = 0.1) (рис. 2в).
Рис. 2.
Соотношение групп мелких млекопитающих в питании бородатой неясыти: а – участок 1; б – участок 2; в – участок 3. Обозначения видов см. на рис. 1.
Взаимосвязь групп жертв на исследуемой территории. В рационе неясытей доля обыкновенной и узкочерепной полевок отрицательно коррелирует с долей экономки и темной полевки (Spearman rank correlations, rS = –0.82, p < 0.05) и не коррелирует с долей бурозубок (rS = 0.14, p = 0.61). Доля экономки и темной полевки отрицательно коррелирует с долей бурозубок (rS = –0.56, p < 0.05).
ОБСУЖДЕНИЕ
Сопоставление видового состава жертв сов с фауной региона. В погадках неясытей зафиксировано 14 видов грызунов. Все они отмечены в литературе для данной территории. Отсутствовали в добыче летяга, бурундук, мышь-малютка, обыкновенный хомяк – виды повсеместно редкие в Свердловской области [16]. Из особенно редких видов для данной местности в погадках отмечен лесной лемминг.
Рацион сов на разных участках. При наличии лугов на участках (уч. 1, 3, 4) в рационе бородатой неясыти серые полевки, селящиеся в луговых биотопах (обыкновенная и узкочерепная полевки), доминировали над серыми полевками лесных местообитаний (темная полевка и экономка) (в 6 из 7 выборок). Бородатая неясыть предпочитала добывать полевок, обитающих в открытых биотопах, другим видам жертв и в других частях ареала [4, 10, 11, 13]. При отсутствии лугов на участке обитания (уч. 2) в питании преобладали серые полевки, селящиеся в лесных биотопах. На значительной части европейского ареала в таежной зоне, где бородатая неясыть тяготеет к болотам, травянистым участкам, гарям, темная полевка и в меньшей степени полевка-экономка являются ее основными жертвами [3, 4, 7, 11, 13]. Вторая по численности группа жертв на данном участке – бурозубки. В рационе бородатой неясыти это постоянная, но не доминирующая группа жертв на всем ареале [3, 5, 7, 8].
Основные и альтернативные жертвы. С учетом представлений об основных и альтернативных жертвах [2, 18, 19] можно заключить, что на изучаемой территории, как и на большей части ареала [3, 4, 7], основными жертвами бородатой неясыти являются серые полевки. Соотношения двух биотопических групп серых полевок в ее рационе и причины их изменений на изучаемой территории требуют отдельного исследования. Можно рассматривать такие факторы, как предпочитаемый совами биотоп для охоты, численность видов из этих групп, размер и социальная структура жертв [20]. Для изучаемой территории характерна следующая взаимосвязь групп жертв: доля видов, образующих поселения на лугах, – обыкновенной и узкочерепной полевки, отрицательно коррелирует с долей серых полевок, населяющих лесные биотопы, – экономки и темной полевки, которая в свою очередь отрицательно связана с долей бурозубок.
На данном этапе исследования, основываясь на пропорциях групп видов и их взаимосвязи, можно предположить, что наиболее доступными и, вероятно, предпочитаемыми жертвами на изучаемой территории являются серые полевки, образующие поселения на лугах. Следующая по предпочитаемости группа – серые полевки лесных местообитаний. Альтернативными жертвами можно считать бурозубок. Это согласуется с данными по питанию бородатой неясыти на других частях ареала [3, 4, 7, 10, 11, 15] и результатами исследований, проведенных с использованием радиодатчиков, которые показали, что бородатая неясыть предпочитает охотиться на лугах и лесных участках без подлеска [4, 20].
Межгодовая динамика рациона. На луговом участке (уч. 1) в 2015–2017 гг. в сравнении с 1978 г. доминирующая группа в питании бородатой неясыти – серые полевки луговых местообитаний – осталась та же, возросла роль серых полевок лесных биотопов. В 1978 г. около 10% рациона составляла водяная полевка [12], тогда как в 2015–2017 гг. ее остатки в погадках были единичными. Вероятно, данные изменения связаны с колебаниями численности этих видов на изучаемой территории. В 2016 и 2017 гг. по сравнению с 1978 и 2015 гг. [12] в питании отмечены большее количество видов грызунов, а также ласка, бурозубки и лягушки. Согласно теории оптимального рациона, определяющим является количество основных жертв, при их недостатке в питание включаются альтернативные, и видовое разнообразие жертв возрастает [21]. Можно предположить, что в годы, когда в питании отсутствовали альтернативные жертвы – бурозубки и отмечено меньшее число жертв (1978 и 2015 гг.), численность основных жертв была достаточно высокой.
При исключительно бедной кормовой базе в 2016 г. перехода на менее предпочитаемые группы жертв на луговом участке (уч. 1) не произошло, по-видимому, из-за их отсутствия. На лесном участке (уч. 2) в мае 2016 г. совы ответили на недостаточное количество основных жертв функционально – перешли на альтернативные жертвы (бурозубок). В конце мая последовал численный ответ – совы оставили гнезда на обоих участках.
Динамика рациона в течение гнездования 2017 г. С мая по июнь 2017 г. в отловах отмечено небольшое увеличение численности мелких млекопитающих. Эта тенденция была более выражена у полевок, обитающих на лугах, чем у видов, населяющих лесные биотопы. Соотношение групп видов в отловах не менялось, тогда как в течение гнездования изменились соотношения групп жертв в рационе сов, селившихся на участках 1 и 2: на уч. 1 доля наиболее предпочитаемой добычи – серых полевок открытых биотопов – снизилась, доля второй по предпочитаемости группы – серых полевок лесных местообитаний – возросла; на уч. 2 доля серых полевок лесных местообитаний уменьшилась, тогда как доля альтернативных жертв – бурозубок – увеличилась.
Для птенцовых птиц наиболее энергозатратная стадия гнездования – период выкармливания птенцов [22]. В этот период (июнь) по сравнению с периодом высиживания яиц (май) у сов увеличилась потребность в количестве корма [22, 23]. Вероятно, обилия предпочитаемых жертв оказалось недостаточно, несмотря на небольшое увеличение численности, зафиксированное в отловах. Это повлекло функциональный ответ, реализованный с помощью расширения охотничьей территории и включения менее предпочитаемых биотопов для охоты. В результате наблюдалось перераспределение долей биотопических групп серых полевок на уч. 1 и увеличение альтернативных жертв на уч. 2. На лесном участке с луговыми территориями (уч. 3) не наблюдалось значительных изменений в рационе с мая по июнь 2017 г.
ЗАКЛЮЧЕНИЕ
На исследованной территории основные жертвы бородатой неясыти – серые полевки, альтернативные – бурозубки. При наличии лугов на участке обитания в рационе преобладали серые полевки, населяющие открытые пространства, – обыкновенная и узкочерепная полевки. В случае отсутствия на участке лугов в питании доминировали серые полевки, обитающие в лесных биотопах, – темная полевка и полевка-экономка. В течение гнездового периода наблюдалось несколько вариантов изменения кормодобывающего поведения. Отмечены два типа изменения рациона: 1) замещение серых полевок открытых пространств серыми полевками, населяющими лесные биотопы; 2) замещение серых полевок лесных местообитаний бурозубками. Крайним вариантом реакции на недостаточную кормовую базу было оставление совами гнезд при переходе от стадии насиживания к стадии выкармливания.
Таким образом, пополнен массив данных по питанию бородатой неясыти, который закрывает белое пятно на Урало-Западно-Сибирской части циркумбореального ареала. Сведения для разных местообитаний дополнены данными об особенностях питания сов при разной численности жертв.
Авторы выражают благодарность В.Н. Бачурину и И.А. Сморкалову за помощь в проведении полевых работ. Работа выполнена в рамках государственного задания Института экологии растений и животных УрО РАН, а также частично поддержана Комплексной программой УрО РАН (проект № 18-4-4-3) и грантом РФФИ (№ 16-04-01017).
Список литературы
Korpimaki E., Norrdahl K. Numerical and functional responses of kestrels, short-eared owls and long-eared owls to owl densities // Ecology. 1991. V. 72(3). P. 814–826.
Zárybnická M., Sedláček O., Korpimäki E. Do Tengmalm’s Owls alter parental feeding effort under varying conditions of main prey availability? // J. of Ornithology. 2009. V. 150(1). P. 231–237.
Mikkola H. Der Bartkauz Strix nebulosa. Wittenberg-Lutherstadt: Ziemsen, 1981. 124 p.
Bull E.L., Duncan J.R. Great Gray Owl (Strix nebulosa) // The birds of North America / Eds. Poole A. and Gill F. Philadelphia, Pennsylvania, USA: Academy of Natural Sciences. 1993. № 41. P. 1–15.
Sulkava S., Huhtala K. The Great Grey Owl (Strix nebulosa) in the changing forest environment of northern Europe // J. of Raptor Research. 1997. V. 31. P. 151–159.
Hipkiss T., Stefansson O., Hörnfeldt B. Effect of cyclic and declining food supply on great grey owls in boreal Sweden // Canad. J. of Zoology. 2008. V. 86(12). P. 1426–1431.
Duncan J., Hayward P.H. Review of technical knowledge: great gray owls // Flammulated, Boreal, and Great Gray Owls in the United States: a technical conservation assessment. 1994. V. 253. 159 p.
Tishechkin A.K. Comparative food niche analysis of Strix owls in Belarus // Proc. symp. ecology and conservation of owls of Northen Hemisphere. Winnipeg, 1997. P. 456–460.
Solheim R., Stefansson O. Life span, dispersal and age of nesting Great Grey Owls Strix nebulosa lapponica in Sweden // Ornis Svecica. 2016. V. 26. P. 125–134.
Карякин И.В. Бородатая неясыть // Пернатые хищники Уральского региона: Соколообразные (Falconiformes) и Совообразные (Strigiformes). Пермь: Центр полевых исследований Союза охраны животных Урала, 1998. С. 385–395.
Пукинский Ю.Б. Бородатая неясыть. Птицы России и сопредельных регионов: Совообразные, Козодоеобразные, Стрижеобразные, Ракшеобразные, Удодообразные, Дятлообразные. М.: КМК, 2005. С. 86–98.
Смирнов Н.Г., Кропачева Ю.Э., Бачурин Г.Н. Динамика современной фауны грызунов предлесостепных лесов Зауралья // Фауна Урала и Сибири. 2015. № 1. С. 167–175.
Шепель А.И. Бородатая неясыть Strix nebulosa Forster, 1772 в Волжско-Камском крае // Вестн. Удмуртского ун-та. Сер. “Биология. Науки о Земле”. 2011. Т. 4. С. 85–89.
Korpimaki E. Niche relationships and life-history tactics of three sympatric Strix owl species in Finland // Ornis Scandinavica. 1986. V. 17. P. 126–132.
Бородин А.В. Определитель зубов полевок Урала и Западной Сибири (поздний плейстоцен–современность). Екатеринбург: УрО РАН, 2009. 100 с.
Большаков В.Н., Бердюгин К.И., Васильева И.А., Кузнецова И.А. Млекопитающие Свердловской области: Справочник-определитель. Екатеринбург: “Екатеринбург”, 2000. 240 с.
Оленев Г.В. Определение видовой принадлежности мышевидных грызунов по отпечаткам зубов у живых животных // Зоол. журн. 1980. № 2. С. 294–295.
Salamolard M., Butet A., Leroux A., Bretagnolle V. Responses of an avian predator to variations in prey density at a temperate latitude // Ecology. 2000. V. 81. P. 2428–2441.
Comay O., Dayan T. What determines prey selection in owls? Roles of prey traits, prey class, environmental variables, and taxonomic specialization // Ecology and Evolution. 2018. V 8(6). P. 3382–3392.
Van Riper C., Van Wagtendonk J. Home range characteristics of Great Gray Owls in Yosemite National Park, California // J. of Raptor Research. 2006. V. 40(2). P. 130–141.
Pulliam H.R. On the theory of optimal diets // Amer. Nat. 1974. V. 108. P. 59–74.
Wellicome T.I., Danielle Todd L., Poulin R.G. et al. Comparing food limitation among three stages of nesting: supplementation experiments with the burrowing owl // Ecology and Evolution. 2013. V. 3(8). P. 2684–2695.
Franclin A.B. Breeding biology of the Great Gray Owl in Southeastern Idaho and Northwestern Wyoming // The Condor. 1988. V. 90(3). P. 689–696.
Дополнительные материалы отсутствуют.