

Экология, 2020, № 2, стр. 149-155
Многолетняя динамика населения рыжей полевки в Ильменском заповеднике
Н. В. Киселева *
Ильменский государственный заповедник, Южно-Уральский научный центр минералогии и геоэкологии УрО РАН
456317 Челябинская область, Миасс, Россия
* E-mail: natakis17@gmail.com
Поступила в редакцию 05.06.2019
После доработки 04.07.2019
Принята к публикации 22.07.2019
Аннотация
Приведены данные учетов численности рыжей полевки в Ильменском заповеднике с 1982 г. по 2018 г. Проанализированы изменения климатических показателей с 1928 г. по 2017 г. Существенных различий в сумме осадков за декабрь–март, апрель–май и величине средних многолетних температур за май и апрель не выявлено. Не обнаружено значимой зависимости динамики численности рыжей полевки от анализируемых климатических параметров. Изменения амплитуды и частоты волн численности рыжей полевки, вероятно, могут быть связаны с увеличением продолжительности безморозного периода, наблюдаемого на территории заповедника в последние десятилетия.
Рыжая полевка (Clethrionomys glareolus Schreber, 1780) – широко распространенный вид мелких грызунов, населяющий различные лесные ландшафты, для которого характерны циклические колебания численности. Динамику численности рыжей полевки (РП) и причины, ее определяющие, рассматривали многие исследователи для разных регионов как в оптимуме ареала [1, 2], так и на периферии [3–5], а также в различных экстремальных условиях [6]. Колебания численности РП некоторые исследователи связывают с урожаем кормов, особенно семян древесных пород – ели, сосны, пихты, дуба, липы и др. [7, 8]. Другие авторы считают, что динамика численности и структура популяций РП в первую очередь являются следствием влияния погодно-климатических и внутрипопуляционных факторов [2, 9, 10]. Существует и обобщенный взгляд на причины, вызывающие колебания численности рыжей полевки, заключающийся в том, что ни один отдельно взятый фактор не может служить единственной причиной динамики не только на протяжении всего ареала вида, но и для отдельных регионов, хотя в некоторых конкретных случаях может приобретать главенствующее значение [11].
В последние десятилетия происходят существенные климатические сдвиги, выражающиеся в изменении количества и режима осадков, отклонениях температуры и других параметров, на которые реагируют как отдельные виды, так и экологические сообщества в целом. Регулярные циклы размножения грызунов изменяются во многих районах одновременно [12–14]. Влияние отдельных погодных факторов, например снежного покрова, на численность грызунов рассматривалось подробно [15]. Однако о взаимосвязи численности животных и климата до сих пор мало что известно, в первую очередь из-за небольшого количества долгосрочных временны́х рядов данных, без которых невозможно подойти к пониманию этой связи. Кроме того, больше наблюдений выполнено на примере полевок рода Microtus, для которых типичны многолетние популяционные циклы [16].
В настоящей работе представлены результаты многолетнего мониторинга за численностью рыжей полевки в Ильменском заповеднике. Цель исследования – анализ особенностей многолетней динамики численности рыжей полевки и изучение потенциального влияния климатических изменений на ее численность. Предполагается, что долгосрочные исследования могут лучше выявить тренды в колебаниях численности населения рыжей полевки из-за климатических изменений.
МАТЕРИАЛ И МЕТОДИКА
Исследования проводили на территории Ильменского заповедника, расположенного в предгорьях восточного склона Южного Урала в подзоне предлесостепных сосновых и сосново-березовых лесов [17]. Для территории заповедника характерен сложный рельеф с разнообразным сочетанием невысоких хребтов и скалистых гряд. Климат умеренно континентальный, среднегодовая температура около +2°C, годовая сумма осадков в среднем 444 мм.
Начиная с 1982 г. учеты грызунов проводились одним и тем же исследователем по единой методике. Численность грызунов определяли с помощью учетов ловушко-линиями. Каждая линия состояла из 50 ловушек Геро, расставленных на расстоянии 10–12 м. Длительность экспозиции каждой линии – 4 сут. В качестве показателя численности использовали количество особей на 100 ловушко-суток. Показатели относительной численности рассчитывали по количеству животных, отловленных за 4 сут. Всего было отловлено 1860 рыжих полевок. Учеты проводили дважды в год – весной (май) и осенью (сентябрь) в биотопах двух типов: “влажных” и “сухих”. “Влажные” биотопы – берега озер, рек и ручьев, а также понижения рельефа, “сухие” – березово-сосновые леса горных склонов. В период с 1982 г. по 1994 г. учеты проводили в однотипных биотопах, но на разных участках, с 1995 г. по 2018 г. – на одних и тех же участках. “Влажный” биотоп – берег р. Большая Черемшанка (среднее течение), “сухой” биотоп – горный склон восточной экспозиции, покрытый березово-сосновым лесом со слабо развитым подлеском, невысоким травянистым покровом. Разница в высоте над уровнем моря между этими двумя участками составляет около 20 м. В отдельные годы дополнительно проводили отловы на разных участках в августе и октябре.
Кроме учетов линиями, в период с 1982 г. по 1987 г. численность оценивали с помощью ежемесячных (с мая по сентябрь включительно) отловов живоловушками и мечения грызунов на участке в 3.5 га, на котором отработано 20 000 лов.-сут, отловлены и помечены 732 рыжие полевки. В связи с очень низкой (близкой к нулю) численностью грызунов в период с 1986 г. по 1994 г. (8 лет) весенние учеты линиями не проводили. Всего с 1982 г. по 2018 г. отработано линиями 31 000 лов.-сут.
Гипотезу о влиянии климатических параметров на численность РП проверяли с помощью метода линейной регрессии. В качестве предикторов были использованы среднемесячные данные по сумме осадков за декабрь–март, апрель–май и среднемесячные температуры апреля и мая. Выбор этих показателей обусловлен мнением многих исследователей о зависимости уровня численности РП от состояния снежного покрова, погодных условий в период размножения, и прежде всего в мае [7, 8, 18].
Метеоданные приводятся на основании сведений, полученных на метеостанции г. Миасс, расположенной в нескольких километрах от территории заповедника. Для приблизительной оценки возможного влияния урожайности семян на численность РП были использованы данные В.Д. Захарова [19] по динамике урожайности сосны (Pinus silvestris) за период с 1986 г. по 2010 г.
РЕЗУЛЬТАТЫ
Средняя величина относительной весенней численности рыжей полевки (РП) во “влажных” биотопах за период 1982–1985 гг. (в сезоны, когда проводились учеты) и за период 1995–1999 гг. составила 0.4 ± 0.13 ос. на 100 лов.-cут, за период с 2000 г. по 2018 г. (17 лет, за исключением 2004 г.) – 2.8 ± 0.1 на 100 лов.-cут, т.е. величина весенней численности РП возросла в 7.0 раза по сравнению с периодом 1982–1999 гг. (p < 0.001). Увеличение средней величины весенней численности РП во “влажных” биотопах произошло особенно существенно с 2001 г. по 2008 г. – в этот период она составила 6.2 ± 0.3 ос. на 100 лов.-cут, за период 2009–2018 гг. снова снизилась, составив 1.8 ± ± 0.12 ос. на 100 лов.-cут (p < 0.001).
Средняя величина осенней численности РП во “влажных” биотопах за период 1982–1999 гг. составила 4.8 ± 0.5 ос. на 100 лов.-cут, с 2000 г. по 2018 г. – 18.6 ± 0.7, т.е. средняя многолетняя величина численности рыжей полевки во “влажных” биотопах заповедника возросла в 3.8 раза (p < 0.001).
В “сухих” биотопах весенняя численность рыжей полевки часто имела нулевые или близкие к нулю значения: за период с 1982 г. по 1999 г. (за те сезоны, когда проводились учеты) она составила 0.3 ± 0.09, за период с 2000 г. по 2018 г. – 4.3 ± 0.98 ос. на 100 лов.-cут (p < 0.05). Показатель весенней численности в “сухих” биотопах превышал значения данного показателя во “влажных” местообитаниях лишь в год пика (2008 г.) и дважды в годы среднего уровня численности населения (2014 и 2017 гг.). Осенняя численность, как правило, не превышала численность рыжих полевок во “влажном” биотопе, и лишь в отдельные годы достигала таких же значений, как во “влажных” биотопах (рис. 1).
Рис. 1.
Многолетняя динамика относительной осенней численности рыжей полевки в “сухих” (1) и “влажных” (2) биотопах Ильменского заповедника.
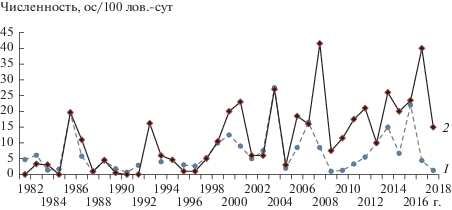
С 1982 г. по 1997 г. зарегистрированы два подъема численности рыжей полевки – в 1986 и 1993 гг. (см. рис. 1). Интервал между подъемами составил 6 лет. По данным ежемесячных учетов на участке мечения, численность рыжей полевки в 1986 г. достигла пикового значения в конце июля, и размножение полевок прекратилось. Данные по количеству отловленных полевок в конце июля приведены в табл. 1.
Таблица 1.
Численность зимовавших и сеголеток рыжей полевки, отловленных на участке мечения за июль, ос. на 1 га
| Год | 1982 | 1983 | 1984 | 1985 | 1986 | 1987 |
|---|---|---|---|---|---|---|
| Зимовавшие | 2.6 | 8.6 | 2.6 | 6.0 | 10.6 | 10.3 |
| Сеголетки | 12.3 | 31.7 | 9.7 | 16.9 | 65.1 | 32.9 |
Начиная с 2000 г. и в последующие 18 лет частота волн численности изменилась. Амплитуда подъемов возросла в 1.4–2.1 раза (рис. 2). С 2006 г. повышенная осенняя численность во “влажных” биотопах сохранялась в течение трех сезонов, затем последовал спад. Период с 2014 г. по 2017 г. характеризовался ежегодным повышением численности. За восемнадцать лет (2000–2018 гг.) относительная осенняя численность дважды (2008 и 2017 гг.) достигала пиковых значений, при которых размножение полевок прекращалось уже в июле или начале августа (см. рис. 2).
Рис. 2.
Многолетняя динамика относительной весенней (1) и осенней (2) численности рыжей полевки во “влажных” биотопах Ильменского заповедника.
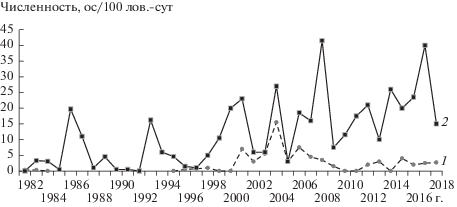
Размножение рыжей полевки в Ильменском заповеднике, как правило, начинается в апреле, первая генерация встречается в отловах в первой декаде июня. Самая ранняя дата (02.06) отлова рыжих полевок первой генерации зарегистрирована в 2008 г. Подснежное размножение нами не было зафиксировано. Заканчивается размножение обычно в конце августа–начале сентября. В 1984 г. беременных самок отлавливали в начале сентября, а самая поздняя дата отлова беременной самки, у которой обнаружено 6 эмбрионов, – 14.09.2015 г.
Между весенней численностью РП в “сухих” и “влажных” биотопах значимая корреляция обнаружена в период с 1995 г. по 2018 г. (r = 0.477 ± 0.2; p ≤ 0.05); в период с 1982 г. по 2018 г. (в сезоны, когда проводили учеты) – между весенней и осенней численностью во “влажном” биотопе (r = = 0.41 ± 0.17; p < 0.05), а также между осенней численностью в “сухих” и “влажных” биотопах (r = = 0.546 ± 0.15; p < 0.01). В связи с тем, что в общем между “сухими” и “влажными” биотопами существует корреляция численности, в регрессионном анализе рассматривали только влажные биотопы. При этом не выявлено значимых зависимостей динамики численности рыжей полевки от выбранных климатических показателей (табл. 2).
Таблица 2.
Результаты регрессионного анализа зависимости весенней и осенней численности рыжей полевки от суммарного количества осадков за декабрь–март, апрель–май и среднемесячных температур за апрель–май во “влажных” биотопах Ильменского заповедника
| Месяц | N, лет | Среднее ± SE | Min | Max | Показатели линейной регрессии | |||
|---|---|---|---|---|---|---|---|---|
| наклон | R2 | r ± SE | p* | |||||
| Весенняя численность/осадки | ||||||||
| Декабрь–март | 27 | 707 ± 6.04 | 16.2 | 157.0 | 0.014 | 0.016 | 0.125 ± 0 .198 | 0.542 |
| Осенняя численность/осадки | ||||||||
| Апрель–май | 35 | 73.47 ± 5.63 | 2.9 | 151.2 | 0.035 | 0.011 | 0.103 ± 0.173 | 0.598 |
| Осенняя численность/температура | ||||||||
| Апрель | 35 | 4.6 ± 0.49 | 1.0 | 10.7 | 1.2 | 0.096 | 0.31 ± 0.166 | 1.87 |
| Май | 35 | 11.64 ± 0.33 | 7.9 | 15.1 | 1.0 | 0.031 | 0.177 ± 0.141 | 1.03 |
Сравнение климатических параметров территории Ильменского заповедника за 89 лет не показало значительных различий в сумме осадков и величине средних многолетних температур за май и апрель (табл. 3). Однако средняя величина длительности безморозного периода в 2000–2017 гг. увеличилась на 86 дней по сравнению с 1928–1955 гг.
Таблица 3.
Многолетние климатические показатели на территории Ильменского заповедника
| Годы | Средняя сумма осадков за декабрь‒ март, мм | Среднее многолетнее кол-во осадков, мм |
Средняя многолетняя температура, °С |
Длительность безморозного периода (от 0 до 0°С), дни |
|||
|---|---|---|---|---|---|---|---|
| апрель | май | апрель | май | min–max | $\overline М $ | ||
| 1928– 1955* | 70.7 | 21.5 | 47.7 | 3.5 | 10.6 | 90–140 | 119 |
| 1983 –1999 | 57.7 | 26.6 | 42.5 | 4.1 | 10.9 | 121–226 | 190 |
| 2000 – 2017 | 73.5 | 30.1 | 47.4 | 5.1 | 12.3 | 142–237 | 205 |
* Метеоданные приводятся по С.С. Жарикову [24].
ОБСУЖДЕНИЕ
Уральский регион представляет собой восточную часть ареала рыжей полевки. Для гор Южного Урала в целом характерен мозаичный тип распределения грызунов [20]. Размещение РП по территории заповедника также очень неравномерное и обусловлено особенностями макро- и микрорельефа, зависит от состава и сомкнутости древостоя, развития подлеска и травянистой растительности, степени захламленности леса. Наиболее благоприятны для рыжей полевки берега лесных рек и ручьев, заросшие кустарником и высокотравьем, низины с обильным подлеском и валежником [21]. Суммарная площадь подобных территорий невелика – намного меньше общей площади лесов заповедника. К таким территориям относятся “влажные” биотопы, в т.ч. участок мечения рыжих полевок и долина р. Большая Черемшанка. Менее благоприятны для грызунов преобладающие по площади сосняки различных типов, так как в них слабо развит подлесок, сомкнутость крон невысокая, мало валежника и укрытий. В наших исследованиях – это “сухие” биотопы.
Весной во время снеготаяния и обильных осадков или во время сильного половодья на пониженных и прибрежных участках может происходить затопление нор полевок, из-за чего во “влажных” биотопах численность снижается, несмотря на успешную зимовку. Так, в апреле 2000 г. были обильные осадки в виде дождя – за один день (24 апреля) выпало 50% месячной нормы, и во “влажном” биотопе весной не было отловлено ни одной рыжей полевки, а в “сухом” биотопе было отловлено минимальное количество зверьков – 0.5 ос. на 100 лов.-сут. В марте 2005 г. многократно происходили дневные оттепели, после которых температура опускалась до –23°С, на снегу образовывалась корка, кроме того, обильные осадки 1.5-месячной нормы вызвали массовую гибель грызунов.
В 2010 г. исходная весенняя численность была очень низкой – в обоих типах биотопов весной не удалось отловить грызунов. Для зимы в этот год было характерно повышенное суммарное количество осадков – почти в 2 раза выше нормы и очень низкие температуры (–34…–39°С) в течение трех зимних месяцев. Глубина промерзания почвы зимой достигала 109 см, что на 3–5 см больше нормы [22]. На горных склонах, как правило, высота снежного покрова намного меньше, чем на равнинных частях и в низинах. Сильное промерзание почвы также ведет к гибели грызунов в норах.
Таким образом, анализ динамики численности грызунов с использованием среднемесячных данных погоды может быть малоинформативным, так как 1–2 дня или несколько дней с критическими погодными условиями могут сильно изменить уровень численности грызунов. Влияние погодных условий зимне-весеннего периода детально описано разными исследователями [4, 15]. Мы лишь уточнили особенности этого влияния для территории Ильменского заповедника.
Считается, что одной из причин увеличения численности грызунов, в том числе и рыжей полевки, наряду с благоприятными метеоусловиями в зоне южной европейской тайги служит хороший урожай семян древесных пород [8, 18]. В Ильменском заповеднике отсутствуют ель и пихта, липа не плодоносит под пологом леса, поэтому можно предположить, что подъемам численности грызунов благоприятствует урожайность сосны. Урожайность сосны была наибольшей в 1986, 1989, 1991, 1994, 2009 гг. [19]. Подъемы численности рыжей полевки только дважды – в 1986 и 1989 гг. – совпадают с повышенной урожайностью сосны, т.е. ее урожайность не является ведущим фактором в подъемах численности полевки в Ильменском заповеднике. Сохранению населения грызунов способствуют ландшафтная гетерогенность территории заповедника и различающиеся условия обитания во “влажных” и “сухих” биотопах. При отсутствии неблагоприятных погодных условий (длительные аномальные температуры, многократно превышающие многолетнюю норму, интенсивные осадки и др.) грызуны выживают в обоих типах биотопов.
По данным исследований в Северной Европе, в последние десятилетия на амплитуду циклов численности грызунов наиболее сильно влияет осенне-зимняя температура. Амплитуда имеет тенденцию быть высокой, если осенью и зимой очень холодно или когда сезоны являются исключительно мягкими [23]. Мы проанализировали температурные показатели за ноябрь–декабрь, январь–февраль в годы, когда наблюдались подъемы и пики численности РП: 1986, 1993, 2001, 2004, 2008, 2012, 2014 и 2017. Среднесуточные температуры за рассмотренные месяцы существенно не отличались от многолетних средних, за исключением ноября 2013 г., который был самым теплым за всю историю инструментальных наблюдений в Челябинской области (табл. 4).
Таблица 4.
Температура осенне-зимних месяцев в Ильменском заповеднике в годы пиков численности рыжей полевки
| Годы | Ноябрь | Декабрь | Январь | Февраль |
|---|---|---|---|---|
| 1985 | ‒7.8/‒7.5 | ‒11.4/‒13.2 | ‒/‒ | ‒/‒ |
| 1986 | ‒/‒ | ‒/‒ | ‒13.3/‒16.1 | ‒15.1/‒15.0 |
| 1992 | ‒4.6/‒7.3 | ‒/‒ | ‒/‒ | ‒/‒ |
| 1993 | ‒/‒ | ‒/‒ | ‒9.7/‒15.8 | ‒13.0/‒14.6 |
| 2000 | ‒6.5/–5.8 | ‒11.1/‒13.2 | –/– | –/– |
| 2001 | –/– | –/– | ‒10.9 /‒14.2 | ‒14.3/‒12.9 |
| 2003 | –6.0/–6.2 | –6.1/–13.2 | –/– | –/– |
| 2004 | –/– | –/– | –11.3/–14.8 | –10.5/–12.7 |
| 2007 | ‒6.5/‒6.2 | ‒12.1/‒12.4 | –/– | –/– |
| 2008 | –/– | –/– | – 5.5/–14.8 | –10.3/–12.7 |
| 2011* | Нет данных | |||
| 2012 | –/– | –/– | –13.8/–14.8 | –15.0/–12.7 |
| 2013 | 2.2/‒5.0 | –7.4/–12.4 | –/– | –/– |
| 2014 | –/– | –/– | –12.7/–14.8 | –13.4/–12.7 |
| 2016 | –8.1/‒5.0 | –14.8/–12.3 | –/– | –/– |
| 2017 | –/– | –/– | –10.4 /–14.8 | –12.5 /–12.7 |
Таким образом, в условиях Ильменского заповедника осенне-зимние температуры не влияют на амплитуду циклов численности рыжей полевки. Не обнаружено достоверной связи многолетней динамики численности рыжей полевки и суммы осадков за декабрь–март, апрель–май, температурой за май и апрель. Однако если значимые взаимосвязи не обнаружены, это не означает, что данные факторы биологически не важны. Возможно, эти связи маскируются другими процессами [24].
Анализ климатических показателей за 89 лет не выявил их значительных изменений, за исключением длительности безморозного периода, который увеличился в среднем на 86 дней [25] (см. табл. 3). Исходя из этого, можно было бы предположить, что размножение РП начинается раньше весной и заканчивается в более поздние сроки осенью. Однако нами не выявлены смещения в сроках размножения рыжей полевки. В условиях Удмуртии фазе “пик” в большинстве случаев предшествует подснежное размножение [2]. В условиях Ильменского заповедника подснежное размножение полевок нами не зарегистрировано.
Как уже отмечалось, анализ колебаний численности РП проводился многими исследователями в разные сезоны года в зависимости от погодных условий и урожайности основных кормов, но не рассматривалось возможное влияние увеличения длительности безморозного периода. По данным Челябинского центра по гидрометеорологии и мониторингу окружающей среды (филиал ФГБУ “Уральское УГМС”), потепление климата действительно происходит: менее чем за 100 лет температура повысилась примерно на 1.8°С. При этом скорость увеличения среднегодовых многолетних температур резко возросла после 70-х годов ХХ в. – в среднем в 2 раза. Зима стала более теплой и затяжной за счет ноября [22].
Прибылые зверьки гибнут в основном осенью, высокая смертность полевок в переходные (ранневесенний и осенний) периоды связана с сезонной перестройкой физиологии (прежде всего терморегуляции) [18]. Уменьшение длительности низкотемпературного периода снижает энергетические затраты зверьков [26, 27], при этом увеличивается возможность поиска пищевых ресурсов, улучшается их качество, за счет чего поддерживается более высокая активность животных, повышается их выживаемость, изменяется структура популяции [28, 29]. Очевидно, все это и вызывает наблюдаемые в Ильменском заповеднике изменения в амплитуде и частоте волн численности РП в последние десятилетия.
Согласно историческим сведениям, на территории Ильменского заповедника рыжая полевка в первой половине ХХ в. значительно уступала по численности другим видам грызунов, составляя в уловах от 6.0 до 34.7% [29, 30]. В настоящее время она доминирует в лесных стациях лишь в отдельные годы, уступая по численности малой лесной мыши [32, 33]. Беспрецедентное нарастание численности (до 80% в уловах) рыжей полевки за период с 1989 г. по 2013 г., зарегистрировано в предгорной тайге Печоро-Илычского заповедника. Основной причиной выхода рыжей полевки на доминирующие позиции авторы [34] считают трансформацию местообитаний – ветровальные процессы. В Ильменском заповеднике эти процессы незначительны и не играют какой-нибудь существенной роли в размещении рыжей полевки. Увеличение численности рыжей полевки в таких разных регионах, как темнохвойная тайга Северного Урала и предлесостепные сосново-березовые леса Южного Урала, свидетельствует, на наш взгляд, о сходных процессах, благоприятствующих ее расселению и увеличению численности. Мы предполагаем, что такими процессами являются глобальные климатические изменения, и в частности увеличение длительности безморозного периода.
ЗАКЛЮЧЕНИЕ
Более 80 лет в Ильменском заповеднике ведутся наблюдения за численностью и видовым составом грызунов. В начале ХХ в. рыжая полевка значительно уступала в численности другим видам. В настоящее время в лесных биотопах она доминирует, составляя в уловах до 98%. Сохранению населения рыжей полевки в сезоны с критическими погодными условиями способствуют ландшафтная гетерогенность территории заповедника и наличие разнотипных местообитаний. При анализе многолетних колебаний численности грызунов из-за кратковременных, но значительных погодных отклонений среднемесячные метеорологические показатели оказываются малоинформативными.
Начиная с 2000 г., амплитуда и частота волн численности рыжей полевки изменились, амплитуда подъемов значительно возросла. Подснежное размножение и смещение сроков размножения не зарегистрированы. Из множества климатических показателей наиболее существенно изменилась длительность безморозного периода – на территории заповедника она увеличилась на 86 дней по сравнению с началом ХХ в., составив в среднем 205 дней. Мы предполагаем, что уменьшение длительности низкотемпературного периода снижает энергетические затраты зверьков, улучшает качество их питания, повышается их выживаемость, изменяется структура населения, за счет чего происходит изменение амплитуды и частоты волн численности.
Список литературы
Буяльска Г., Лукьянов О.А., Мешковска Д. Детерминаты локального пространственного распределения численности островной популяции численности рыжей полевки // Экология. 1995. № 1. С. 35–45.
Жигальский О.А. Динамика численности и структура населения рыжей полевки (Myodes (Clethrionomys glareolus)) при зимнем и весеннем начале размножения // Зоол. журн. 2012. Т. 91. № 5. С. 619–628.
Кошкина Т.В. Мышевидные грызуны Кольского полуострова и динамика их численности // Труды Кандалакшского заповедника. 1958. Вып. 1. С. 161–191.
Ивантер Э.В., Жигальский О.А. Опыт популяционного анализа механизмов динамики численности рыжей полевки на северном пределе ареала // Зоол. журн. 2000. Т. 79. № 8. С. 976–989.
Москвитина Н.С., Кравченко Л.Б., Сучкова Н.Г. Динамика численности европейской рыжей полевки (Clethrionomys glareolus Schreber) на восточной периферии ареала // Сиб. экол. журн. 2000. № 3. С. 373–382.
Оленев Г.В., Григоркина Е.Б. Эволюционно-экологический анализ стратегий адаптации популяций грызунов в экстремальных условиях // Экология. 2016. № 5. С. 375–381. [Olenev G.V., Grigorkina E.V. Evolutionary ecological analysis of adaptation strategies of rodent populations under extreme conditions // Rus. J. of Ecology. 2016. V. 47. № 5. P. 486–492.] https://doi.org/10.1134/S106741361605009X
Попов В.А. Млекопитающие Волжско-Камского края. Насекомоядные, рукокрылые, грызуны. Казань: Изд-во Казан. фил. АН СССР, 1960. 468 с.
Башенина Н.В. Пути адаптаций мышевидных грызунов. М.: Наука, 1977. 355 с.
Турьева В.В. Мышевидные грызуны лесной части Коми АССР // Труды Коми филиала АН СССР. 1953. Вып. 1. С. 1–36.
Krebs C.J. Population Fluctuations in Rodents. Chicago: The University of Chicago Press, 2013. 306 p.
Европейская рыжая полевка. М.: Наука, 1981. С. 245–267.
Hörnfeldt B. Long-term decline in numbers of cyclic voles in boreal Sweden: analysis and presentation of hypotheses // Oikos. 2004. V. 107. P. 376–392.
Ims R.A., Henden J.-A., Killengreen S. Collapsing population cycles // Trends Ecol. Evol. 2008. V. 23. P. 79–86.
Kausrud K.L., Mysterud A., Steen H. et al. Linking climate change to lemming cycles // Nature. 2008. V. 456. P. 93–97.
Формозов А.Н. Снежный покров в жизни млекопитающих и птиц. М.: Изд-во МГУ, 1990. 286 с.
Elton C.S. Periodic fluctuations in the numbers of animals: their causes and effects // J. Exp. Biol. 1924. № 2. P.119–163.
Дорогостайская Е.В. Конспект флоры цветковых растений Ильменского заповедника // Флора и лесная растительность Ильменского государственного заповедника им. В. И. Ленина / Тр. Ильменск. гос. заповед. им. В.И. Ленина. Свердловск: УФАН СССР, 1961. Вып. 8. С. 9–50.
Ивантер Э.В. Популяционная экология мелких млекопитающих таежного Северо-Запада СССР. Л.: Наука, 1975. 246 с.
Захаров В.Д. Многолетняя динамика зимнего населения лесных птиц Ильменского заповедника // Экология. 2018. № 4. С. 299–305 [Zakharov V.D. Long-term dynamics of winter communities of forest birds in the Ilmen Nature Reserve // Rus. J. of Ecology. 2018. V. 49. № 4. P. 332–337.]https://doi.org/10.1134/S10
Большаков В.Н., Балахонов В.С., Бененсон И.Е. и др. Мелкие млекопитающие Уральских гор (экология млекопитающих Урала). Свердловск: УНЦ АН СССР, 1986. 100 с.
Киселева Н.В. Фауна мышевидных грызунов Ильменского заповедника // Растительный и животный мир Ильменского заповедника. Свердловск: УрО АН СССР, 1989. С. 3–18.
http://www.chelpogoda.ru.
Korpela K., Delgado M., Henttonen H. et al. Nonlinear effects of climate on boreal rodent dynamics: mild winters do not negate high-amplitude cycles // Global Change Biology. 2013. V. 19. P. 697–710. https://doi.org/10.1111/gcb.12099
McMillan B.R., Kaufman G.A., Kaufman D.W. Factors influencing persistence of white-footed mice (Peromyscus leucopus) // Prairie Naturalist. 2005. № 37. P. 29–40.
Жариков С.С. Климат района Ильменского заповедника и сопредельных пространств Южного Урала // Труды Ильменского государственного заповедника им. В.И. Ленина. Свердловск: УФАН СССР, 1959. Вып. VII. С. 3–32.
Coulson T., Milner-Gulland E.J., Clutton-Brock T. The relative roles of density and climatic variation on population dynamics and fecundity rates in three contrasting ungulate species // Proc. R. Soc. B-Biol. Sci. 2000. V. 267. P. 1771–1779.
Nieminen P., Hohtola E., Mustonen A.-M. Body temperature rhythms in Microtus voles during feeding, food deprivation, and winter acclimatization // J. Mammal. 2013. V. 94. P. 591–600.
Langvatn R., Albon S.D., Burkey T., Clutton-Brock T.H. Climate, plant phenology and variation in age of first reproduction in a temperate herbivore // J. Anim. Ecol. 1996. V. 65. P. 653–670.
Forbes K.M., Stuart P., Mappes T. et al. Diet quality limits summer growth of field vole populations // PLoS One. 2014. 9(3):e 91113. https://doi.org/10.1371/journal.pone.0091113
Дукельская Н.М. Опыт обзора млекопитающих Государственного Ильменского заповедника // Труды по изучению заповедников. М., 1928. С. 53–54.
Данини Е.С., Володина З.С., Котлечков В.Г. Материалы по экологии и биологии Micromammalia Ильменского заповедника и его окрестностей // Уч. зап. Молотовского ун-та. 1940. Т. 4. Вып. 1. С. 32–58.
Киселева Н.В. Об изменении фауны мышевидных грызунов Ильменского заповедника // Экология. 1990. № 5. С. 86–89.
Колчева Н.Е., Оленев Г.В. Сопряженность популяционных изменений у лесной мыши и рыжей полевки в лесных биогеоценозах Южного Урала // Экология. 1991. № 1. С. 43–52 [Kolcheva N.E., Olenev G.V. Interlinking of population changes in wood mouse and bank voles in forest biogeocoenoses of the Southern Urals // Sov. J. of Ecology. 1991. V. 22. № 5. P. 36–44.]
Бобрецов А.В., Петров А.Н., Лукьянова Л.Е., Быховец Н.М. Структурные перестройки в населении лесных полевок (Clethrionomys, Rodentia) предгорий Северного Урала // Зоол. журн. 2015. Т. 94. № 6. С. 731–738. https://doi.org/10.7868/S0044513415060070
Дополнительные материалы отсутствуют.