

Экология, 2019, № 3, стр. 189-195
Cтруктура ценопопуляций ключевых видов черневых кедровых лесов Западного Саяна на климаксовой стадии развития
М. Е. Коновалова a, *, Д. М. Данилина a, **
a Институт леса им. В.Н. Сукачева СО РАН – обособленное подразделение ФИЦ КНЦ СО РАН
660036 Красноярск, Академгородок, 50/28, Россия
* E-mail: markonovalova@mail.ru
** E-mail: dismailova@mail.ru
Поступила в редакцию 18.04.2018
После доработки 09.07.2018
Принята к публикации 14.08.2018
Аннотация
На примере климаксового кедровника осочково-крупнотравно-папоротникового (фонового типа леса для черневых лесов Западного Саяна) изучены онтогенетические спектры и возрастная структура ценопопуляций ключевых видов: Pinus sibirica (Du Tour) и Abies sibirica (Ledeb.). Выявлены темпы их онтогенетического развития в данных лесорастительных условиях. Ценопопуляции P. sibirica и A. sibirica являются устойчивыми, хотя и отличаются по структуре. Показан механизм устойчивости сообществ, обеспечиваемый сложным взаимодействием видов ценозообразователей в условиях конкуренции со стороны мощно развитой травяной растительности.
Коренные черневые пихтово-кедровые леса северного макросклона Западного Саяна благодаря многовековому отсутствию пирогенного фактора в избыточно влажном климате имеют девственный, почти первобытный характер сообществ. Их отличает от зеленомошной тайги особая фитоценотическая структура [1–3] и сохранившиеся в составе наиболее древние для Сибири виды широкотравья и крупных папоротников, относимые к неморальным реликтам [4]. На данной территории на протяжении XX в. отмечалось возрастание антропогенной нагрузки [5]. Для оценки ее влияния и прогноза состояния природных экосистем требуются мониторинговые исследования и разработки базовых критериев. Одним из таких интегральных показателей считается структура популяций древесных растений, являющихся основными ценозообразователями (возрастная, размерная, онтогенетическая, виталитетная и т.д.). Однако для лесных экосистем черневого пояса Западного Саяна ранее не проводилась оценка онтогенетической структуры популяций видов ценозообразователей и их взаимодействия во времени.
Цель настоящей работы состояла в анализе онтогенетической и возрастной структуры ценопопуляций ключевых видов климаксовых черневых кедровников осочково-крупнотравно-папоротникового типа леса Западного Саяна.
МАТЕРИАЛ И МЕТОДЫ
Исследовали ценопопуляции Pinus sibirica Du Tour и Abies sibirica Ledeb. в климаксовом кедровнике осочково-крупнотравно-папоротниковом. Этот тип леса является фоновым в пределах черневого пояса Джебашско-Амыльского лесорастительного округа Западного Саяна, распространенного на высотах от 350 до 800 м над ур. м. [1]. Район исследований характеризуется континентальным избыточно влажным (циклоническим) климатом [2]. Кедровники отличаются высокой продуктивностью (Iа–II классы бонитета) и низкой сомкнутостью древостоя (относительная полнота 0.4–0.6); большими показателями возраста старшего поколения P. sibirica (свыше 450 лет), крупномерностью его стволов (диаметр до 1 м, высота до 45 м); мощно развитой травяной растительностью и слабым естественным возобновлением P. sibirica [5, 6].
Исследования проводили на постоянной пробной площади № 3 (ПП) размером 1.56 га Ермаковского стационара Института леса им. В.Н. Сукачева СО РАН, заложенной в 1960 г. Она расположена в бассейне р. Малый Кебеж (Кулумысский хребет), на нижней части склона юго-западной экспозиции, крутизной 22–25°, на абсолютной высоте 530 м (53°01′ с.ш., 92°59′ в.д.). Почва горная лесная бурая высокощебнистая на делювии хлоритовых сланцев, сформирована в условиях промывного режима и хорошего дренажа, развитого внутрипочвенного стока.
Фитоценоз обладает всеми признаками позднесукцессионного сообщества: имеет следы прошлых поколений всех доминантных видов, абсолютную разновозрастность и сложную мозаику ключевых видов, а также связанных с ними подчиненных видов. Большой размер ПП позволил учесть все элементы горизонтальной мозаики биогеоценоза. Последняя проявляется в неравномерной сомкнутости древесного полога (0.3–0.9) и разнообразии устойчивых микрогруппировок растений (соответствующих понятию “синузий” по В.Н. Сукачеву [7]), обособленных флористически, экологически и пространственно (табл. 1). Древостой (II класса бонитетa) имеет характерный для климаксовых черневых кедровников смешанный кедрово-пихтовый состав и сложную вертикальную структуру (табл. 2).
Таблица 1.
Общая характеристика микрогруппировок растений в кедровнике осочково-крупнотравно-папоротниковом
| Микрогруппировка растений с доминированием | ОПП*, % | Высота травяного яруса, см | Число видов | Сомкнутость древесного полога | Доля площади на ПП, % |
|---|---|---|---|---|---|
| Oxalis acetosella L. под группами Pinus sibirica и Abies sibirica | 20–30 | 3–8 | 11–13 | 0.9–1.0 | 7 |
| Carex macroura Meinsh. под группами Pinus sibirica и Abies sibirica | 70–80 | 20–40 | 16–18 | 0.8–0.9 | 20 |
| Aconitum septentrionale Koelle и Carex macroura под группами Pinus sibirica со вторым ярусом Abies sibirica | 60–70 | 70–120 | 20–26 | 0.6–0.7 | 13 |
| Calamagrostis obtusata Trin. и Dryopteris expansa (C. Presl) Fraser-Jenk. & Jermy под группами и отдельно стоящими деревьями Pinus sibirica | 60–70 | 60–70 | 30–35 | 0.4–0.6 | 12 |
| Athyrium filix-femina (L.) Roth, Dryopteris expansa,Diplazium sibiricum (Turcz. ex G. Kunze) Kurata) в “окнах” и под отдельно стоящими деревьями Pinus sibirica | 90–100 | 80–120 | 41–45 | 0.1–0.4 | 47 |
Таблица 2.
Основные таксационные показатели древостоя (данные за 2017 г.)
| Ярус | Состав (возраст, лет) | Средняя высота, м | Запас древесины, м3 га–1 | Полнота | Число деревьев, шт. га–1 | |
|---|---|---|---|---|---|---|
| К | П | |||||
| I | 7К(246) 3К(330) + П(182) ед. К(422) | 31.4 | 482 | 0.75 | 128 | 8 |
| II | 9П(134) 1К(172) | 22.3 | 60 | 0.22 | 8 | 129 |
| III | 6П(74) 3П(50) 1Б(50) | 12.2 | 16 | 0.19 | – | 347 |
В 2017 г. проведено очередное обследование ПП. Выполнено общее геоботаническое описание пробной площади по ранее разработанной методике [8], а также описание каждой микрогруппировки растений с определением их площади по результатам картирования ПП в масштабе 1 : 100. Номенклатура сосудистых растений приведена в соответствии со сводкой С.К. Черепанова [9]. У каждого дерева на ПП (виргинильного, молодого, зрелого и старого генеративного состояния) измерены диаметр на высоте 1.3 м (с точностью до 2 см) и высота (с точностью до 0.5 м). Особи предъювенильного, ювенильного и имматурного состояний подсчитаны на 45 площадках каждая размером 4 м2. Учетные площадки размещены равномерно по пяти типам микрогруппировок растений, пропорционально площади последних. Для молодых особей подсчитаны встречаемость и численность на единицу площади (1 га) в пределах каждой микрогруппировки растений и всей ПП. Встречаемость рассчитана как отношение числа учетных площадок с молодыми особями к общему числу площадок, выраженное в %. Для анализа динамики численности возобновления по годам использованы собственные учеты 2007 и 2011 гг., а также данные наблюдений на ПП за период 1960–2005 гг., опубликованные ранее [10, 11 и др.].
Онтогенетическое состояние Pinus sibirica определяли по работе С.С. Николаевой с соавт. [12], а Abies sibirica – по И. Д. Махаткову [13]. Онтогенетическую структуру ценопопуляций ключевых видов оценивали по ранее разработанной методике [14, 15 и др.].
Возраст деревьев, каждое из которых пронумеровано, актуализировали по данным инструментального определения, выполненного в 1999 г. Возраст молодых особей определяли по мутовкам и модельным деревьям. Возрастную структуру древостоев оценивали по общепринятой классификации [16, 17 и др.]. При анализе возрастных рядов характеризовали форму его распределения: диапазон, коэффициенты асимметрии и эксцесса [18].
При оценке санитарного состояния особей древесных видов за основу принята шкала В.А. Алексеева [19]: 1-я категория – здоровые деревья; 2-я – ослабленные; 3-я – сильно ослабленные; 4-я – усыхающие; 5-я – свежий сухостой; 6‑я – старый сухостой.
РЕЗУЛЬТАТЫ И ИХ ОБСУЖДЕНИЕ
Сопоставление численности особей различного онтогенетического и санитарного состояния видов ценозообразователей кедровника осочково-крупнотравно-папоротникового (рис. 1) дает общее представление о структуре древесного ценоза. Она характеризуется явным численным превосходством Abies sibirica (число деревьев Pinus sibirica – 136 шт. га–1, Abies sibirica – 484 шт. га–1), однако это не уменьшает ценотической роли P. sibirica (запас древесины P. sibirica – 473 м3 га–1, A. sibirica – 84 м3 га–1), которая, отличаясь большей продолжительностью жизни, стабильно занимает господствующее положение в пологе (см. табл. 2). Механизм устойчивости такой структуры древесного ценоза раскрывается при подробном анализе строения ценопопуляций P. sibirica и A. sibirica.
Рис. 1.
Онтогенетические спектры ценопопуляций ключевых видов в климаксовом кедровнике осочково-крупнотравно-папоротниковом. Санитарное состояние особей: 1 – здоровые и ослабленные, 2 – сильно ослабленные и усыхающие, 3 – свежий и старый сухостой. Здесь и на рис. 2 онтогенетические состояния: jv – ювенильное, im – имматурное; v – виргинильное, g1 – молодое генеративное, g2 – зрелое генеративное, g3 – старое генеративное.
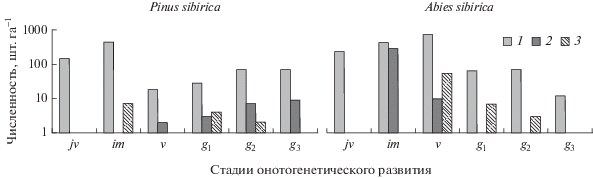
Онтогенетическая структура ценопопуляции P. sibirica (см. рис. 1) характеризуется нормальным состоянием с полночленным двухвершинным спектром. В численности преобладают имматурные (55%) и ювенильные (18%) особи, а также зрелые и старые генеративные особи (в сумме составляющие 19% численности популяции). Важно отметить, что сенильных особей P. sibirica нами не найдено, как и в работе С.А. Николаевой с соавт. [12], так как наиболее старые деревья (старше 480 лет), несмотря на имеющиеся признаки ослабления, сохраняют генеративную функцию. Наименьшей численностью отличаются особи виргинильного состояния (2%).
Двухвершинность онтогенетического спектра P. sibirica вызвана биологическими и экологическими особенностями вида. Во-первых, достаточно высокой теневыносливостью ювенильных и имматурных особей, резко снижающейся при переходе от имматурного к виргинильному состоянию [2], в связи с чем усиливается отпад. Во-вторых, накоплением зрелых и старых генеративных особей разного возраста, связанным с большой продолжительностью их жизни. Таким образом, несмотря на то, что число виргинильных растений в онтогенетическом спектре невелико, сам спектр соответствует нормальному состоянию, и популяция P. sibirica может считаться устойчивой.
Возрастная структура древостоя P. sibirica относится к циклично-разновозрастному типу. Она характеризуется полным возрастным рядом (от 1 до 540 лет), островершинным распределением с положительной асимметрией (коэффициент асимметрии – 1.4; значение эксцесса – 3.4), что свидетельствует о численном доминировании особей старших возрастов. Строение древостоя по 20-летним возрастным периодам имеет отчетливый “волновой” характер, отражающий дискретность возобновительного процесса (рис. 2). Максимумы возрастной структуры приходятся на 260, 320, 160, 400, 500 и 100 лет, наибольший всплеск численности – 260 лет.
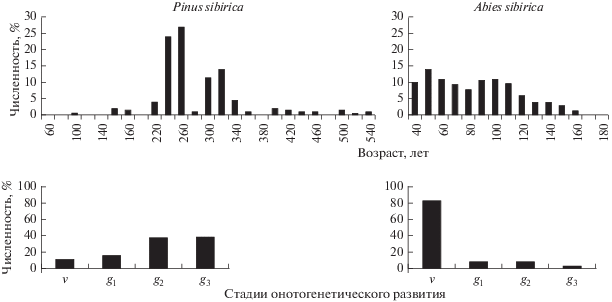
В отличие от возрастной структуры онтогенетический спектр древостоя P. sibirica выглядит более сглаженным (см. рис. 2) за счет меньшего числа градаций, несопоставимых с календарным возрастом (от нескольких десятков лет продолжительности виргинильной стадии до 150–200 лет – зрелой генеративной). В результате такой неравномерной по времени группировки распределение онтогенетической структуры приобретает отрицательную (правостороннюю) асимметрию.
При сопоставлении возрастной и онтогенетической структур ценопопуляции P. sibirica выявлено, что особи старой генеративной стадии онтогенетического развития (78 деревьев на 1 га) характеризуются наибольшим варьированием по возрасту. К ним отнесены деревья со средним возрастом от 330 ± 40 до 540 ± 40 лет в ослабленном и усыхающем состоянии и, реже, усыхающие. Они достигли максимальных для данных условий диаметра (92 ± 8 см) и высоты (33.5 ± 1.5 м). Особи зрелого генеративного состояния в здоровом, ослабленном, усыхающем и сухостойном состояниях (79 деревьев на 1 га) имеют возраст около 246 ± 40 лет. Менее многочисленные особи на молодой генеративной (35 деревьев на 1 га) и виргинильной (20 деревьев на 1 га) стадиях развития в здоровом, редко в ослабленном состоянии имеют средний возраст 132 ± 80 лет. Имматурные (8–80 лет) и ювенильные (1–10 лет) особи P. sibirica отличает хорошее санитарное состояние (в сумме на ПП составляющие 600 ± 36 шт. га–1, из них 98% в здоровом состоянии или с незначительными признаками ослабления). При этом их численность существенно варьировала по годам (в основном особей в возрасте до 5, реже 10 лет) за счет их периодического появления и гибели, что особенно характерно для участков под сомкнутыми кронами деревьев.
Наличие микрогруппировок растений, отличающихся по составу и мощности развития трав, является показателем достаточно выраженной неоднородности биотопических условий, что влияет на процесс возобновления (табл. 3 ). На площади, занятой микрогруппировками Oxalis acetosella под наиболее сомкнутыми биогруппами деревьев, возобновление полностью отсутствовало во все годы наблюдений. Наиболее успешно P. sibirica возобновляется при относительном затенении пологом древостоя (см. табл. 1): более стабильно (по годам) в микрогруппировках с доминированием Aconitum septentrionale и Carex macroura и доминированием Calamagrostis obtusata и Dryopteris expansa, менее стабильно – в микрогруппировках с доминированием Carex macroura. В окнах, где развиваются микрогруппировки с доминированием Athyrium filix-femina, Dryopteris expansa и Diplazium sibiricum, молодые особи P. sibirica размещаются в основном на микроповышениях, образованных ветровальными комплексами и валежом. Именно в “окнах”, занимающих 47% площади насаждения, подрост меньше всего испытывает конкуренцию со стороны материнского полога и, несмотря на низкую численность, непрерывно выходит в основной ярус древостоя. Об этом свидетельствует частое групповое размещение по площади всех особей P. sibirica (в виде линейных структур), связанное с приуроченностью возобновления к валежу.
Онтогенетический спектр Abies sibirica (см. рис. 1) характеризуется полночленностью и выраженной левосторонней асимметрией: численно доминируют особи ювенильного (13%), имматурного (38%) и виргинильного (39%) состояний. Суммарная численность молодых, зрелых и старых генеративных особей составляет 8%. Это свидетельствует о непрерывном возобновлении A. sibirica и постепенном изреживании, усиливающемся со временем.
Таблица 3.
Характеристика возобновления Pinus sibirica в различных микрогруппировках растений
| Микрогруппировки растений с доминированием | Ювенильные особи | Имматурные особи | ||
|---|---|---|---|---|
| число, шт. га–1 | встречаемость, % | число, шт. га–1 | встречаемость, % | |
| Oxalis acetosella | – | – | – | – |
| Carex macroura | – | – | 896 ± 687 | 25 |
| Aconitum septentrionale и Carex macroura | 178 ± 29 | 32 | 478 ± 136 | 42 |
| Calamagrostis obtusata и Dryopteris expansa | 132 ± 12 | 14 | 489 ± 13 | 14 |
| Athyrium filix-femina, Dryopteris expansa, Diplazium sibiricum | 102 ± 43 | 14 | 330 ± 205 | 43 |
Возрастная структура древостоя A. sibirica (см. рис. 2) характеризуется распределением с выраженной положительной асимметрией (коэффициент ассиметрии 0.43, значение эксцесса минус 0.6), что свидетельствует о доминировании особей младших возрастов. Онтогенетический спектр древостоя сохраняет те же закономерности.
Малочисленные особи старой генеративной стадии развития (12 деревьев на 1 га) в возрасте 182 ± 20 лет имеют удовлетворительное санитарное состояние. Более многочисленные зрелые генеративные особи (74 дерева на 1 га) в возрасте 134 ± 20 лет находятся в здоровом и, редко, в усыхающем состоянии (4% от числа деревьев данной стадии развития). Молодые генеративные деревья (76 деревьев на 1 га) имеют большее варьирование по возрасту – от 74 ± 20 до 34 ± 20 лет. Аналогичная амплитуда возраста характерна для самых многочисленных виргинильных особей (836 деревьев на 1 га) – от 74 ± 20 до 50 ± 20 лет. На этих двух стадиях развития увеличено число ослабленных, усыхающих и сухостойных деревьев (до 10% численности особей каждой стадии). Еще более плохое санитарное состояние отмечено у имматурных особей (750 деревьев на 1 га; 40% численности особей данной стадии находятся в сильно ослабленном состоянии) в возрасте от 5 до 40 лет. Напротив, ювенильные особи (250 деревьев на 1 га) в возрасте до 10 лет имеют хорошее санитарное состояние. Таким образом, численность молодых особей пихты в возрасте до 20–30 лет постоянно накапливается, а затем происходит сильный отпад, чаще всего вызванный поражением грибными болезнями, в основном Melampsorella caryophyllacearum Chroet [20].
В отличие от возобновления P. sibirica имматурные и ювенильные особи A. sibirica (в сумме на ПП составляющие 1000 ± 72 шт. га–1) довольно равномерно размещены по всей площади (встречаемость 62%). Возобновление полностью отсутствует только в условиях сильного затенения в микрогруппировках Oxalis acetosella (табл. 4). При меньшей сомкнутости древесного полога (см. табл. 1) в микрогруппировках с доминированием Carex macroura образование молодых особей улучшается. Наиболее успешно оно происходит при относительном затенении пологом древостоя в микрогруппировках растений с доминированием Aconitum septentrionale и Carex macroura, а также с доминированием Calamagrostis obtusata и Dryopteris expansa. В “окнах” в микрогруппировках с доминированием Athyrium filix-femina, Dryopteris expansa и Diplazium sibiricum возобновление A. sibirica менее успешно и так же, как у P. sibirica, приурочено к микроповышениям.
Таблица 4.
Характеристика возобновления Abies sibirica в различных микрогруппировках растений
| Микрогруппировки растений с доминированием | Ювенильные особи | Имматурные особи | ||
|---|---|---|---|---|
| число, шт. га–1 | встречаемость, % | число, шт. га–1 | встречаемость, % | |
| Oxalis acetosella | – | – | – | – |
| Carex macroura | – | – | 417 ± 208 | 25 |
| Aconitum septentrionale и Carex macroura | 132 ± 15 | 42 | 1097 ± 61 | 42 |
| Calamagrostis obtusata и Dryopteris expansa | 357 ± 18 | 86 | 2024 ± 124 | 76 |
| Athyrium filix-femina, Dryopteris expansa, Diplazium sibiricum | 351 ± 143 | 43 | 526 ± 206 | 29 |
В отличие от P. sibirica способность A. sibirica в рассматриваемых условиях давать массовые всходы и их большая теневыносливость [2] позволяют ей постоянно образовывать возобновление (более успешное под пологом крупных деревьев P. sibirica), лучше выдерживая конкуренцию как со стороны травяного яруса, так и древостоя. Отпад особей происходит непрерывно на всех стадиях развития, с некоторым его усилением на виргинильной и молодой генеративной стадиях, при внедрении особей в основной полог древостоя, как следствие высокой конкуренции со стороны господствующего полога [21, 22]. Абсолютно-разновозрастная структура древостоя A. sibirica соответствует представлениям о непрерывном самовоспроизводстве и устойчивости ценопопуляций древесных видов.
Таким образом, постоянное взаимодействие ценопопуляций Pinus sibirica и Abies sibirica в условиях высочайшей фитоценотической роли травяной растительности обеспечивает стабильность всей экосистемы. Являясь основным эдификатором сообщества, P. sibirica формирует общий характер ее мозаики [3]. Однако успешность его возобновления в условиях господства в травяном ярусе эколого-ценотических групп видов крупнотравья и крупных лесных папоротников во многом зависит от динамики ценопопуляции A. sibirica. Последняя непрерывно пополняет древостой новыми поколениями, более успешно образуя биогруппы в фитогенных полях деревьев старших поколений P. sibirica. Под их пологом снижается мощность травяного яруса, а затем постоянно формируются (и периодически гибнут) новые поколения P. sibirica. Выход этих поколений в основной ярус древостоя сопряжен с периодами частичного распада биогрупп деревьев A. sibirica (при достижении возраста 160–190 лет), в результате происходит смена возрастных поколений P. sibirica. Менее интенсивное, но более стабильное пополнение ценопопуляции P. sibirica происходит за счет формирования молодых поколений на полуразложившемся валеже в “окнах”, возникающих после гибели старовозрастных деревьев кедра.
ЗАКЛЮЧЕНИЕ
В целом оба ключевых вида климаксовых черневых осочково-крупнотравно-папоротниковых кедровников Западного Саяна (Pinus sibirica и Abies sibirica) характеризуются устойчивым состоянием ценопопуляций. Нормальное состояние полночленного двухвершинного онтогенетического спектра P. sibirica обеспечивается непрерывным образованием молодых поколений и длительным накоплением взрослых особей в пологе древостоя (на протяжении 300–400 лет). На этапе перехода из имматурного в виргинильное состояние особи P. sibirica чаще всего выживают в уже существующих “окнах”, где возобновление крайне немногочисленно (на валеже), либо этот переход приурочен к моменту распада старых биогрупп A. sibirica (в возрасте 160–190 лет), под пологом которых постоянно накапливается существенное количество имматурных особей P. sibirica (до 896 ± 687 шт. га–1). Во втором случае внедрение особей P. sibirica в полог древостоя имеет более массовый характер, но происходит мозаично с периодичностью 60–100 лет (по всей площади, занятой элементарной популяцией P. sibirica). В результате формируются выраженный “волновой” характер возрастной структуры и групповое размещение древостоя P. sibirica. Нормальное состояние полночленного онтогенетического спектра A. sibirica с выраженной левосторонней асимметрией свидетельствует о постоянной насыщенности ценопопуляции молодыми поколениями. Формирующаяся в результате абсолютно-разновозрастная структура древостоя A. sibirica характеризует устойчивость ценопопуляции.
Рассмотрение онтогенетической структуры с учетом пространственной мозаики сообщества позволило получить представление о характере взаимодействия двух основных видов-ценозообразователей климаксовых черневых кедровников. Успех совместного устойчивого произрастания этих двух видов кроется в некотором различии экологических требований и существенной разнице в продолжительности генеративного периода развития, который для P. sibirica продолжается более 400 лет, а для A. sibirica – 80–100 лет. В каждой отдельной пространственной группировке всегда происходит волновой процесс смены численного доминирования P. sibirica и A. sibirica, приводящий к их динамическому равновесию на значительной площади ненарушенных черневых лесов. Особо следует отметить, что в образовании устойчивых потоков поколений ключевых видов важное значение имеют ветровальные комплексы и крупный валеж, которые обеспечивают не такое массовое, но непрерывное пополнение древостоя молодыми поколениями на открытых участках.
Таким образом, устойчивость черневых кедровников Западного Саяна, ранее подвергавшаяся сомнению [10], подтвердил анализ онтогенетических и возрастных структур ценопопуляций Pinus sibirica и Abies sibirica с учетом горизонтальной мозаики биогеоценоза, относящегося к фоновому для данной территории типу леса.
Исследование выполнено в рамках Базового проекта № 0356-2016-0708 и при частичной финансовой поддержке РФФИ (проект № 18-05-00781 А).
Список литературы
Типы лесов гор Южной Сибири. Новосибирск: Наука, 1980. 336 с.
Поликарпов Н.П., Чебакова Н.М., Назимова Д.И. Климат и горные леса Южной Сибири. Новосибирск: Наука, 1986. 226 с.
Nazimova D.I., Danilina D.M., Stepanov N.V. Rain-Barrier Forest Ecosystems of the Sayan Mountains // Botanica Pacifica. A Journal of Plant Science and Conservation. 2014. V. 3(1). P. 39–47. https://doi.org/10.15372/ SJFS20150401
Степанов Н.В. Сосудистые растения приенисейских Саян: Дис. … докт. биол. наук. Красноярск, 2014. 791 с.
Кедровые леса Сибири / Семечкин И.В., Поликарпов Н.П., Ирошников А.И. и др. Новосибирск: Наука, 1985. 257 с.
Поликарпов Н.П. Комплексные исследования в горных лесах Западного Саяна // Вопросы лесоведения. Красноярск, 1970. Т. 1. С. 26–79.
Сукачев В.Н. Некоторые общие вопросы теоретической фитоценологии // Вопросы ботаники. М., 1954. С. 290–300.
Сукачев В.Н., Зонн С.В. Методические указания к изучению типов леса. М.: Изд-во АН СССР, 1961. 144 с.
Черепанов С.К. Сосудистые растения России и сопредельных государств (в пределах бывшего СССР). СПб., 1995. 990 с.
Поляков В.И., Семечкин И.В. Динамика и устойчивость черневых кедровников Западного Саяна // Лесоведение. 2004. № 4. С. 1–14.
Поляков В.И. Черневые кедровники Западного Саяна: контроль и прогнозирование хода роста. Новосибирск: Изд-во СО РАН, 2007. 181 с.
Николаева С.А., Велисевич С.Н., Савчук Д.А. Онтогенез кедра сибирского в условиях Кеть-Чулымского междуречья // Вестник Томского гос. ун-та. Биология. 2008. № 3(4). С. 24–34.
Махатков И.Д. Поливариантность онтогенеза пихты сибирской // Бюл. МОИП. Отд. биол. 1991. Т. 96. Вып. 4. С. 79–88.
Восточноевропейские леса: история в голоцене и современность. М.: Наука, 2004. 575 с.
Ставрова Н.И. Структура популяций древесных растений на разных стадиях восстановительных сукцессий в лесах Европейского Севера России // Актуальные проблемы геоботаники: III Всероссийская школа-конференция. Петрозаводск: Карельский научный центр РАН, 2007. С. 397–407.
Дыренков С.А. Структура и динамика таежных ельников. Л.: Наука, 1984. 174 с.
Семечкин И.В. Структура и динамика кедровников Сибири. Новосибирск: Изд-во СО РАН, 2002. 253 с.
Noss R.F. Indicators for monitoring biodiversity: a hierarchical approach // Conservation Biology. 1990. V. 4. № 4. P. 355–364.
Алексеев В.А. Диагностика жизненного состояния деревьев и древостоев // Лесоведение. 1997. № 4. С. 51–57.
Сташкевич Н.Ю., Данилина Д.М., Сенашова В.А. Оценка состояния подроста Pinus sibirica Du Tour и Abies sibirica Ledeb. в смешанных производных лесах черневого пояса Западного Саяна // Вестник КрасГАУ. 2013. № 9. С. 145–149.
Ермоленко П.М. Микроклимат хвойно-лиственных молодняков в связи с их фитоценотической структурой в черневом подпоясе Западного Саяна // Стационарные лесоводственные исследования в Сибири. Красноярскск: Изд. Ин-та леса и древ. СО РАН, 1978. С. 52–97.
Коловский Р.А. О механизме корневой конкуренции // Лесоведение. 1968. № 1. С. 37–43.
Дополнительные материалы отсутствуют.