

Экология, 2020, № 2, стр. 141-148
К экологии размножения малой бурозубки (Sorex minutus L.) на северной периферии ареала
Э. В. Ивантер *
Петрозаводский государственный университет
185910 Петрозаводск, просп. Ленина, 33, Россия
* E-mail: ivanter@petrsu.ru
Поступила в редакцию 29.05.2019
После доработки 20.06.2019
Принята к публикации 26.06.2019
Аннотация
На основе многолетних стационарных и экспедиционных исследований на таежном Северо-Западе России (1958‒2017 гг.) анализируются популяционно-экологические особенности размножения малой бурозубки, связанные с ее обитанием у северных границ распространения. Установлено, что в этих условиях для вида характерен более продолжительный, длящийся с середины апреля по конец сентября репродуктивный период, отличающийся участием в размножении в основном лишь перезимовавших зверьков. Общее число выводков за сезон – два, чему соответствуют и два четких пика расселения молодых: в конце июня‒начале июля и начале августа. Среднее число эмбрионов на самку составляет 6.0 ± 0.21 с колебаниями от 2 до 11. Величина помета меняется по месяцам, закономерно снижаясь от начала лета к осени.
Стационарные и экспедиционные исследования малой бурозубки проводились в течение 60 лет (1958‒2017 гг.) в Восточной Фенноскандии. Эта территория, помимо Карелии, охватывает ряд прилежащих регионов таежного Северо-Запада России ‒ Карельский перешеек Ленинградской области и таежные массивы Кольского полуострова. Обитая в условиях северной периферии ареала, малая бурозубка вырабатывает весьма эффективные структурно-популяционные адаптации и благодаря этому успешно преодолевает экстремальные северные условия, сохраняя достаточно высокие показатели репродукции и лидирующее положение в формирующемся населении мелких млекопитающих. Так, по уровню численности она стабильно занимает в нем твердое третье место, уступая лишь обыкновенной бурозубке и рыжей полевке. Тем не менее экология и особенно размножение малой бурозубки по-прежнему исследованы недостаточно. Настоящая публикация представляет собой попытку восполнить этот существенный пробел.
МАТЕРИАЛ И МЕТОДЫ
Материалом для работы послужили многолетние сборы зверьков, отловленных двумя основными методами: ловушко-линиями (плашки Геро) и ловчими 30-метровыми канавками. Первым было отработано 243 600 ловушко-суток и добыто 297 малых бурозубок, вторым – 5400 канавко-суток и отловлено 2096 зверьков. В итоге общее число добытых и проанализированных по состоянию репродуктивной системы землероек составило 2393 экз.
Для изучения размножения малой бурозубки всех отловленных зверьков подразделяли на две возрастные группы. В качестве основных возрастных критериев использовали форму (конфигурацию и состояние швов) черепа и степень стертости зубов, а второстепенных (вспомогательных) ‒ особенности снашивания волосяного покрова на хвосте, лапах и ушных раковинах. Кроме того, в сомнительных случаях привлекали и некоторые другие показатели, в частности размеры тимуса, который, как известно, полностью инволюирует к осени первого года жизни и у перезимовавших зверьков практически не выражен.
Особи первой возрастной группы (сеголетки) характеризуются высокой сжатой с боков черепной коробкой (череп “вздутый”) и отсутствием гребней. Швы между костями хорошо заметны. Зубы имеют острые вершины и гребни. Эмаль коронок не стерта. Половая система, как правило, недоразвита. Тимус хорошо выражен, его абсолютный вес 15‒250 мг. Хвост равномерно покрыт волосами (“пушистый”). Волосяной покров на лапках и ушных раковинах хорошо развит и почти не имеет следов снашивания. В большинстве случаев это неполовозрелые зверьки (subadultus).
Зверьки второй возрастной группы (перезимовавшие) отличаются утолщенным черепом с хорошо развитыми гребнями и слабо заметными швами, последние имеют зигзагообразный рисунок. Вершины промежуточных зубов в результате снашивания округленные, притупленные; коренные зубы уплощены, с тупыми конусами и гребнями. Пигментация на зубах почти не заметна. Волосяной покров хвоста, лап и ушных раковин сильно сношен, а местами совсем отсутствует (“лысины”). Это половозрелые зверьки (adultus).
Различаются возрастные группы и по строению половой системы (табл. 1). У неполовозрелых самцов семенники очень малы и, как правило, имеют округлую форму (диаметр около 1 мм, вес не более 5‒6 мг). Придатки семенника и семенные пузырьки недоразвиты. У половозрелых, перезимовавших бурозубок семенники значительно крупнее (вес двух семенников обычно более 150 мг), имеют продолговатую форму (длина 3‒12 мм, ширина 2‒6 мм) и хорошо заметны даже у невскрытых зверьков (в виде двух вздутий на нижней стороне тела). Придатки семенника, простатическая и куперова железы и семенные пузырьки очень хорошо выражены; последние наполнены спермой и сильно вздуты. У самцов, участвующих в размножении, кроме того, увеличены боковые кожные железы (7 × 3 мм), появление которых принимается в качестве симптома полового созревания [1, 2].
Таблица 1.
Характеристика половой системы малой бурозубки
| Показатели | Зимовавшие | Прибылые |
|---|---|---|
| Самцы | ||
| Число исследованных зверьков | 192 | 538 |
| Длина семенника, мм | 5.8 (1.0‒11.5) | 1.1 (0.8‒7.0) |
| Длина семенных пузырьков, мм | 6.1 (2.1‒16.7) | 5.0 (3.8‒6.0) |
| Вес двух семенников, мг | 124.7 (89.0‒150.0) | 5.7 (0.7‒4.0) |
| Самки | ||
| Число исследованных зверьков | 38 | 373 |
| Суммарная длина рогов матки, мм | 17.5 (6.6‒35.2) | 9.0 (3.0‒16.0) |
| Длина влагалища (тела матки), мм | 8.5 (3.3‒13.0) | 5.2 (1.5‒9.2) |
У молодых неполовозрелых самок матка полупрозрачная, тонкая и короткая; общая длина ее рогов составляет в среднем 9.52 (2.8‒22.0) мм, тела 6.6 (1.7‒22.4) мм. У перезимовавших половозрелых самок матка набухает, утолщается и становится шире, стенки ее гиперемированы, рыхлы. В длину она увеличивается почти втрое: суммарная длина ее рогов 17.5 (6.6‒35.2) мм, тела 8.5 (3.3‒13.0) мм. У взрослых зверьков тело матки составляет в среднем 35% от длины рогов, а у молодых ‒ 46%. При этом соотношение (индекс) тела и длины рогов матки у малой бурозубки несколько иное, чем у других представителей рода Sorex. Например, у обыкновенной (Sorex araneus) и средней (S. caecutiens) бурозубок эти индексы составляют соответственно 65‒75 и 40‒50%. Таким образом, у малой бурозубки рога матки относительно короче, а тело длиннее, чем у других видов.
У кормящих или ранее кормивших самок соски обсосаны, и волосяной покров вокруг них сильно вытерт (особенно если самка кормит второй выводок), но у первых млечные железы лактируют, а у вторых ‒ нет. У беременных на ранних стадиях хорошо заметны розоватые желтые тела в яичниках, позднее ‒ эмбрионы на рогах матки. Матка сильно увеличена в длину и ширину. Плацентарные пятна образуются, но очень быстро (уже на 2‒3-й день после родов) исчезают [1, 3]. Иногда удавалось определить и недавно оплодотворенных самок, признаки беременности у которых еще не были выражены. В матке у таких самок иногда заметна мутная беловатая жидкость ‒ сперма.
Сроки и интенсивность размножения обыкновенной бурозубки определяли на основании предпринятого нами специального гистологического исследования полового цикла самцов и самок [7]. Остальные исследования проводили по общепринятым методикам, неоднократно описанных в наших предыдущих публикациях [5‒7].
РЕЗУЛЬТАТЫ И ИХ ОБСУЖДЕНИЕ
Согласно полученным данным, в половом цикле самцов малой бурозубки, как и у других видов Soricidae, с момента рождения и до марта наблюдается период покоя, характеризующийся присутствием в семенных канальцах только клеток Сертоли и сперматогониев (рис. 1a). Размеры и вес семенников в этот период минимальные (0.8‒1.5 мм, вес двух семенников 0.7‒4 мг), диаметр семенных канальцев 30‒40 мкм, придатки и семенные пузырьки не выражены.
Рис. 1.
Поперечные срезы извитых канальцев семенника малой бурозубки в состоянии пассивного (а) и активного (б) сперматогенеза.
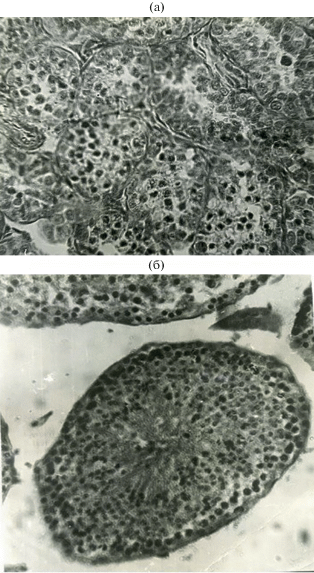
С февраля‒марта и до середины апреля для самцов малой бурозубки характерен пассивный сперматогенез, т.е. стадия подготовки к массовому продуцированию сперматозоидов. В это время наблюдаются начало митотической активности сперматогониев и усложнение зародышевого эпителия семенных канальцев до сперматид. При этом первый этап пассивного сперматогенеза приходится у землероек на февраль и сопровождается появлением отдельных сперматоцитов I порядка, но они еще не включаются в развитие эпителия и дегенерируют.
Далее следует второй этап, характеризующийся увеличением размеров гениталий (рис. 2). Длится он с конца февраля до середины апреля и характеризуется последовательным развитием зародышевого эпителия от однорядного до многорядного с клетками Сертоли, сперматогониями, сперматоцитами обоих порядков и сперматидами (см. рис. 1б). Вес семенников увеличивается к этому времени в 10‒15 раз (с 1.3‒2.6 мг в июне‒феврале до 20‒80 мг в марте), а средний диаметр семенных канальцев приближается к максимальному (50‒80 мкм). Зародышевый эпителий тоже изменяется. В феврале он двухрядный: первый ряд состоит из выстилающих базальную мембрану клеток Сертоли и сперматогониев, а второй ‒ из сперматоцитов I порядка, которые в массе появляются во всех семенных канальцах, а сами канальцы достигают 30 мкм. К середине марта зародышевый эпителий семенных канальцев преобразуется в многорядный: первый ряд содержит клетки Сертоли и сперматогонии, второй ‒ сперматоциты I порядка, третий ‒ сперматоциты II порядка и сперматиды. Диаметр канальцев увеличивается в среднем до 90 мкм.
Рис. 2.
Изменение длины (1) и веса двух семенников (2), суммарной длины рогов матки (3) и влагалища (4) на протяжении жизненного цикла малой бурозубки: А – длина семенника, мм: Б – вес двух семенников, г; В – длина влагалища, мм; Г – суммарная длина рогов матки, мм.
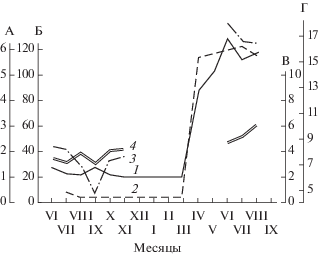
Активный сперматогенез, начинающийся в апреле, состоит из двух стадий: преобразование сперматид в зрелые сперматозоиды и интенсивное продуцирование и формирование спермиев (см. рис. 1б). Семенники достигают в это время максимальных размеров и веса, увеличиваясь в несколько раз ‒ от 20‒30 мг в марте до 100‒130 мг в апреле и июне (см. рис. 2). Весь период активного сперматогенеза продолжается у самцов малых бурозубок с середины апреля до самого конца их жизни, т.е. до начала октября, когда были отловлены последние в этот год взрослые зверьки. Причем в это время в семенных канальцах присутствуют все клеточные элементы ‒ от клеток Сертоли и сперматогониев до зрелых сперматозоидов, в массе собирающихся в семенных канальцах. При этом никаких признаков затухания сперматогенеза не наблюдается. Диаметр семенных канальцев составляет в среднем от 145 до 180 мкм, а вес двух семенников ‒ от 140 до 250 мг.
Таким образом, общая гистологическая картина сперматогенеза демонстрирует у малой бурозубки стабильную активность с конца апреля до октября без каких-либо подъемов и спадов, соответствующих периодам спаривания и паузам между ними.
Эстральный цикл самок малой бурозубки по срокам соответствует сперматогенезу самцов, но наблюдается несколько позднее. Период полового покоя длится у самок с рождения и до конца марта. Он характеризуется наличием в яичниках примордиальных, полостных и атрезирующих фолликулов разной выраженности и величины (рис. 3). С конца апреля‒первых чисел мая гистологическая картина яичников демонстрирует определенное оживление, выражающееся прежде всего в увеличении числа фолликулов на разных стадиях развития ‒ от примордиальных до граафовых пузырьков; некоторые полостные фолликулы близки к разрыву. Эпителий слизистой влагалища также характеризует состояние эструса: он 10‒15-рядный, в нижних рядах хорошо заметны ороговение клеток и их усиленная дисквамация, в это время они в массе наблюдаются в просвете влагалища.
Рис. 3.
Гистологическое строение яичников и слизистой влагалища у малой бурозубки в различных стадиях полового цикла: а – яичник молодой неполовозрелой самки в состоянии покоя; б – слизистая влагалища взрослой самки в период эструса; в – яичник взрослой самки с желтым телом и полостными фолликулами.
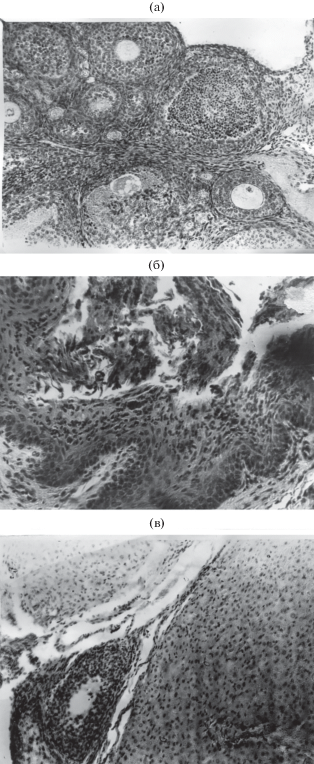
С появлением беременных самок (начало июня) гистологическая картина их органов размножения заметно меняется. В яичниках образуются желтые тела, примордиальные и полостные фолликулы на разных стадиях развития и атрезии. Матка гиперемирована и на поперечном разрезе имеет звездчатую форму с секретом маточных желез в просвете. После родов наблюдается спад фолликулярной активности, рост фолликулов замедляется и одновременно усиливается их атрезия.
Одновременно с описанными гистологическими изменениями в яичнике и матке изменяются размеры и вес этих органов. При этом эструсы способны повторяться в соответствии с числом пометов, а вторая беременность может наступить еще до окончания кормления первого выводка.
Как и у других видов землероек в изучаемом регионе, у малой бурозубки спаривание начинается в конце апреля‒начале мая, хотя самцы созревают несколько раньше. В мае величина семенников уже достигает 6 мм и более с весом 90‒110 г и более, что подтверждают и сроки отлова молодых зверьков. Судя по тому, что прибылые появляются в наших отловах лишь с третьей декады июня (22.06.1959, 26.06.1961, 21.06.1962, 28.06.1971, 22.06.1978, 24.06.1983), малая бурозубка начинает размножаться в середине мая, т.е. в более поздние сроки, чем обыкновенная и средняя. К такому же выводу на основании наблюдений в Сибири приходит и Б.С. Юдин [8]. В то же время Р. Брембелл и К. Холл [4], а также С. Боровский и А. Денель [2] считают, что у малой бурозубки размножение начинается на 15‒30 дней раньше, чем у обыкновенной. С этим согласна и А.Э. Айрапетьянц [9]. С другой стороны, Н. Михельсeн [10], исследовавшая оба вида бурозубок в Нидерландах, вообще не обнаружила у них разницы в сроках размножения.
Начальный период воспроизводства популяции малой бурозубки в Карелии, как, впрочем, и во многих других местах ареала, например в условиях Барабы [11], отличаются высокой интенсивностью. Так, за все годы исследований в первой половине лета нам ни разу не удавалось встретить взрослую самку с угнетенной или неразвитой половой системой. Об этом свидетельствует, кроме того, и хорошо проявляющийся резкий подъем подвижности малой бурозубки в июне, во многом обусловленный массовым выходом из гнезд молодых первой генерации. В результате повышения подвижности репродуцирующих особей малой бурозубки в конце июня‒начале июля заметно увеличивается в отловах и число беременных самок. Среди них основу составляют особи со следами более раннего кормления выводка или лактирующие на момент отлова.
Очередная волна расселения прибылых наблюдается в Карелии ближе к середине августа, тогда как, например, в Барабе почти на месяц раньше – во вторую половину июля, т.е. через 40‒50 дней после первой. Это соответствует приведенным нами выше для малой бурозубки срокам продолжительности беременности и кормления молодняка. В конечном счете это позволяет считать, что второй подъем половой активности взрослых самок начинается сразу за распадом выводков первой генерации.
Продолжающееся более месяца, до середины‒конца августа, медленное и постепенное нарастание численности прибылых зверьков было вызвано, скорее всего, некоторым замедлением интенсивности размножения. Например, в Барабе уже в конце первой декады августа в уловах регистрируются взрослые, закончившие размножение самки. Можно также предположить, что начавшийся в июне очередной подъем размножения малой бурозубки сильно растянут во времени. Это можно объяснить и неодновременным вступлением в размножение части молодых самок (их созревание и вступление в репродукцию в год рождения в сибирской части ареала далеко не редкость), и выселением из гнезд особей третьего помета, в результате чего волны расселения молодых второй генерации перекрываются с третьей.
Как видно из рис. 4, на котором представлена динамика отлова сеголеток по пятидневкам, в условиях таежного Северо-Запада России основная масса зимовавших малых бурозубок приносит как минимум два помета. Им соответствуют два четких пика выхода и расселения прибылых: в конце июня‒начале июля и начале августа. Исходя из 20-дневной беременности и 20‒30-дневного периода развития в гнезде [10], можно рассчитать сроки спаривания и нарисовать общую картину размножения.
Рис. 4.
Динамика отлова молодых малых бурозубок в годы массового размножения: 1 – заповедник “Кивач” (1959 г.); 2 – Ладожский стационар “Каркку” (1969 г.); по оси абсцисс – пятидневки.
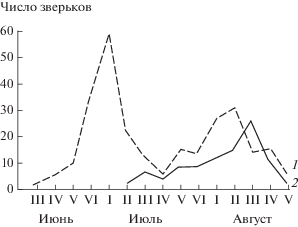
Первое спаривание у этих зверьков происходит, по-видимому, в период с 5‒10 по 15‒20 мая, т.е. приблизительно на 10‒15 дней позднее, чем у прочих видов бурозубок. В июне, как уже указывалось, большинство взрослых самок беременны, а со второй половины месяца ‒ беременны или кормящие (табл. 2). Рождение молодых первого выводка (ему соответствует первая волна их выхода и расселения в конце июня‒начале июля) наблюдается между 1 и 15 июня. Второе спаривание (судя по меньшей выраженности соответствующего ему пика расселения молодых, оно охватывает не всех взрослых бурозубок и, кроме того, значительно растянуто во времени) происходит через 30‒40 дней после первого (приблизительно 15‒30 июня). С этого времени и до конца сентября взрослые самки либо беременны, либо лактируют, либо (реже) не имеют следов второго размножения (см. табл. 2). Рождение детенышей происходит в первой половине июля, массовый выход и их расселение ‒ в первой половине августа.
Таблица 2.
Интенсивность размножения взрослых самок малой бурозубки в Карелии (по материалам отлова 1959‒2014 гг.)
| Месяц | Общее число исследованных самок | Относительное количество самок, % от общего числа | |||
|---|---|---|---|---|---|
| беременных | беременных и одновременно кормящих | кормящих | без следов размножения | ||
| Июнь | 12 | 50.0 | ‒ | 8.3 | 41.7 |
| Июль | 43 | 27.9 | ‒ | 7.0 | 65.1 |
| Август | 64 | 12.2 | 1.4 | 12.2 | 74.3 |
| Сентябрь | 4 | 50.0 | ‒ | 25.0 | 25.0 |
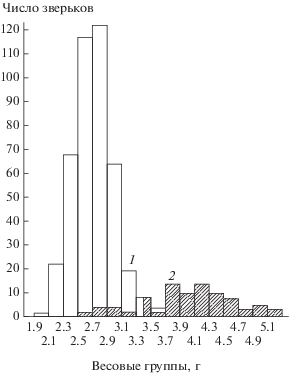
Первая беременная самка этого вида добыта 09.06.1959, последняя ‒ 11.09.1966 (заповедник “Кивач”). Следовательно, общая продолжительность периода размножения малой бурозубки составляет 125‒130 дней, т.е. короче, чем у других видов землероек. Третья генерация если и бывает, то у очень немногих самок и не каждый год. Об этом свидетельствует двухвершинный характер кривой отлова молодых и более значительная, чем у размножающейся до 3 раз в лето обыкновенной бурозубки, доля самок без следов размножения. Из более чем 1100 исследованных нами молодых зверьков лишь у трех (самки, добытые 16.08.1970, 22.07.1987 и 19.07.1993) обнаружено участие в размножении – беременность или следы кормления выводка с 6 послеродовыми пятнами в матке. Следует добавить, что у малой бурозубки, как и у обыкновенной, прибылые особи, приближаясь по длине тела и черепа к взрослым, значительно уступают им по весу (рис. 5).
Отсутствие или крайне редкие случаи (до 5%) полового созревания самок-сеголеток малой бурозубки в Англии наблюдали Р. Брембелл и К. Холл [3, 4], в Польше ‒ З. Пуцек [12], в Средней Сибири ‒ Н.Ф. Реймерс [13], на Тянь-Шане ‒ А.К. Кыдыралиев [14]. Между тем в Ленинградской и Архангельской областях, в Башкирии и на Среднем Урале значительная часть самок нормально созревает и размножается в год рождения [9, 15‒17].
По нашим данным , число эмбрионов, приходящихся на одну самку малой бурозубки, варьирует от 2 до 11 и составляет в среднем по 53 случаям 6.0 ± 0.21 (табл. 3). Наиболее часто (70%) встречаются выводки из 5 и 6 детенышей. Величина помета меняется по месяцам, закономерно снижаясь от первой половины лета к осени. В сравнении с данными из других частей ареала (табл. 4) плодовитость карельских малых бурозубок выглядит невысокой. Анализ эмбрионов отмечен только дважды: у самки, добытой 05.08.1959 в заповеднике “Кивач”, у которой резорбции подверглись, очевидно, все эмбрионы, и у самки, отловленной на стационаре Каркку 11.07.1973, ‒ у нее рассосались 3 из 5 эмбрионов.
Таблица 3.
Величина выводка малой бурозубки в Карелии
| Сезон | n | Вариация числа зверьков на одну самку | Среднее число эмбрионов | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 11 | |||
| Июнь | 15 | ‒ | 1 | 2 | 2 | 3 | 3 | 2 | 1 | ‒ | 1 | 6.4 ± 0.21 |
| Июль | 14 | 1 | 1 | ‒ | 4 | 5 | 1 | 1 | ‒ | 1 | ‒ | 5.7 ± 0.19 |
| Август | 24 | 1 | 2 | 4 | 5 | 5 | 2 | 2 | 1 | 1 | 1 | 5.8 ± 0.22 |
| Всего | 53 | 2 | 4 | 6 | 11 | 13 | 6 | 5 | 2 | 2 | 2 | 6.0 ± 0.21 |
Таблица 4.
Плодовитость малой бурозубки в разных частях ареала
| Район исследований | n | Вариации величины выводка | Среднее | Источник |
|---|---|---|---|---|
| Польша | 34 | 2‒9 | 6.0 | [2] |
| Карелия | 53 | 2‒11 | 6.0 | Наши данные |
| Бассейн Вычегды (Коми) | 20 | 4‒9 | 7.1 | [22] |
| Ленинградская обл. | ‒ | ‒ | 7.2 | [9] |
| Волжско-Камский край | 18 | 5‒12 | 8.1 | [23] |
| Томская обл. | 6 | 5‒10 | 7.7 | [11] |
| Западная Сибирь | 19 | 4‒12 | 7.5 | [8] |
| Барабинская низменность | 19 | 4‒8 | 5.8 | [11] |
| Горный и степной Алтай | 4 | ‒ | 7.0 | [24] |
| Юг Средней Сибири | 4 | 3‒9 | 7.0 | [13] |
ЗАКЛЮЧЕНИЕ
Отмеченные выше экологические особенности размножения малой бурозубки, характерные для специфических условий северной периферии ареала, ‒ лабильность и разнообразие адаптивных реакций репродуктивной сферы, их трансформация и смена во времени и пространстве и, наконец, выработка широких, проявляющихся на популяционном уровне адаптивных комплексов, отличающихся динамичностью и высокой скоростью компенсаторной перестройки, вполне укладываются в современную концепцию периферических популяций, разработанную применительно к широко распространенным политипическим видам наземных животных, к которым принадлежит и малая бурозубка.
Проведенный нами анализ закономерных географических отличий, характеризующих популяционную специфику репродукции зверьков этого вида, обитающих на северной периферии видового ареал, подтвердил выдвинутое нами ранее положение [7, 18‒21], согласно которому в экологическом центре (оптимуме) видового ареала население вида благодаря функционированию внутрипопуляционных компенсаторных механизмов отличается не только высоким уровнем численности, но и большей устойчивостью, тогда как у северных границ распространения эти стабилизирующие механизмы практически отсутствуют, и организация населения изменяется в широком диапазоне и с большей амплитудой. Соответственно более выражены и гораздо рельефнее проявляются близ северных границ ареала и специфические структурно-популяционные адаптации, направленные на максимально эффективное воспроизводство видового населения, способное обеспечить видам и их популяциям необходимое преодоление экстремальных условий экологического пессимума. Таково, например, отмеченное нами для северных зон ареала более частое созревание и участие в размножении прибылых зверьков.
Вместе с тем для периферических популяций малой бурозубки характерны и так называемые “популяционные волны”, возникающие в связи с чрезвычайно резкими и неритмичными изменениями численности [7, 21]. Эти резкие колебания численности, наряду с вызываемыми ими ужесточением отбора и специфической перестройкой пространственной, возрастной и экологической структуры, значительно активизируют микроэволюционные процессы и обеспечивают популяционные преобразования, ведущие к завоеванию видом новых территорий, смене экологической ниши, формированию новых популяций и даже видов.
Список литературы
Дунаева Т.Н. К изучению биологии размножения обыкновенной бурозубки (Sorex araneus) // Бюл. МОИП. Отд. биол. 1955. Т. 60. Вып. 6. С. 27‒43.
Borowski S., Dehnel A. Materiali do biologii Soricidae // Ann. Univ. M. Currie-Sklodowska. 1952. № 7. P. 13‒21.
Brambell R. Reproduction in the common shrew (S. araneus L.). 1. The oestrus cycle of the female // Phil. Trans. Soc. B. 1935. V. 518. № 225. P. 123‒134.
Brambell R., Hall K. Reproduction of the lesser shrew (Sorex minutus L.) // Proc. Zool. Soc. 1937. V. 49. P. 89‒103.
Ивантер Э.В., Ивантер Т.В. Обыкновенная бурозубка в заповеднике “Кивач” // Тр. гос. запов. “Кивач”. Петрозаводск, 1968. Вып. 1. С. 115‒135.
Ивантер Т.В., Ивантер Э.В., Терноушко Е.И. Биология размножения и структура популяций землероек (Soricidae) Карелии // Вопросы экологии животных. Петрозаводск, 1974. С. 95‒143.
Ивантер Э.В. Популяционная экология мелких млекопитающих таежного Северо-Запада СССР. Л.: Наука, 1975. 246 с.
Юдин Б.С. Экология бурозубок (род Sorex) Западной Сибири // Вопросы экологии, зоогеографии и систематики животных / Тр. Биол. ин-та СО АН СССР. Новосибирск, 1962. Вып. 8. С. 33‒134.
Айрапетьянц А.Э. Насекомоядные и грызуны // Звери Ленинградской области. Л.: Изд-во ЛГУ, 1970. С. 47‒165.
Michielsen N.C. Intraspecific and interspecific competition in the shrews Sorex araneus L. and S. minutus L. // Arch. Neer-land. Zool. 1966. V. 17. № 1. P. 73‒174.
Глотов И.Н., Ердаков Л.Н., Кузякин В.А. и др. Сообщества мелких млекопитающих Барабы. Новосибирск: Наука, 1978. 231 с.
Pucek Z. Seasonal and age changes in weight of internal organs of shrews // Acta theriol. 1960. V. 10. № 26. P. 34‒176.
Реймерс Н.Ф. Птицы и млекопитающие южной тайги Средней Cибири. М.; Л.: Наука, 1966. 419 с.
Кыдыралиев А.К. Землеройки (Soricidae) хребта Терскей-Алатау (Северный Тянь-Шань) // Зоол. журн. 1969. Т. 48. Вып. 8. С. 861‒868.
Снигиревская Е.М. Материалы по биологии размножения и колебания численности землероек в Башкирском заповеднике // Тр. Башкирского гос. запов. 1947. Вып. 1. С. 12‒18.
Долгов В.А., Чабовский В.И., Шилова С.А., Эфрон К.М. Некоторые вопросы экологии бурозубок и их значение в очагах клещевого энцефалита // Бюл. МОИП. Отд. биол. 1968. Т. 73. Вып. 6. С. 17‒28.
Пучковский С.В. Сравнительно-экологические исследования бурозубок Онежского полуострова: Автореф. дис. … канд. биол. наук. М., 1970. 21 с.
Ивантер Э.В. Эволюционная роль периферических популяций // Микроэволюция. М., 1985. С. 80‒84.
Ивантер Э.В. К изучению популяционной организации политипического вида // Уч. зап. ПетрГУ. 2008. № 1. С. 39‒60.
Ивантер Э.В. К разработке экологической концепции периферических популяций // Экология. 2017. № 1. С. 60‒65.
Ивантер Э.В. Очерки популяционной экологии мелких млекопитающих на северной периферии ареала. М.: Тов-во научн. изд. КМК, 2018. 770 с.
Куприянова И.Ф. Сем. Soricidae – землеройковые // Фауна Европейского Северо-Востока России. Млекопитающие. СПб.: Наука, 1994. Т. 2. Ч. 1. С. 11–67.
Попов В.А. Млекопитающие Волжско-Камского края. Казань, 1960. 466 с.
Юдин Б.С., Барсова Л.И. Землеройки кедровых лесов Прителецкого очага клещевого энцефалита // Природа очагов клещевого энцефалита на Алтае. Новосибирск: Наука, 1967. С. 51–59.
Дополнительные материалы отсутствуют.